Accidental Land Animals
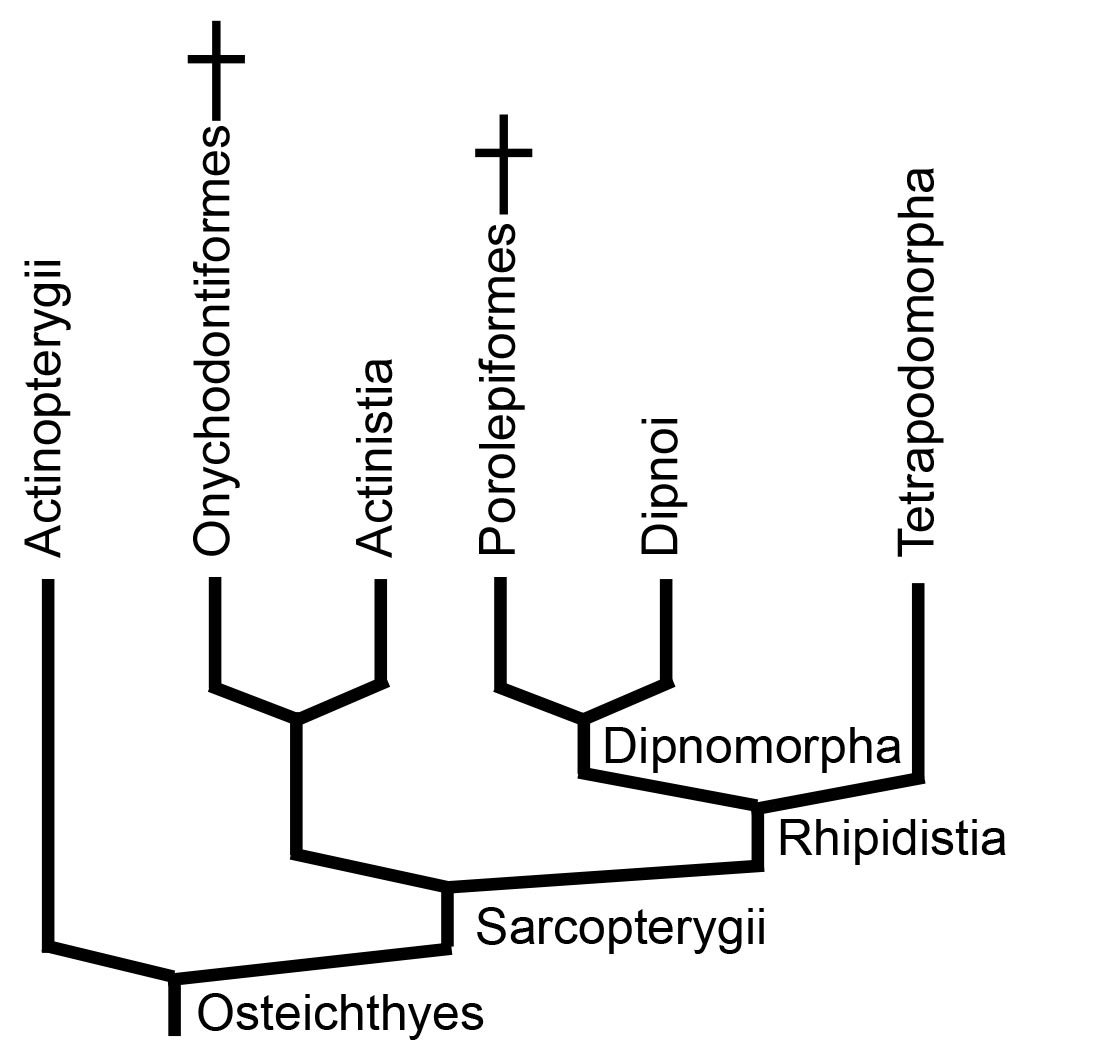
And Now For Something Completely Different: Sarcopterygii
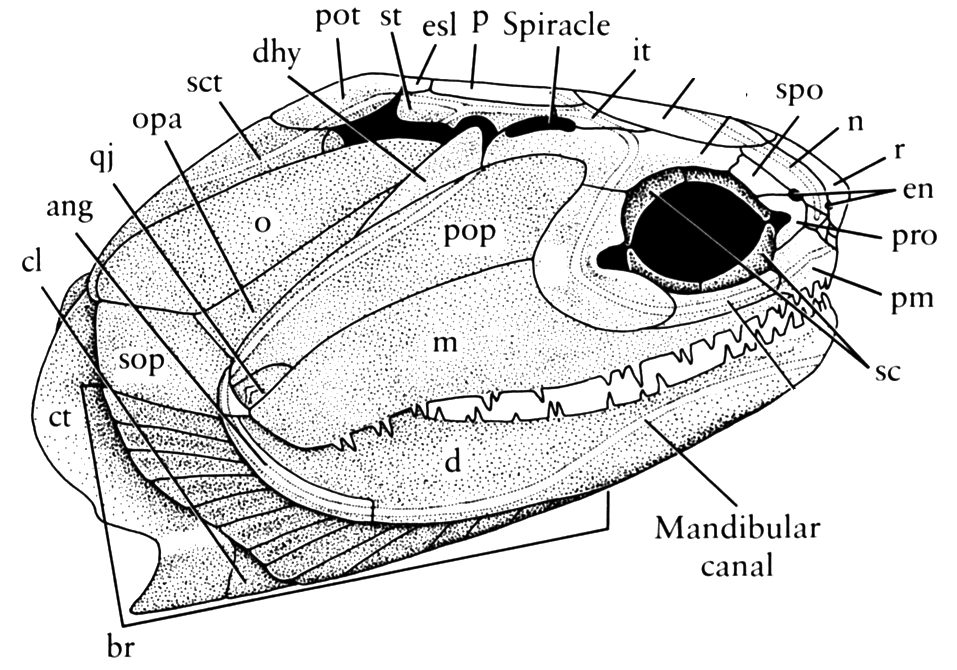
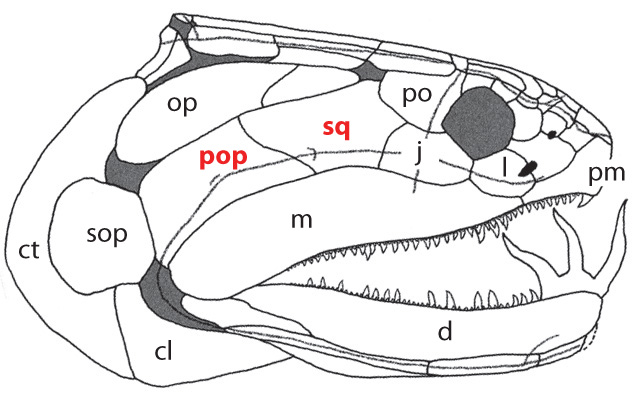
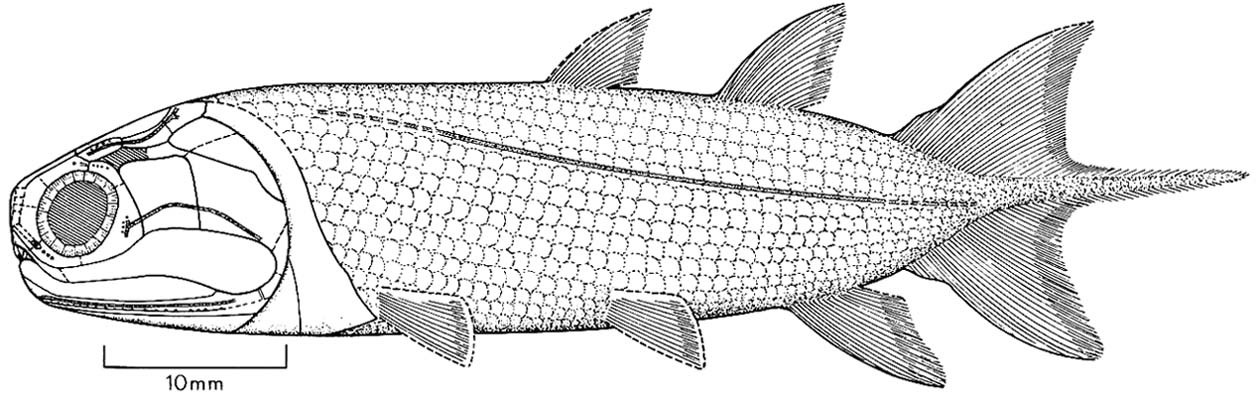
Onychodus - basal sarcopterygian (left) and Cheirolepis canadensis basal actinopterygian (right).
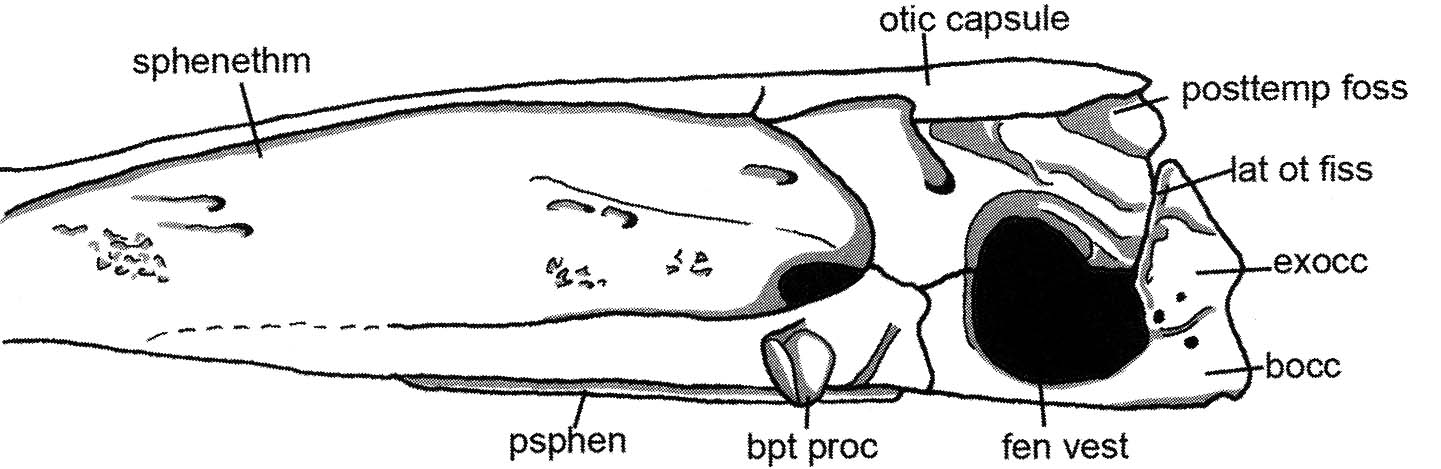
- The neurocranium ossifies as two units completely separated by the ventral cranial fissure. Their contact is a mobile joint.
- Parietal and postparietal bones in dermal skull roof meet in a dermal intracranial joint, the external manifestation of the endocranial hinge. Note: In basal sarcopterygian, the parietal roofs the sphenoid region and the postparietals, the otico-occipital region.
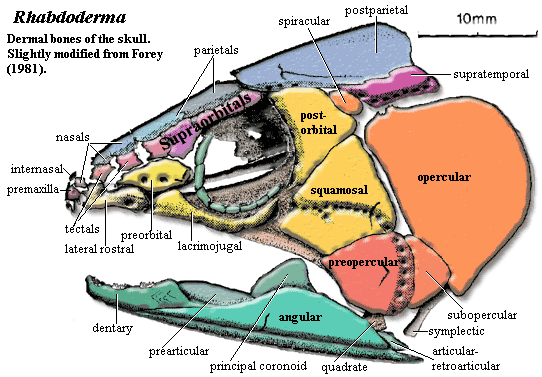
- The presence of distinct squamosal (sq) and preopercular (pop) bones in the cheek (see Onychodus - right)

Pectoral fin of rhizodontid Sauripteris - The limbs articulate with the girdles by a single element, the humerus (as in Sauripteris) or femur.
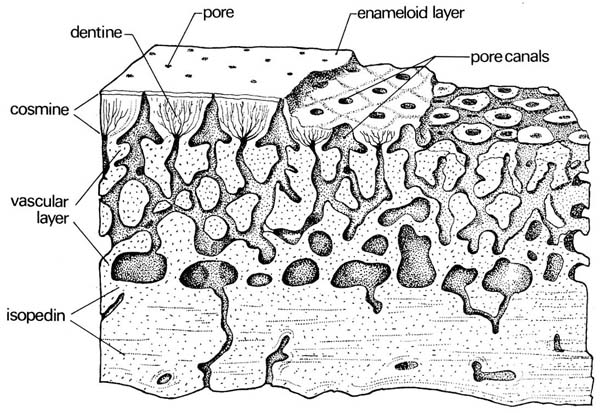
- Cosmoid scales completely lacking ganoine.
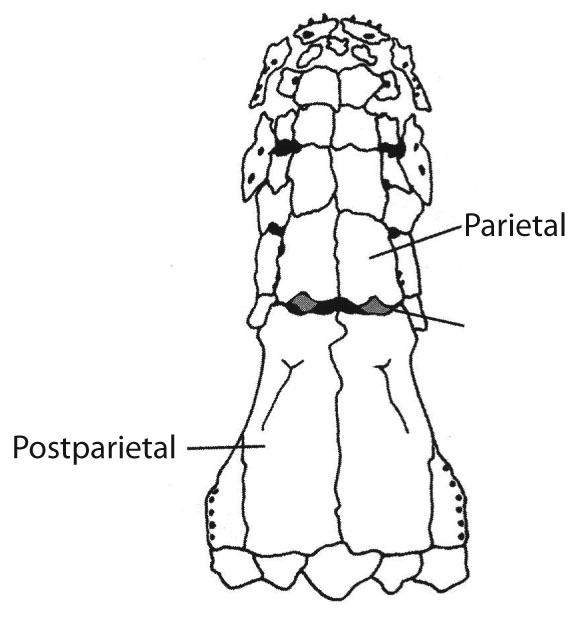
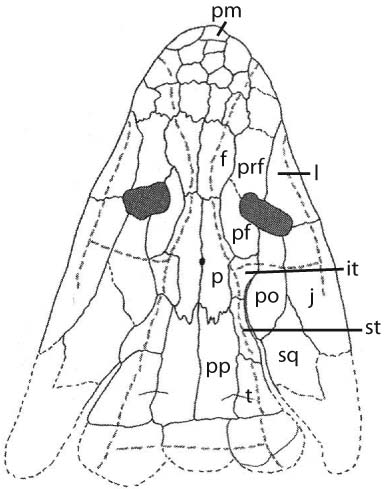
Skull roof of the coelacanth Rhabdoderma
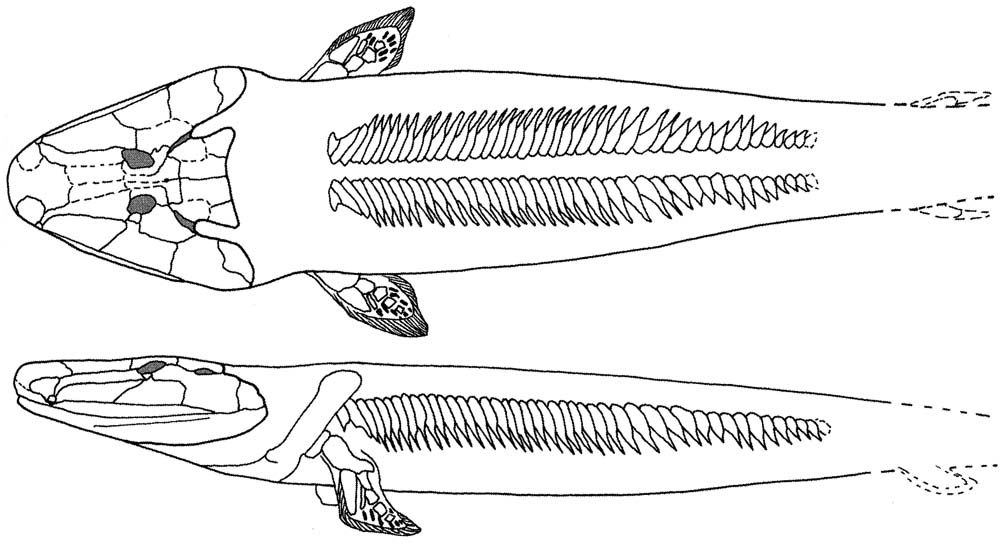
Onychodontiformes include a small number of taxa including Onychodus (above right) and Strunius. Their synapomorphy:
- Prominent parasymphyseal tooth whorls that could be protracted when the jaw was opened, and significantly modified palatal bones to accommodate them.

Latimeria menadoensis
- Discovered in 1938 by amateur ichthyologist and museum curator Marjorie Courtenay Latimer on the commercial fishing wharf of East London, South Africa. Despite great effort, another specimen was not found until 1952, off the Comoros Islands. Apparently the South African specimen had been way out of range. In the Comoros, it was known to local anglers. Recently, a second population has been found in Indonesia.
- Maxilla lost.
Miguashaia
The actinistian record:

- True range is Late Devonian to recent, however fossils indicate extinction in Cretaceous.
- Two major periods of diversification and success:
- Paleozoic forms representing an initial diversity of morphologies including Miguashaia (above right), Allenypterus during the Devonian, ultimately stabilizing on the classic actinistian morphology during the Carboniferous. (E. G. Rhabdoderma). This diversity was pruned by the Permo-Triassic extinction.
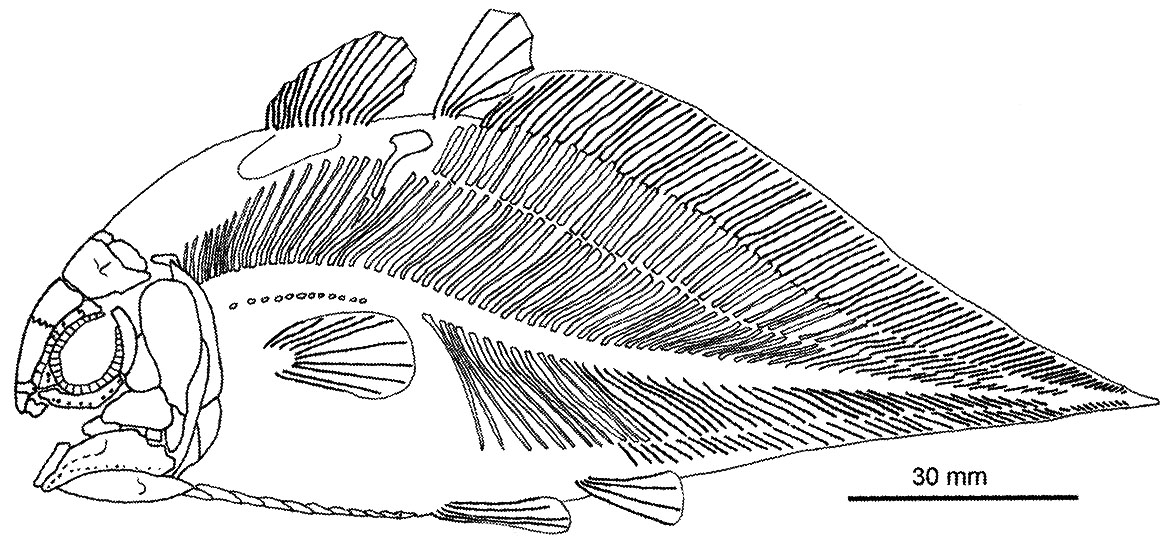
- Mesozoic radiation including ambush predator Rebellatrix (Triassic), Axelrodichthys (right), the giant Mawsonia.
Actinistian biology: Latimeria tells us much about the biology and ecology of ancestral sarcopterygians.
- It is the only sarcopterygian to retain the bipartite neurocranium that diagnoses Sarcopterygii ancestrally. In it, muscles on the ventral side connect the two parts of the neurocranium. When they contract, the anterior section is flexed downward, inreasing the power of the animal's bite.
- The actinistian lung is variously modified. In Latimeria, it contains fatty tissue. In some fossil forms, it possesses a calcified outer membrane.
- Latimeria is ovoviviparous: it practices internal fertilization and young are retained in the oviduct until highly developed, like in some sharks. Note, the young are nourished by yolk. There is no maternal nourishment.
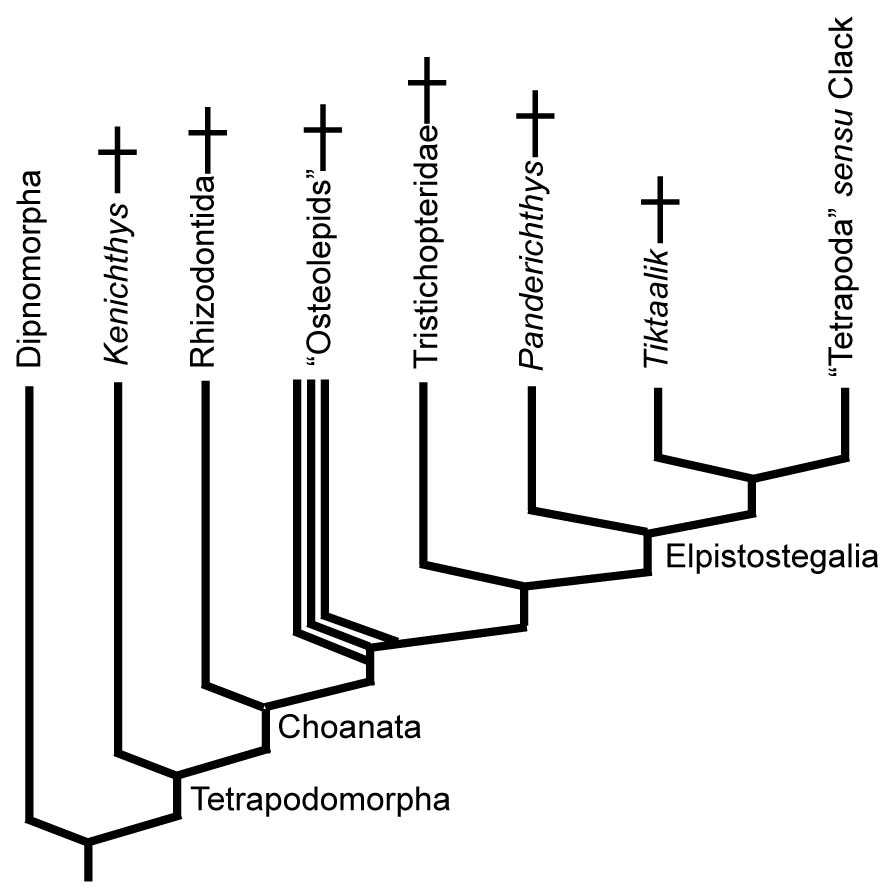
Dipnomorpha: (Devonian - Quaternary) Total group containing Dipnoi (lungfish) and their fossil relatives.

- Living lungfish and their adaptations: Today there are three genera of lungfish, one each for:
- Australia (Neoceratodus)
- Africa (Protopterus)
- South America (Lepidosiren)
Attributes of living lungfish:
- Maxillae and premaxillae lost.
- Mouths are modified for crushing hard-shelled prey. Teeth are consolidated into stout crushing plates.
- Holostylic jaw suspension with palatoquadrates fused to neurocranium.
- Both incurrent and excurrent nostrils open onto the palate.
- Long fleshy limbs.
- Reliance on air breathing: Lungfish's lungs have convoluted infoldings to increase surface area. Branchial arches are reduced in size. In fact, the African and South American varieties require air to survive.
- Aestivation: During periods of drought, lungfish burrow into the mud, secrete a cocoon of mucus, and enter a state of suspended animation. When their pool fills up, they awake and emerge. In fact, in the pet trade, lungfish are occasionally collected and shipped in this state - as genuine dehydrated pets. BTW, their adptations make them very hardy aquarium fish.

Ancient dipnomorphs and their fossil record:
- Lungfish similar to modern ones appear in the Triassic (Early Mesozoic).
- Fossil aestivation burrows are known from the Early Permian.
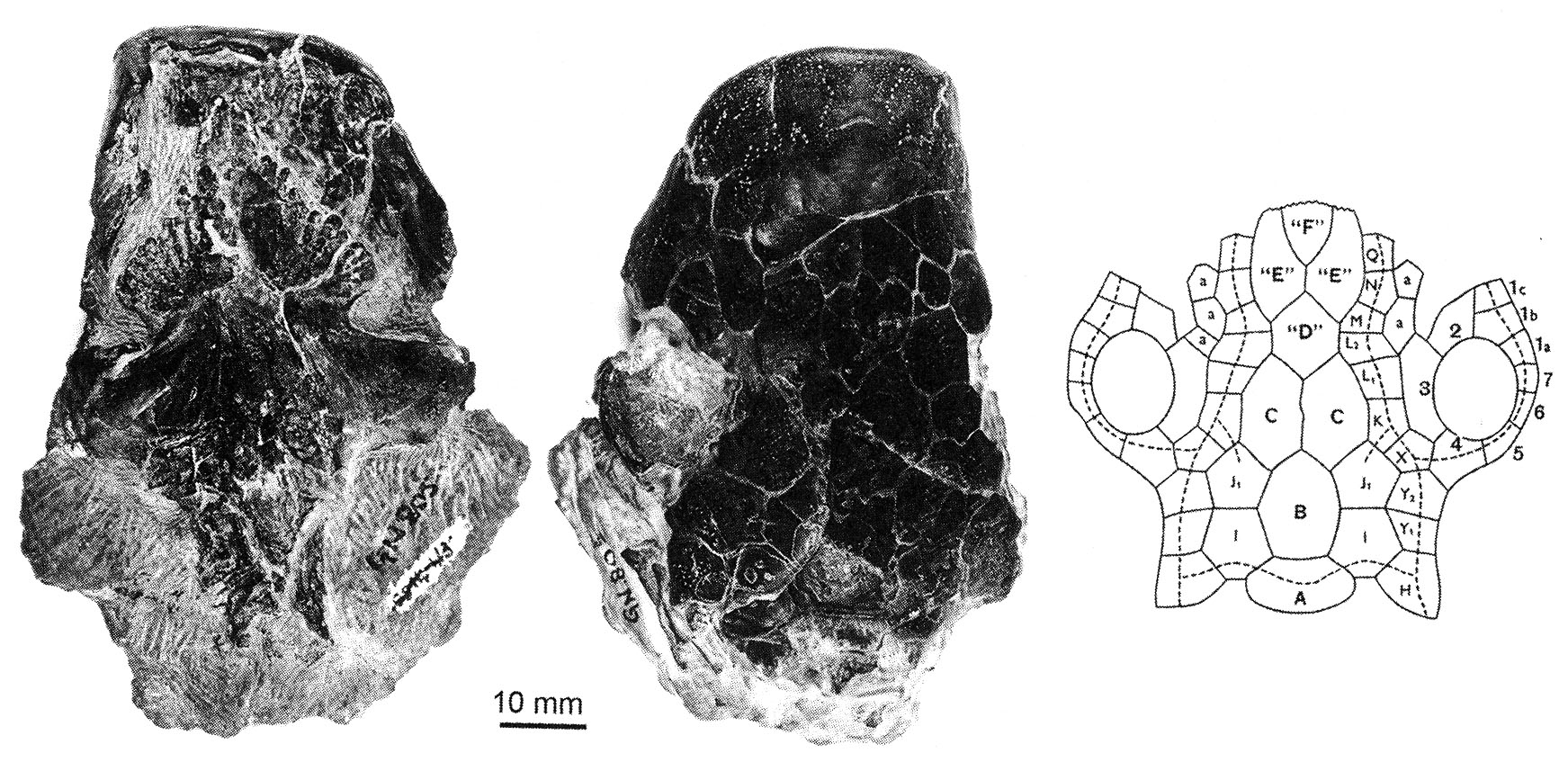
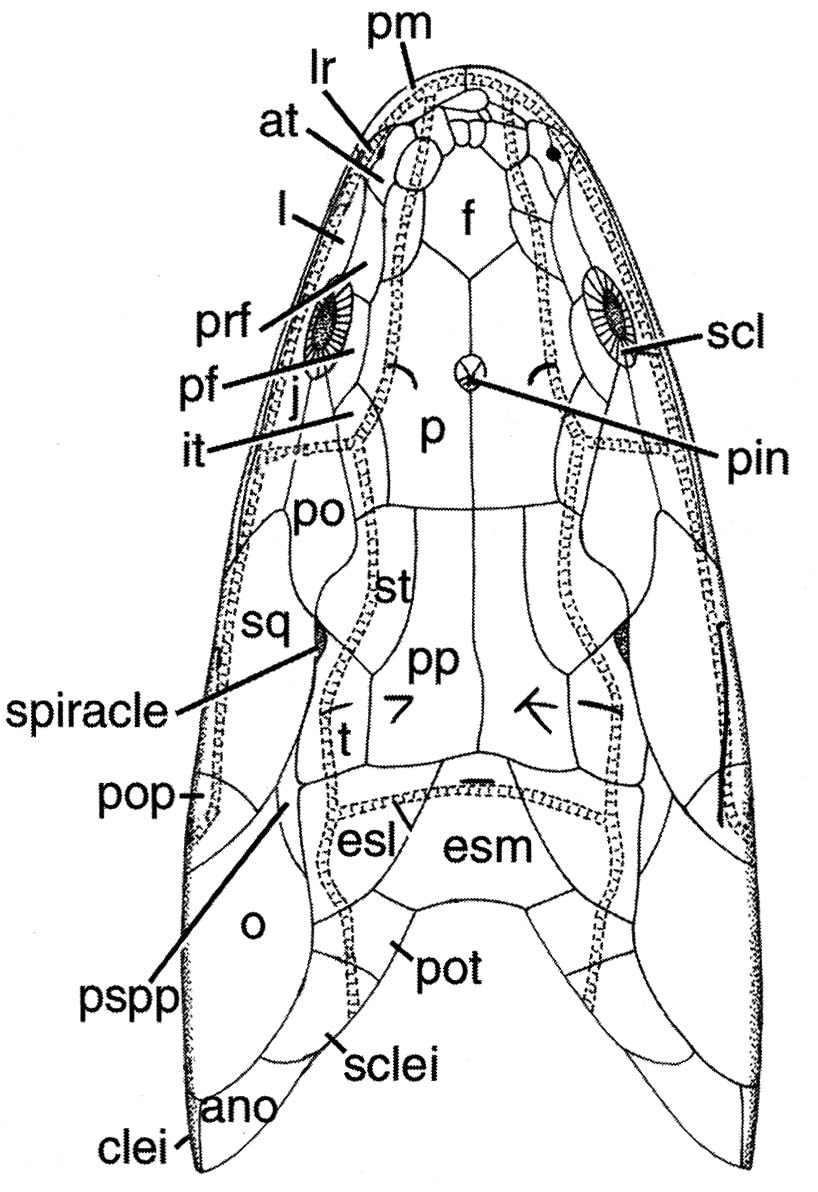
Skull of Dipterus in ventral and dorsal view, and schematic of dermal elements
- In the course of evolution, the recognizable dermal bones of their heads have fragmented into small unrecognizable units.
- Even the earliest lungfish have lost the separation of the two parts of the neurocranium, rendering even their status as sarcopterygians ambiguous at first.
- Spongy bone in snout containing many small tubules. Paradoxically, the synapomorphy of Dipnomorpha has been lost by its living members. This tissue probably contained the nerves of a sensory organ in the snout, and the blood vessels to support them.
- Long slender forelimb. Characterized by a metapterygial axis that does not branch into pre and postaxial elements.
Tetrapodomorpha
Tetrapodomorpha: All organisms more closely related to land vertebrates than to lungfish. These creatures didn't actually set out to become land animals. They were perfectly happy as fish. In fact, most of the evolutionary novelties that predisposed them toward life on land were near-term adaptations to life in water or were simply accidents. But first, what were they?
Our starting point is the completely aquatic Holoptychius, a porolepiform dipnomorph. Fully aquatic with external incurrent and excurrent nares. From such beginnings, we witness a series of transformations that give rise to creatures with at least the latent ability to invade the land.
What must happen to these creatures for them to be able to move about on land, even occasionally?
Roster of transformations: Becoming a land vertebrate is not easy and certainly not inevitable. Major adaptations are required in the following systems:
- Respiratory: Lungs aren't a problem. Inherited from the LCA of Osteichthyes. Gills are a problem. Out of water, they become a sure means of desiccation and must be reduced.
- Skeletal: On land, one must support one's weight against gravity and still be able to move. This requires:
- Robust paired limbs
- Digits
- Vertebral ossifications and articulations
- A sacrum
- The ability to support the heavy cantilever of the head from the pectoral girdle.
- Feeding: A fish has no neck. To pick food up off the ground, the head must bend with respect to the torso. So, a neck - strong enough to support the head, but also flexible.
- Sensory: All of the senses must be retooled to function in air.
- Olfaction: Olfactory epithelium must be kept moist.
- Vision: Refractive indices differ in air and water. Some form of blinking to keep the eye moist.
- Teleosts like the mudskipper simply "blink" by retracting their eyes into their sockets.
- Living amphibians continue to retract their eyes, but also use eyelids and nictitating membranes (Aiello et al., 2023).
- Among amniotes, eyelids and nictitating membranes take over the work.
- Lateral line sense: Useless out of water. Still, as an ancestral feature, they tend to be retained by larvae and adults that spend any significant amount of time in the water.
- Hearing: Air is much less dense than water. Thus, old structures must be radically
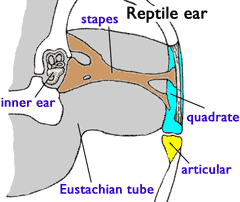
- Hearing: Acoustic impedance is the resistance of a system to the flow of sound through it. It is a function of the density of the medium. The density of the body is similar to that of water. Thus, sound can pass directly from water through the body into the otic capsule at full intensity. Air is much less dense than water. Thus, for airborn sound to register in the otic capsule, it must be collected and amplified. This is accomplished by an impedance matching ear:
- Sound is collected over a broad area by a tympanum (A.K.A. ear drum) or through the jaw.
- In land vertebrates, a rod-like bone called the stapes transmits the vibrations of the tympanum to a small opening in the otic capsule, the fenestra ovalis (oval window.) By being concentrated into a small area, the sound is amplified so that it can be detected by the inner ear.
- The stapes is the hyomandibula, modified from the transmission of sound. The air filled space it occupies is called the middle ear. Together with the eustacean tube that connects the middle ear to the pharynx, it is homologous to the spiracle of aquatic vertebrates.
- The fenestra ovalis is the ancestral vestibular fontanelle, the ventral extremity of the otico-occipital fissure.
- The tympanum represents the developmental failure of the spiracle to penetrate the surface of the body. Instead, a thin membrane is stretched across the spiracular notch.
The transformation of the hyomandibula into a stapes is gradual, and in some creatures, the structure probably served both load-bearing and acoustic functions. The transformation of the spiracle into an otic notch for the tympanum has no direct fossil record. When a notch is present at the posterior cheek margin, we rely on inference. We assume:
- Acanthostega's stapes was short and massive, but rather than articulating with an articular face in the neurocranium, it's head fit into the vestibular fontanelle.
- Creatures like Crassigyrinus with heavy stapes and extensive lateral line systems might have retained an open spiracle.
- Creatures like Greererpeton with no notch might have no external spiracle or tympanum at all.
- Creatures with reduced lateral line systems and slender stapes probably had tympana.
As we will see, the impedance-matching ear evolved slowly and independently several times in different lineages.
- Sound is collected over a broad area by a tympanum (A.K.A. ear drum) or through the jaw.
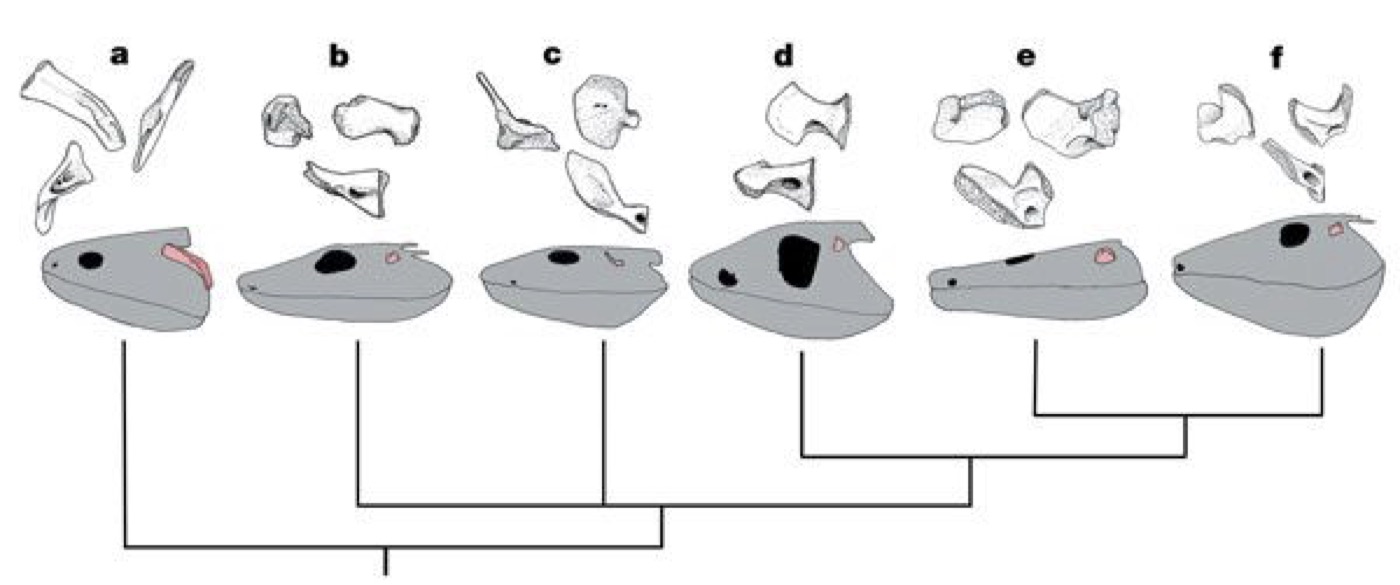
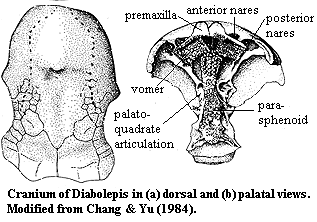
The choana: Most aquatic osteichthyans get air into their lungs by gulping and swallowing it. In land vertebrates, however, air flows through the nares into the nasal capsulethen through am "internal naris" or choana into the oral chamber. In effect, the excurrent naris of other aquatic osteichthyans has been shifted across the upper lip. Kenichthys campbelli (Early Devonian), the basal member of Tetrapodomorpha, shows this transition in progress:
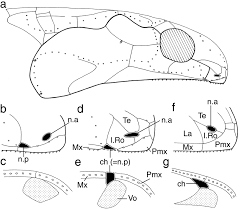
- b, c: Youngolepis, a dipnoan with both nares external
- a, d, e: Kenichthys, with the excurrent naris embedded in the upper lip.
- f, g: Eusthenopteron, a derived tetrapodomorph with external naris and internal choana.
- Choanae present.
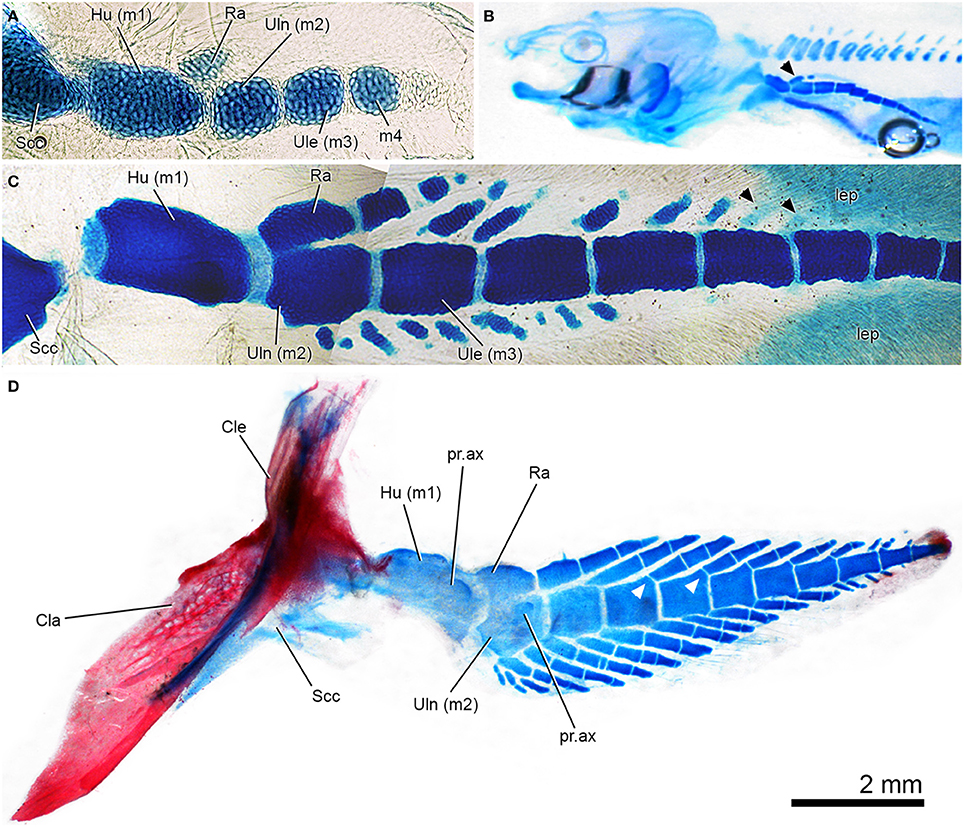
- Paired fins with two radial elements (radius and ulna in pectoral fin, tibia and fibula in pelvic). (Note: in Kenichthys the condition is unknown. According to Jude et al. 2014, the living dipnoan Neoceratodus briefly shows this condition as an embryo, but the radials soon fuse into a single element.
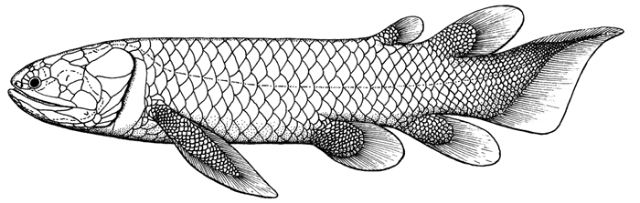
Rhizodontida: (Devonian - Carboniferous) Tetrapodomorph superlatives. Fresh water ambush predators including Rhizodus (right) which at up to 7 m may be the largest fresh water fish ever. Characterized by:

- Both dorsal fins and anal fin displaced posteriorly into the "ambush predator" ecomorph.
- Fins reinforced for strength with unsegmented lepidotrichia and a superficial layer of scales. Did they use their reinforced pectoral fins partially to support their weight or to push their heads above water?
- Complex lateral line system. (On taxon has three parallel lateral lines.)
"Osteolepids" (Devonian - Permian) A speciose paraphyletic assemblage of aquatic sarcopterygians. The synapomorphies that unite them with tetrapods to the exclusion of rhizodontids are highly technical. For us, they provide a picture of aquatic choanates that are known from long and thorough study.
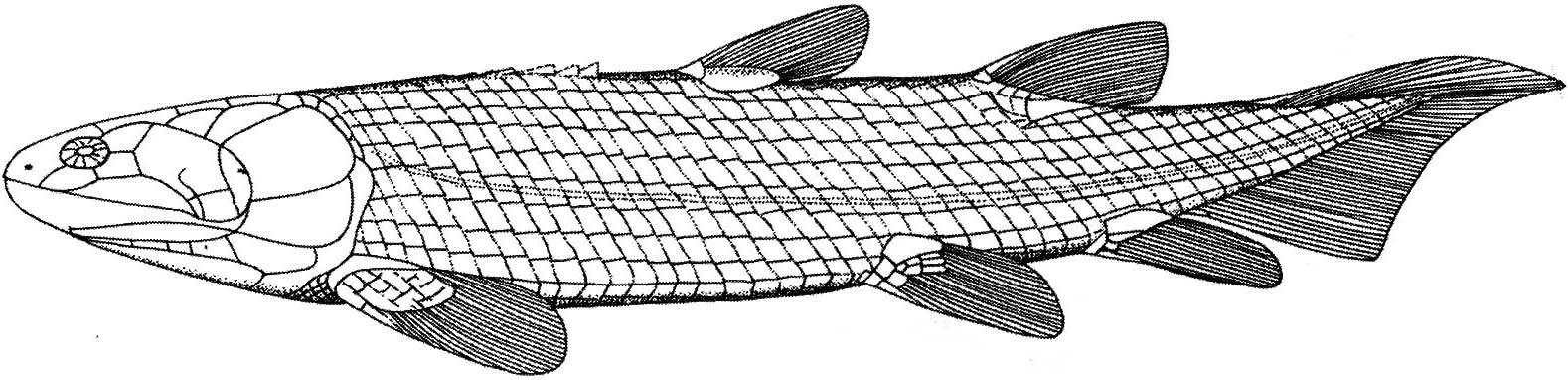
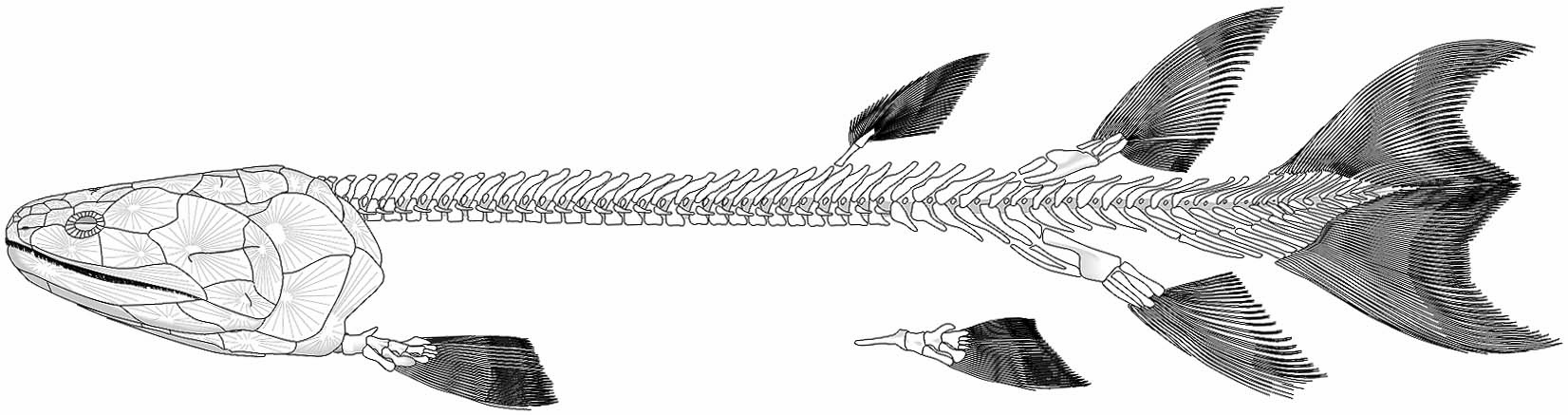
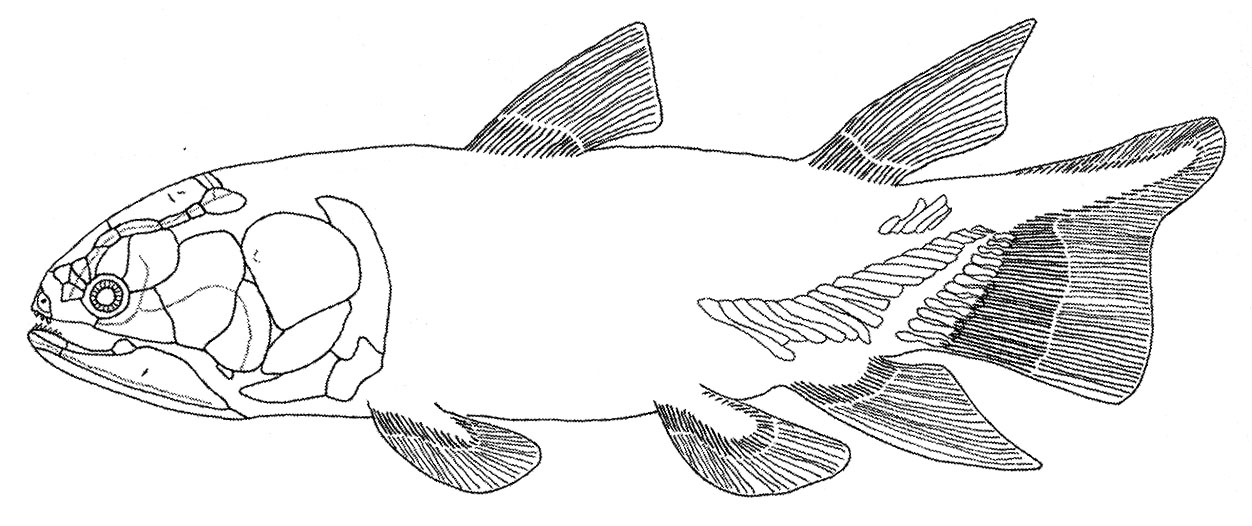
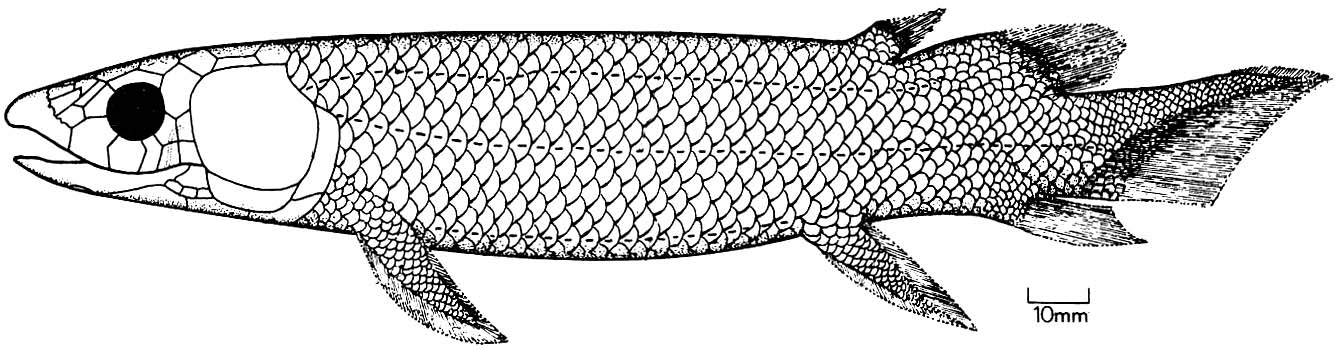
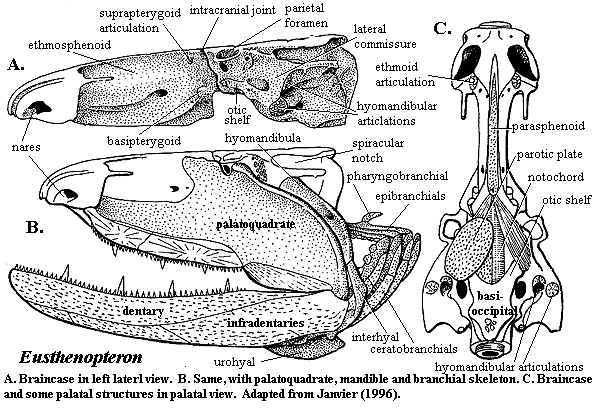
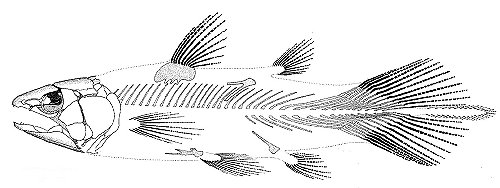
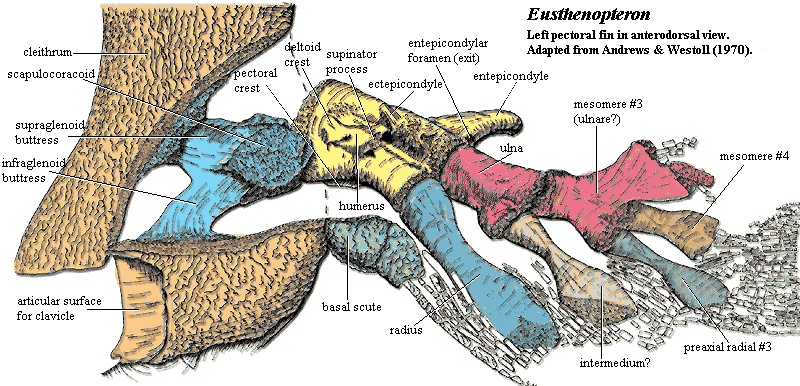
Eusthenopteron foordi
Synapomorphies of Trstichopteridae include:
- The presence of a postspiracular bone in the upper cheek margin.
- A three-lobed caudal fin and caudal skeleton reinforced by elongate neural spines dorsally and accessory epihaemal spines ventrally.
- Absence of cosmine from scales.
Synapomorphy of Tristichopteridae and Tetrapoda:
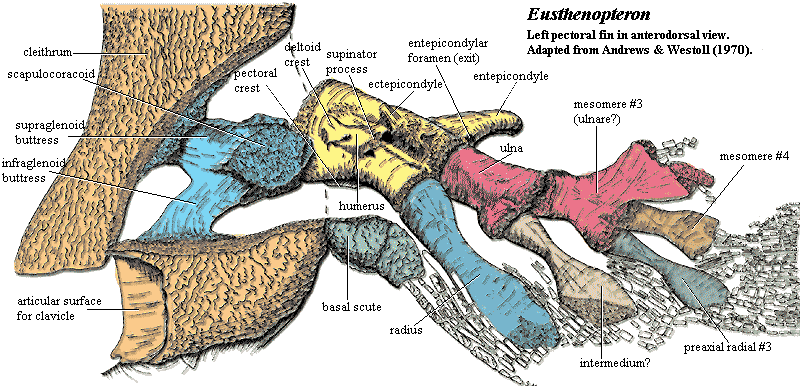
- A humerus with recognizable entepicondyle and ectepicondyle. (Regions for muscle insertions.)
Elpistostegalia
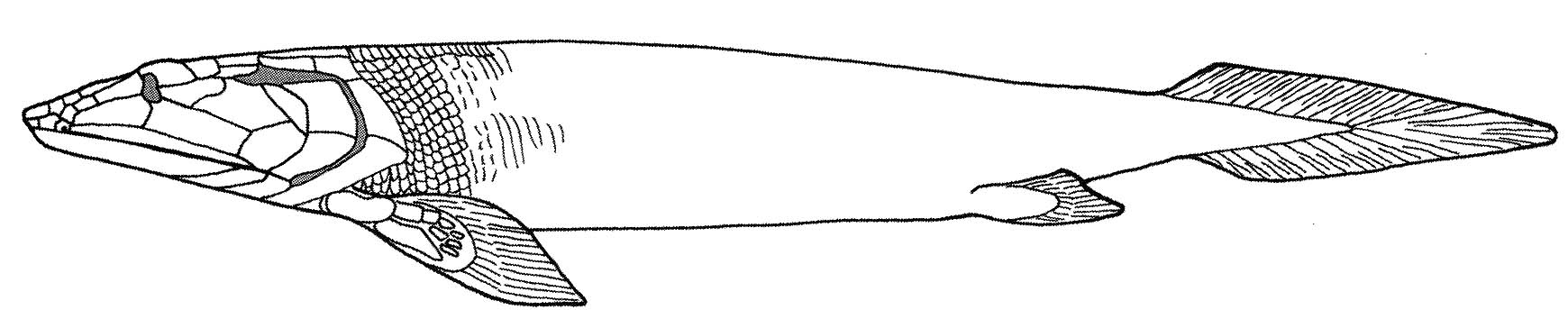
The first true indication of a tendency to place at least parts of the body out of the water occurs in Elpistostegalia (Latest Devonian - Quaternary - named for a poorly known member, Elpistostege.) The last common ancestor of Panderichthys and Tetrapoda and descendants. Members of this clade, from basal clades, up through the LCA of Tetrapoda inhabited a combination of brackish and fresh water. (Laurin, 2024) The first and most basal well-known member is....
Panderichthys (Late Devonian) Known from the Baltic region. Up to over a meter length, it is slightly flattened with a pointed snout. Its orbits are located on the dorsal surface of the skull, as are the spiracles, as if they were meant to project above the water. The impression given is of an aquatic animal specialized for shallow water and for hunting creatures just above the water's surface. Panderichthys retains rather heavy scales.
Synapomorphies of Elpistostegalia and Tetrapoda:
- Skull flattened with orbits and spiracles dorsal
- Paired frontals
- The intracranial joint between parietals and postparietals sutured and immobile (even though basal forms like Panderichthy retain the sarcopterygian bipartite neurocranium. Compare with Eusthenopteron)
- Large spiracles and spiracular chambers
- Loss of dorsal and anal fins
Tiktaalik roseae: (Late Devonian) Described in 2006 by Daeschler et al., this creature has become the current poster-child of vertebrate evolution. It is featured in:

- A new popular book
- The Colbert Report
- A Southpark episode (rather vulgar. Search for "retard baby fish.")
In its general profile, it's similar to Panderichthys, there are two important differences.
- The wrist. Although fin rays are still present (and digits absent), its wrist is capable of flexing. Thus, it could support at least some of its weight on its forelimbs. No indication that it ever emerged completely from the water.
- The neck: Tiktaalik lacks:
- opercular and subopercular. (The preopercular remains)
- all dermal elements of the pectoral girdle dorsal to the cleithrum.
As a result, its skull and pectoral girdle were no longer in contact, allowing the head to be flexed with respect to the torso - the beginnings of the neck.
- Tiktaalik retained heavy scales that must have sustained considerable mechanical load.
- Tiktaalik's pelvis was described by Shubin et al., 2014. Although the hindlimb is unknown, the pelvis is surprisingly large and the hip socket is circular, indicating that the hindlimb was strong and had a wide range of motion. Tiktaalik's vertebral column and hindlimb has recently been described by Stewart et al., 2023 based on new preparation of a specimen. Typically, the pectoral fins of sarcopterygians are larger, however the hindlimb (pelvic fin) of Tiktaalik approaches the size of the pectoral fin. Its vertebral column preserves neural arches and intercentra. Stewart et al. identify distinct sacral ribs that are broad and downturned, but which lack a bony articulation with the ilium. Almost a sacrum!
From this it seems that Tiktaalik could support at least a little of its weight on its paired fins. What would be the point? For an idea, we look at the head.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
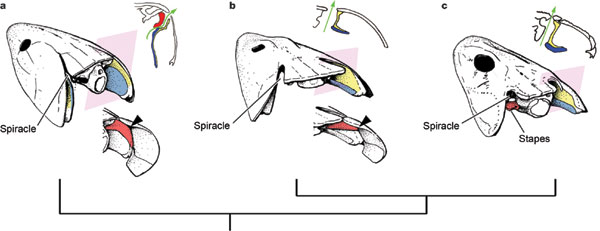
Spiracles of a tristichopterid and a basal elpistostegalian compared from Brazeau and Ahlberg, 2006.
- Open
- Large
- Positioned on the upper surface of the skull
But note that there is a positive-feedback effect here: Modern fish who rely on air-breathing in hypoxic water environments tend to reduce their gills in order to avoid losing oxygen from the blood through outward diffusion, making them more dependent on air-breathing. Why not quickly lose gills altogether? Because it is metabolically easier to excrete CO2 through them than through the lungs because CO2 shed into water is quickly converted to HCO3-, maintaining a strong CO2 concentration gradient ( Pelster, 2021) at the gills. As we will see, tetrapods held onto their gills for quite a while for this reason. (Witzmann, 2015)
- Brett R. Aiello, M. Saad Bhamla, Jeff Gau, and Thomas A. Stewart. 2023. The origin of blinking in both mudskippe. 120 (18) e2220404120
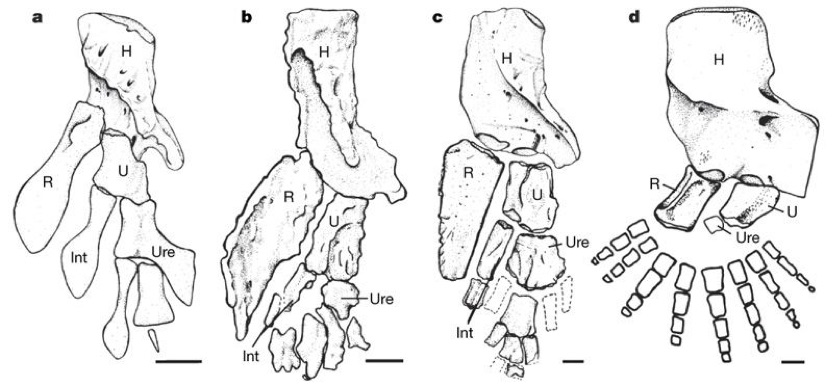
- Catherine Boisvert, Elga Mark-Kurik, and Per Ahlberg, 2008. The pectoral fin of Panderichthys and the origin of digits. Nature 456, 636-638
- Martin D. Brazeau1 & Per E. Ahlberg1, 2006. Tetrapod-like middle ear architecture in a Devonian fish. Nature 439, 318-321
- Robert Carroll, 2009. The Rise of Amphibians: 365 Million Years of Evolution. Johns Hopkins University Press, Baltimore. 360 pp.
- María Ciudad Real, Jorge Mondejar Fernandez, Daniel Vidal, and Hector Botella. 2020. Is Onychodontida (Osteichthyes, Sarcopterygii) monophyletic? Assessing discordant phylogenies with quantitative comparative cladistics. Spanish Journal of Palaeontology. 537(1): 87–100.
- Jennifer Clack, 2007. Devonian climate change, breathing, and the origin of the tetrapod stem group. Integrative and Comparative Biology, 47 (4): 510-523.
- Jennifer Clack, 2014. Gaining Ground: The Origin and Evolution of Tetrapods. Indiana University Press, Bloomington. 523 pp.
- Jennifer Clack. 2012. A Carboniferous embolomere tail with supraneural radials. Journal of Vertebrate Paleontology, 31(5) pp. 1150-1153.
- Alice M Clement, Benedict King, Sam Giles, Brian Choo, Per E Ahlberg, Gavin C Young, John A Long, 2018. Neurocranial anatomy of an enigmatic Early Devonian fish sheds light on early osteichthyan evolution. eLife May 29, 2018.
- Michael Coates, Marcello Ruta, and Matt Friedman. 2008. Ever Since Owen: Changing Perspectives on the Early Evolution of Tetrapods. Annual Review of Ecology, Evolution, and Systematics. 2008. 39:571–92.
- Edward Daeschler, Neil Shubin, Farish Jenkins, Jr. 2006. A Devonian tetrapod-like fish and the evolution of the tetrapod body plan. Nature 440, 757-763
- Emma Jude, Zerina Johanson, Anton Kearsley, and Matt Friedman, 2014. Early evolution of the lungfish pectoral-fin endoskeleton: evidence from the Middle Devonian (Givetian) Pentlandia macroptera. Frontiers in Earth Science 12
- Jozef Klembara, Jennifer Clack, Andrew Milner, and Marcello Ruta. 2014. Cranial anatomy, ontogeny, and relationships of the Late Carboniferous tetrapod Gephyrostegus bohemicus Jaekel, 1902. Journal of Vertebrate Paleontology, 34(4):774–792.
- Michel Laurin. 2024. Habitat of early stegocephalians (Chordata, Vertebrata, Sarcopterygii): a little saltier than most paleontologists like?. Fossil Record, 27(3):299–332.
- Grzegorz Niedzwiedski, Piotr Szrek, Katarzyna Narkiewicz, Marek Narkiewicz, and Per Ahlberg. 2010. Tetrapod trackways from the early Middle Devonian period of Poland. Nature 463, 43-48
- Jason Pardo. 2024. New information on the neurocranium of Archeria crassidisca and the relationships of the Embolomeri. Journal of experimental Zoology 201(3)
- Bernd Pelster (2021) Using the swimbladder as a respiratory organ and/or a buoyancy structure—Benefits and consequences. Journal of experimental Zoology Part A 335(9-10):831-842.
- Tuo Qiao and Min Zhu. 2010. Cranial morphology of the Silurian sarcopterygian Guiyu oneiros (Gnathostomata: Osteichthyes). Science China Earth Sciences. 53, (12) 1836-1848.
- Neil Shubin, Edward Daeschler, Farish Jenkins, Jr., 2014. Pelvic girdle and fin of Tiktaalik roseae. Proceedings of the National Academy of Sciences. 111(3) 893-899
- NT.A. Stewart, J.B. Lemberg, E.J. Hillan, I. Magallanes, E.B. Daeschler, N.H. Shubin 2023. Axial regionalization in Tiktaalik roseae and the origin of quadrupedal 2 locomotion. BioRxiv doi: https://doi.org/10.1101/2023.01.11.523301
- Florian Witzmann. 2015. CO2-metabolism in early tetrapods revisited: inferences from osteological correlates of gills, skin and lung ventilation in the fossil record. Lethaia 49(4), 492–506.
- Min Zhu and Per Ahlberg, 2004. The origin of the internal nostril of tetrapods. Nature 432, 94-97
- Catherine Boisvert, Elga Mark-Kurik, and Per Ahlberg, 2008. The pectoral fin of Panderichthys and the origin of digits. Nature 456, 636-638