
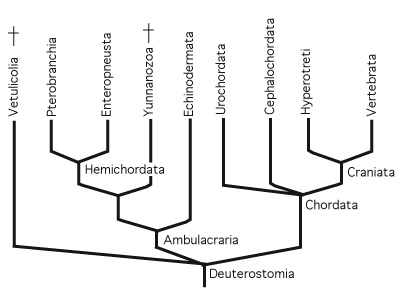
 Deuterostomia: (Cambrian - Rec.) Encompasses the major traditional phyla Chordata and Echinodermata plus several interesting minor groups.
Deuterostomia: (Cambrian - Rec.) Encompasses the major traditional phyla Chordata and Echinodermata plus several interesting minor groups.
Synapomorphies:
- Deuterostomous development
- Molecular characters
- A three-part coelom
- A pharynx (a chamber between the mouth and esophagus for the filtration of food particles from water.)
Note: Literature older than ten years has been rendered obsolete by molecular analyses that dispelled once and for all the notions the other creatures with three-part coeloms, including brachiopods, phoronids, and chaetognaths (arrow worms) might also be deuterostomes.
Deuterostome phylogeny: characterized by two major groups:
- Ambulacraria: Echinodermata and Hemichordata
- Chordata: Vertebrates and their relatives.
- Echinodermata: After mollusks and arthropods, the most commonly fossilized bilaterian taxon. Indeed, disarticulated echinoderm plates are major constituents of Phanerozoic carbonates, especially from the Early Carboniferous. Moreover, echinoderms are appealingly bizarre.
- Chordata: Contains the creatures that dominate animal biomass, occupy top of terrestrial and marine food chains, and serve as keystone species, ecologically.
{kind=link}
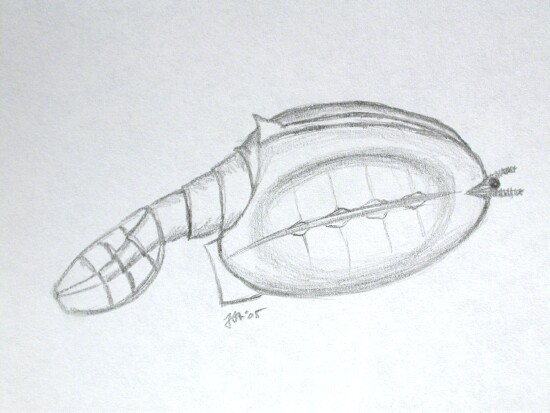
 First, a recently discovered fossil group:
Vetulicolia :
First, a recently discovered fossil group:
Vetulicolia :
- From Early Cambrian Chengjiang Fm. Lagerstätte in China
- Looks like what we might expect of something close to the ancestry of Deuterostomia.
- Front end looks like a pharynx (filter feeding organ) with five pharygeal slits
- Authors interpret ventral structure in pharynx to be endostyle (gland rich groove of basal chordates; homolog of thryoid gland of vertebrates)
- Lacks any kind of internal skeleton.
- The anterior segment is armored by plates of cuticle, giving it an arthropod-like aspect.
- The rear is a laterally compressed segmented tail. (See restoration fo Xidazoon stephanus (right) or Vetulicolia cuneata.)
{kind=link}
 Ambulacraria: Recent molecular phylogenies indicate sister taxon relationship between Echinodermata and Hemichordata. Most members are suspension or deposit feeders, although some obtain food in more interesting ways.
Ambulacraria: Recent molecular phylogenies indicate sister taxon relationship between Echinodermata and Hemichordata. Most members are suspension or deposit feeders, although some obtain food in more interesting ways.
Potential synapomorphies of hemichordates and echinoderms:
- Molecular characters
- Appendages (proboscis and feeding appendages of hemicordates, arms of echinoderms) containing coelomic extensions
- Auricularia larvae (B and C right) of holothurians resemble torneria larvae (D and F, right) of enteropneusts.
{kind=link}
{kind=link}

- Exclusively suspension and deposit feeders, however food capture is by means of ciliary action.
- Bodies divided into proboscis, collar, and trunk, each of which is invested with its own part of the three-part coelom:
- Cilia on the proboscis move food particles to mouth.
- Collar encircles mouth and proboscis.
- Trunk contains a large pharynx with pharyngeal openings (homologous to gill slits)
- Open circulatory system present, with blood propelled by contractions of primary blood vessels. No hemoglobin.
- Cilia on the proboscis move food particles to mouth.
- Dorsal nerve cord, hollow in some parts of collar.
- Bodies divided into proboscis, collar, and trunk, each of which is invested with its own part of the three-part coelom:
{kind=link}
{kind=link}
 Diversity:
Diversity:
Enteropneusta: "Acorn worms": (No record)
- Largish (5 cm - >2m) worm-like burrowing suspension or deposit feeders.
- Pharynxes large and perforated by many pairs of gill openings.
- Acorn-shaped proboscis gives common name
- Invariably solitary.
- Infaunal, inhabiting U-shaped burrows. Probably responsible for many U-shaped domichnia.
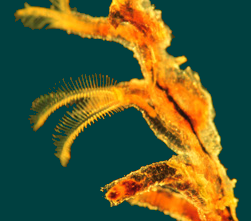 Pterobranchia: (Spotty record, Cambrian - Rec, excluding graptolites)
Pterobranchia: (Spotty record, Cambrian - Rec, excluding graptolites)
Characteristics:
- Tiny (<2 mm.) colonial critters that secrete branching colony consisting of proteinaceous tubes.
- The proboscis is developed into a cephalic shield, used to secrete the proteinaceous material of the colony structure.
- The collar is developed into one to five pairs of ciliated lophophore-like arms, each housing an extension of the coelom. Arms sport cilia-covered tentacles that transport food particles to mouth.
- Only one pair of pharyngeal openings.
- The gut is U-shaped.
- Each individual zooid lives in a cylindrical zooidal tube.
- Unlike bryozoans, zooids are able to move around on the outside of the colony, attached by a strand of contractile tissue, the peduncle to a common tissue thread, the stolon. When the colony is alarmed, individuals are quickly "reeled in." Cute.
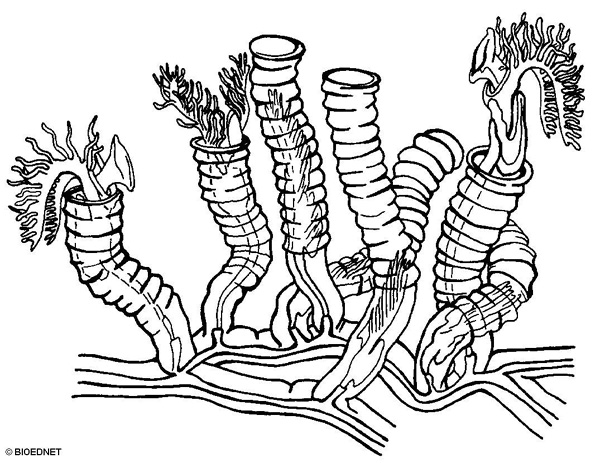
- Pterobranch growth. The stolon grows from the peduncle of the ancestral zooid. New zooids bud off from the stolon. As the stolon grows, it is encased in a creeping tube secreted by a specialized zooid, the terminal bud. Other, regular zooids bud from the stolon and secrete their own zooidal tubes.
{kind=link}
{kind=link}
 Graptolites (Cambrian - Carb.)
Graptolites (Cambrian - Carb.)
- Enigmatic fossils of the Early Paleozoic. Typically compressed into two dimensions.
- Very common and diverse in Ord - Sil with pelagic global distribution. The premier index fossils of these periods.
- Discovery of three-dimensionally preserved specimens, and subsequent study of pterobranchs led to realization that graptolite rhabdosomes were colonies of zooids that secreted a proteinaceous hard structure very similar to that of pterobranchs. Implies very close relationship.
- BUT, because they had long been studied and used as index fossile prior to being biologically understood, they have their own set of terminology which has been covered in lab. Geologists note: You may actually have to use these some day.
Graptolite diversity: There are two major groups. (We spare you the minor groups.)
- Dendroidea: (Cam. - Penn)
- Stipes branch many times in sequence, yielding a dendritic rhabdosome.
- The nema attaches the rhabdosome to the substrate.
- Thecae are dimorphic, with adjacent pairs of large autothecae and small bithecae (probably female and male zooids respectively.)
- Graptoloidea: (Latest Cam. - Dev.)
- Stipes branch at most once. See lab for rhabdosome morphology terminology based on stipe angle to nema. BUT NOTE, in some, the stipes are scandent, - i.e. they grow up the sides of the nema.
- The nema attaches the rhabdosome to a floating object. (Some genera seem to have secreted siphonophore-style floats from which numerous rhabdosomes hung.)
- Thecae are uniform.
- Monograptid morphology an unbranched stipe growing up one side of the nema. Such rhabdosomes often assume spiral shapes. Perhaps they did not attach and depended on a slow sinking rate to remain in the photic zone.
- Biostratigrapher's delight: Peak abundance and diversity in Ord. - Sil. Key index fossils for this interval.
- Dendroidea: (Cam. - Penn)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 Yunnanozoans: A final fossil group from Chengjiang (Early Cambrian), these creatures (including Yunnanozoon (right) and Haikouella) were described in the 1990s and optimistically regarded as stem chordates or even craniates (fish). Recent reinterpretation by Shu et al. suggests something more interesting - that they were stem hemichordates (basal to the enteropneust/pterobranch split) with distinct collars, pharynges, and proboscis, capable os some swimming. Opinion remains divided, with others, including Mallatt and Chen maintaining that these are stem chordates.
Yunnanozoans: A final fossil group from Chengjiang (Early Cambrian), these creatures (including Yunnanozoon (right) and Haikouella) were described in the 1990s and optimistically regarded as stem chordates or even craniates (fish). Recent reinterpretation by Shu et al. suggests something more interesting - that they were stem hemichordates (basal to the enteropneust/pterobranch split) with distinct collars, pharynges, and proboscis, capable os some swimming. Opinion remains divided, with others, including Mallatt and Chen maintaining that these are stem chordates.
Echinodermata: Including starfish, brittle-stars, sea urchins, sea cucumbers, crinoids, and a host of fossil forms.
 Echinoderm characteristics:
Echinoderm characteristics:
- Exclusively marine: Echinoderms lack osmoregulatory mechanisms that might allow them to live in brackish or fresh water.
- Skeleton is internal test comprised of individuals plates of porous high-Mg calcite. In life, the pores are occupied by a protein matrix and dermal cells. Such skeletal tissue is known as stereom. Carbonate petrologists typically call the pores "meat holes." individual birefringent elements
- One portion of the coelom develops into water vascular system. (Dissection)
- This system is passively involved in gas exchange, maintainance of posture, and locomotion.
- The latter is effected by outpouchings of the WVS that penetrate the body wall to form podia or tube feet that can be employed in suspension feeding or in locomotion, depending on the critter.
- Tube feet are arranged into five double-rows termed ambulacra. Typically, these converge on the mouth and/or anus.
- Muscles are used to pump water around the WVS, and each tube foot is equipped with small longitudinal muscles to help aim it, yet the the hydrolic force of the WVS is what primarily effects movement in most echinoderms.
- The WVS obtains water from the outside. It is connnected by a calcite-reinforced stone canal that opens to the exterior in the hydropore. The hydropore is covered by a seive-like plate, the madreporite, that strains incoming water.
- The lining of the WVS is ciliated, allowing circulation of its fluid. thus, tube-feet function as gas exchange organs.
- The coelom (including the WVS) contains coelomocytes which attack foreign material and, in some cases, carry oxygen and CO2.
- Despite this weirdness, they are proper bilateralians with a mouth, flow-through gut, and anus.
- NOTE: In addition to the WVS, echinoderms also retain a normal enterocoelic coelom.
- All but most primitive have strong pentameral symmetry.
- Mutable collagen: A form of collagen that can be partially emulsified by the application of nervous action potentials is present in crinoids.
- The dominant echinoderms of the Paleozoic were suspension-feeders. Living crinoids still suspension feed, however Eleutherozoans have other strategies. The feeding strategy of the first echinoderms is less clear, but recent discoveries suggest that they were deposit feeders.s
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
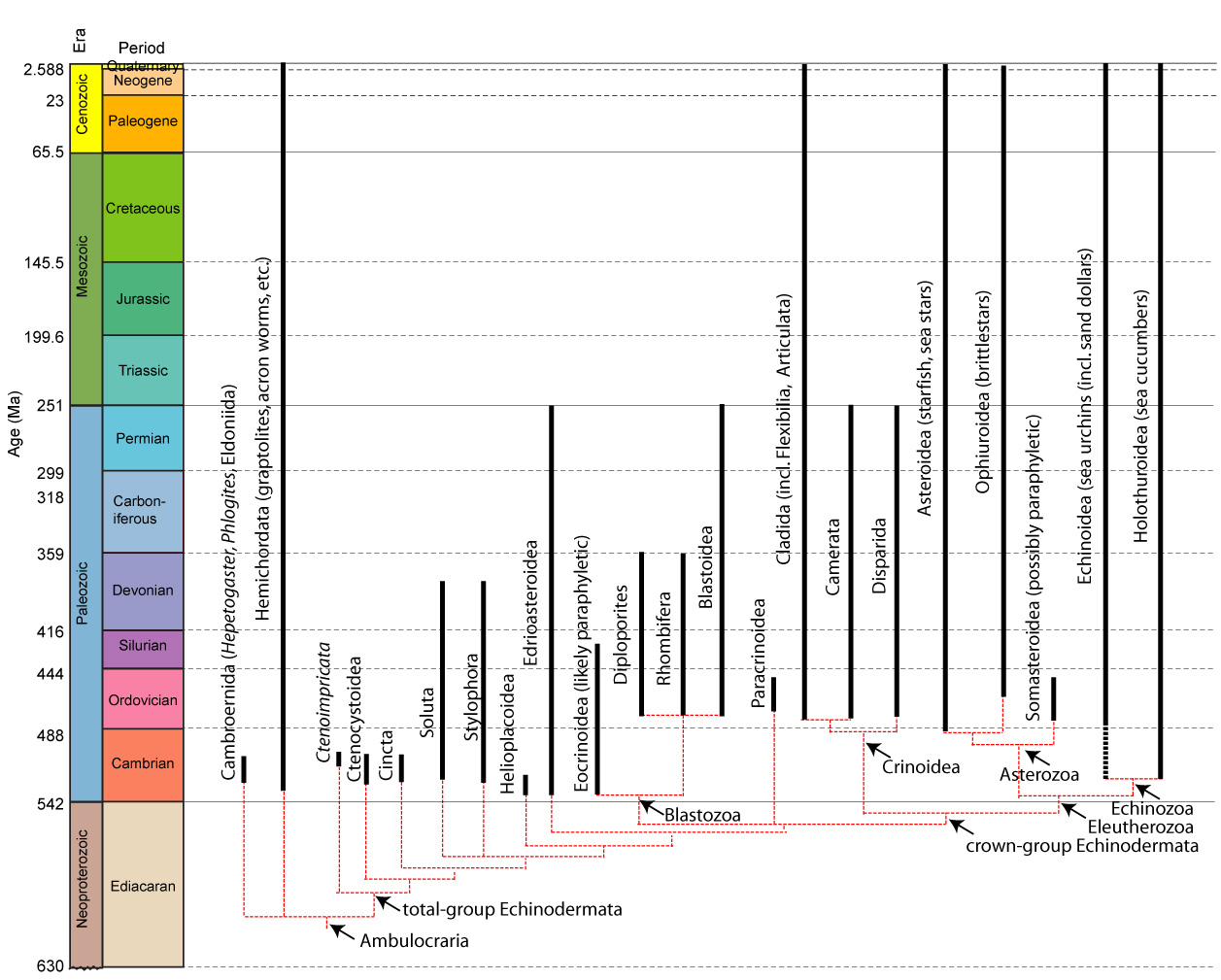 Echinoderm Systematics
Echinoderm Systematics
The cladogram at right is provided for your reference. To vertebrate paleontologists, echinoderm systematics are an issue in as much as they inform us about basal conditions for Deuterostomia and help us polarize characters at the base of Chordata. Unfortunately this has not been easy, as the earliest and most basal echinoderms are elusive and difficult to interpret. As a result, they have, at times, played a larger role in vertebrate systematics than, perhaps they should. To understand, we must examine the most basal and least understood stem-echinoderms.
 "Homolazoa": (Cam. Ord.)
"Homolazoa": (Cam. Ord.)
Echino-uglies. A problematic group of Early Paleozoic echinoderms. True apples of discord. Mono-, poly-, or paraphyletic, depending on whom you ask.
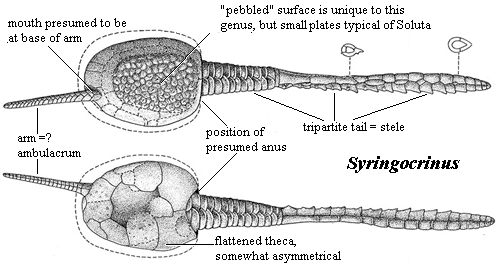 What we DO know:
What we DO know:
- Tend to have flattened thecae.
- Some with appendages including:
- aulacophore - a feeding or locomotion appendage. (See "arm" in illustration at right.)
- Stele - yet another feeding or locomotion appendage. (right)
- Some have rough bilateral symmetry, others have no obvious plane or axis of symmetry. NONE are pentamerally symmetrical.
{kind=link}
"Includes":
 Stylophora (Cambrian - Devonian). With:
Stylophora (Cambrian - Devonian). With:
- flattened theca
- elongate three-part stele ("tail")
- large openings to the left of the stele and at the opposite side from it.
- Numerous small pores on dorsal side.
- Cornuta
- Mitrata
 Soluta (Cambrian - Devonian). Similar to Stylophora but with:
Soluta (Cambrian - Devonian). Similar to Stylophora but with:
- an smaller appendage, the aulacophore opposite the stele.
- elongate stele.
- large opening near the stele.
 Cincta (Middle Cambrian). Again, similar but with:
Cincta (Middle Cambrian). Again, similar but with:
- Bilaterally symmetrical ambulacra and mouth on dorsal surface opposite the stele.
- Stele is short and stiff.
- Some indications that the stele may have ended in a holdfast.
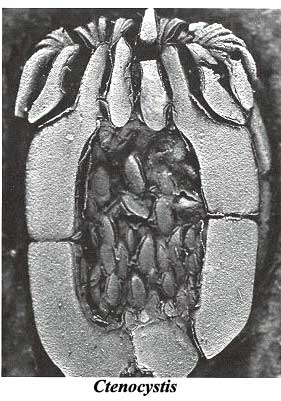 Ctenocystoidea (Cambrian - Ordovician). Strange. No stele or aulacophore. Only an approximation of bilateral symmetry, BUT...
Ctenocystoidea (Cambrian - Ordovician). Strange. No stele or aulacophore. Only an approximation of bilateral symmetry, BUT...
- Anterior and (more or less) posterior openings - mouth and anus.
- Two anterior bilaterally symmetrical ambulacra.
- Although the overall profile is more or less bilaterally symmetrical, the skeletal elements aren't.
This was the state of the art when this lecture was taught in 2010. Any attempt to speculate on ambulacrarian origins got bogged down in the Ediacaran Rohrschach-test, often with bad results. Since then, some interesting fossils have come to light, but first an excursion into paleontological pareidolia.
I have only described "homalozoan" features and named a few. No homologies with other organisms have been proposed. This is where the trouble starts. Consider the solute stele. It could be:
- an ambulacrum bearing feeding structure like the arm of a crinoid.
- a tail, used for propulsion, homologous to the chordate tail.
Similar things could be said about any of the openings of the theca, which could be mouths, anuses, pharyngeal slits, hypropores, etc. Into this chasm of ignorance steps the human imagination.
During the 1980s, Richard Jeffries of the British Museum interpreted the various homalozoans as ancestral to the vertebrates (making vertebrates a polyphyletic group within Echinodermata). This hypothesis (sort of) rests on his convictions about the homologies of the structures. Consider the stylophoran stele.
 Jeffries emphatically views it as a chordate-like tail, with room inside for a notochord and myotomes. He also claims to see pharyngeal slits in the openings of the theca. In some ways, these appendages function differently. For instance, according to Jeffries, the "tail" is used to pull the theca along over the substrate.
Jeffries emphatically views it as a chordate-like tail, with room inside for a notochord and myotomes. He also claims to see pharyngeal slits in the openings of the theca. In some ways, these appendages function differently. For instance, according to Jeffries, the "tail" is used to pull the theca along over the substrate.
His conclusion: Chordates are derived from these primitive echinoderms. Indeed, in his scheme, specific homalozoans gave rise to specific chordate groups. To emphasize the propinquity of the relationship, he coined the term Calcichordate. This hypothesis of "calcichordate" phylogeny was developed in the early days of cladistics, and Jeffries does not seem to have used a parsimony analysis.
Objections to his scheme include:
- Morphological interpretation seems far-fetched.
- The transition from them to chordates involves:
- Reorganization of body for 180 deg. change of direction of movement.
- Switch from calcium carbonate to calcium phosphate skeleton.
- Even if we accept his morphology at face value, his preferred phylogeny of deuterostomes is far from the most parsimonious tree.
- Jeffries, himself, seems to approach the issue with the style of a true-believer.
 When you add "calcichordates" to the mix, the basic pattern of deuterostome phylogeny seems completely up for grabs.
When you add "calcichordates" to the mix, the basic pattern of deuterostome phylogeny seems completely up for grabs.
As of 2011, echinoderm systematists agreed that Jeffries' phylogeny is wrong. But were all over the map otherwise. Some maintained that his assessment of homology may, in part, be right. recent analysis of the stylophoran appendage suggests that it is, indeed, a locomotor appendage. Others, such as David et al., asserted that the stylophoran appendage is an ambulacrum on a crinoid-like arm, pure and simple, and that Ctenocystoidea have blastozoan-like brachioles. To them, Homalozoa is polyphyletic and its members belong to better known groups.
But that is so 2009. In 2010 and 2012, two new fossil animals cast significant illumination on the problem:
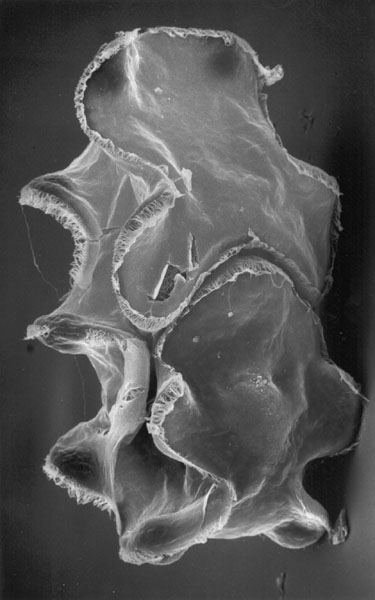
 Caron et al.. 2010: Described Herpetogaster collinsi (Cambrian, right) from the Burgess Shale, a macroscopic (1 - 2 cm total length) soft bodied creature that resembled a solitary pterobranch with:
Caron et al.. 2010: Described Herpetogaster collinsi (Cambrian, right) from the Burgess Shale, a macroscopic (1 - 2 cm total length) soft bodied creature that resembled a solitary pterobranch with:
- Two lophophore-like feeding tentacles.
- A segmented U-shaped body.
- A short stalk for attachment (temporary?) to the substrate.

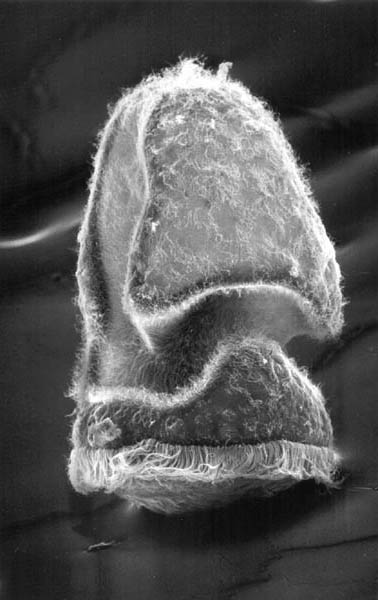
- Ctenoimbricata is bilaterally symmetrical.
- Its mouth is definitely in front and its anus is definitely in the rear.
- an echinoderm-style calcite internal skeleton with stereom and
- bilateral symmetry.
{kind=link}
Now, finally, a coherent picture of early echinoderm evolution is emerging. From a vetulicolian-like "swimming pharynx" ancestor, one can picture the evolution of both mobile yunnanozoan-like and attached Herpetogaster-like hemichordate suspension-feeders and of benthic deposit-feeding creatures like Ctenoimbricata with an internal skeleton. The subsequent history of echinoderms would then involve the achievement of alternate feeding modes (suspension, grazing, predation) and methods of locomotion.
{kind=link}
But note: The position of Stylophora is still up for grabs.
We turn to crown-group Echinodermata in the next lecture.
References:
- Clausen, S. & A. B. Smith. 2005. Palaeoanatomy and biological affinities of a Cambrian deuterostome (Stylophora). Nature 438, 351-354.
- David, B. 2000, B. Lefebvre, R. Mooi, and R. Parsley. Are homalozoans echinoderms? An answer from the extraxial-axial theory. Paleobiology 26, 4, 529-555.
- Mallatt J, J. Y. Chen. 2003. Fossil sister group of craniates: predicted and found. Journal of Morphology Oct;258(1):1-31.
- Ruta, M. 1999. Brief review of the stylophoran debate. Evolution and Development Oct;1(2):73-135.
- Shu, D. G. S. Conway Morris, J. Han, L. Chen, X.-L. Zhang, Z.-F. Zhang, H.-Q. Liu1, Y. Li & J.-N. Liu. 2001. Primitive deuterostomes from the Chengjiang Lagerstaette (Lower Cambrian, China). Nature 414, 419-424.
- Shu, D. G., X. Zhang & L. Chen. 1996. Reinterpretation of Yunnanozoon as the earliest known hemichordate. Nature 380, 428-43.