Lepidosauromorpha


Spencer's monitor Varanus spenceri a squamate; the tuatara Sphenodon punctatus a rhynchocephalian.
Lepidosauromorpha: (Triassic - Rec.) All organisms more closely related to lepidosaurs than to archosaurs. Max ∇ (Lepidosauria ~ Archosauria). Membership includes:
- Lepidosauria proper:
- Squamata: Min ∇ (Gekkota, Anguimorpha, Mosasauroidea, Lacertidae, Serpentes)
- Rhynchocephalia: Min ∇ (Gephyrosaurus, Sphenodontida)
- Problematic possible stem-lepidosaurs.
Stem Lepidosauria
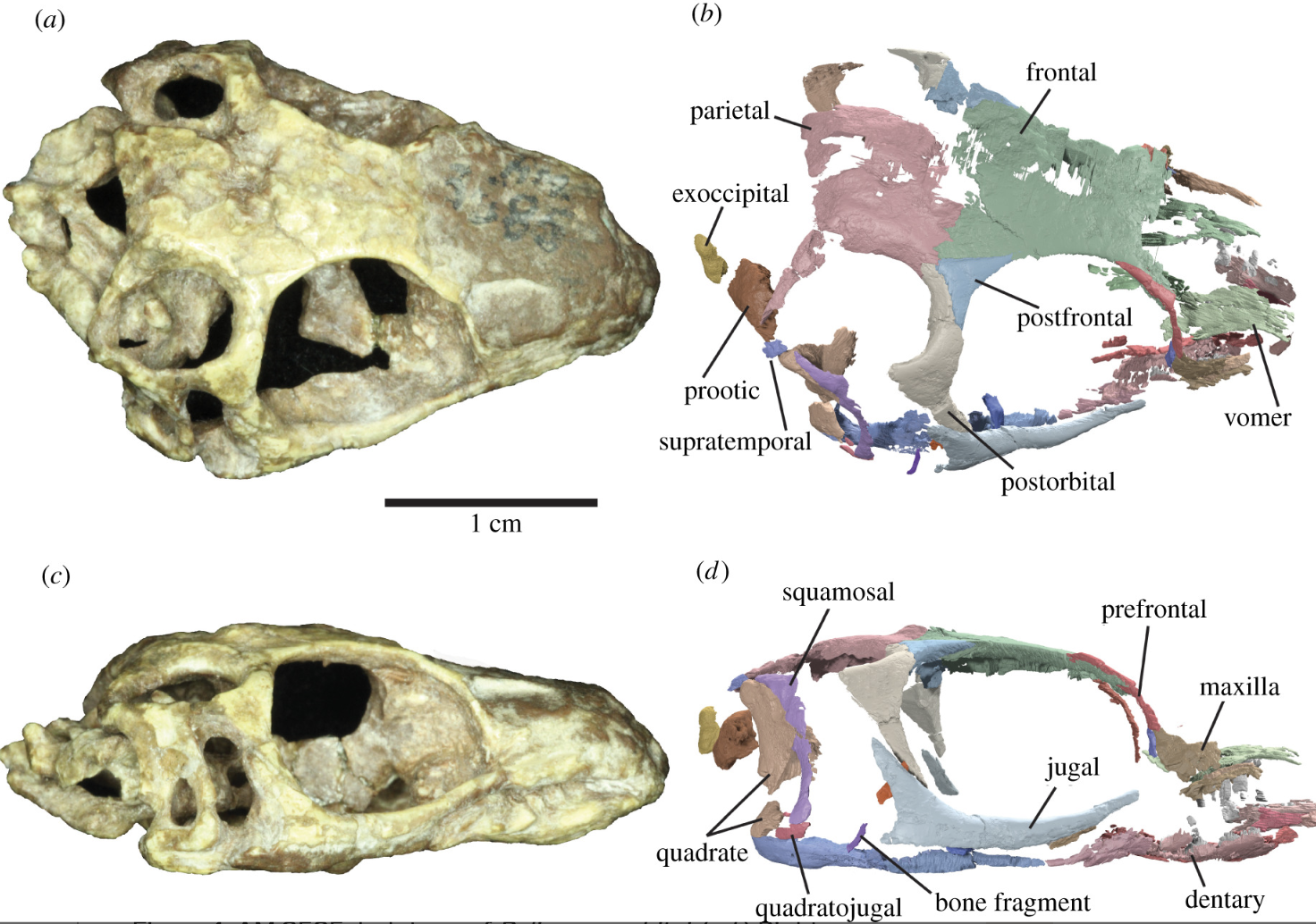
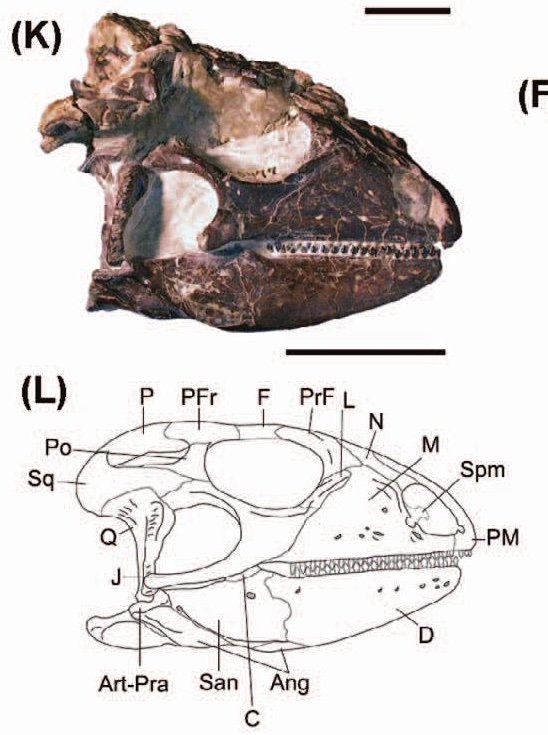
The membership of stem-Lepidosauria has been historically unstable. As of 2018, most of the fossil taxa placed on the lepidosauromorph stem have been evicted to other (and varying) parts of the diapsid tree (Simões et al., 2018, Ford et al., 2021) leaving only a handful of fragmentary potential stem lepidosauromorphs. Not really surprising given the fragmentary nature of these small animals' remains. In this part of the tree, we dine on scraps.The earliest reliable lepidosauromorph: The Early Triassic Paliguana whitei, recently redescribed by Ford et al., 2021 and known from a single skull. Broadly resembles the similar-sized stem saurians, but with potential synapomorphies with Lepidosauria:
- Greatly reduced lacrimal
- Fused frontals
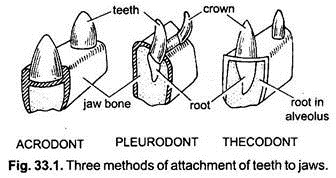
- Teeth set in groove with labial/buccal wall significantly taller than lingual wall - a condition approaching the pleurodont dentition of living squamates.
- Tataylura's frontals are paired
- It's teeth are set in grooves, but the lingual walls are not reduced.
Sophineta, a stem lepidosauromorph.
Lepidosaur Polytomy
Lepidosauria: (Late Triassic - Rec.) The last common ancestor of Squamata and Rhynchocephalia, and all of its descendants. The earliest fossils of each are Late Triassic in age, but calibrated molecular clocks indicate a divergence right around the Permo-Triassic extinction event. In our consensus cladogram, we follow Ford et al., 2021 in placing Squamata and Rhynchocephalia in a polytomy with:
- Sophineta cracoviensis (Early Triassic) (Evans and Borsuk-Bialynicka, 2009)
- Megachirella wachtleri (Middle Triassic) (Simões et al., 2018)
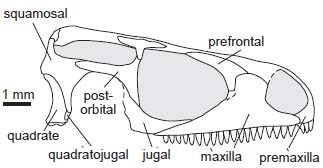
- The lacrimal becomes small and is crowded by the maxilla, which has a tall dorsal process and broadly enters the orbit margin.
- The quadratojugal becomes small
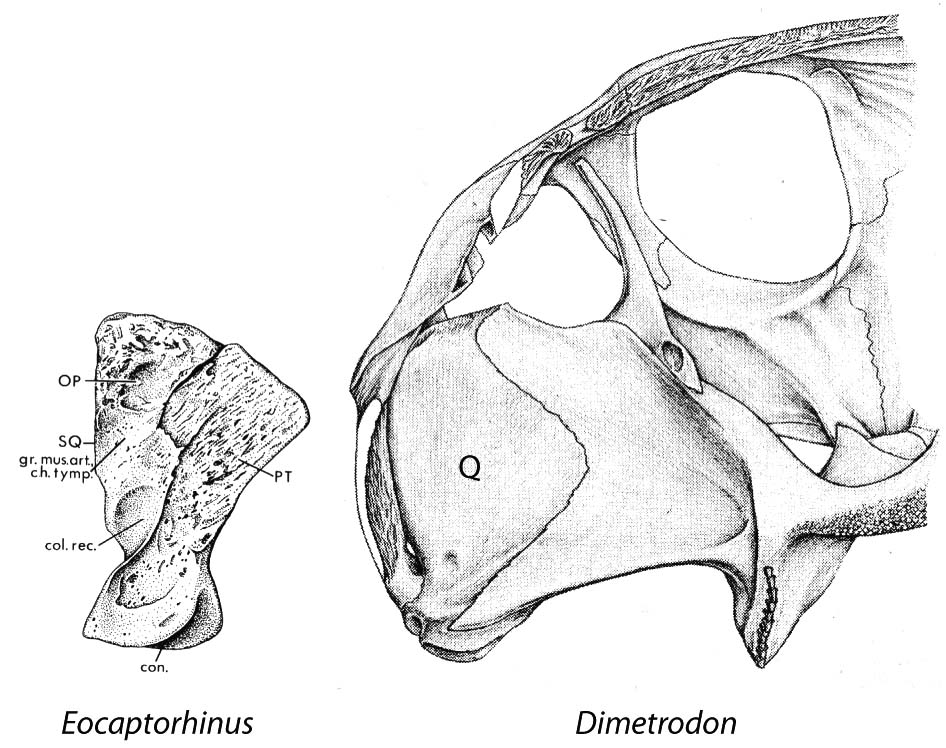
- The area of overlap between the quadrate and pterygoid is greatly reduced. (Compare Dimetrodon to Tupinambis, a squamate.)
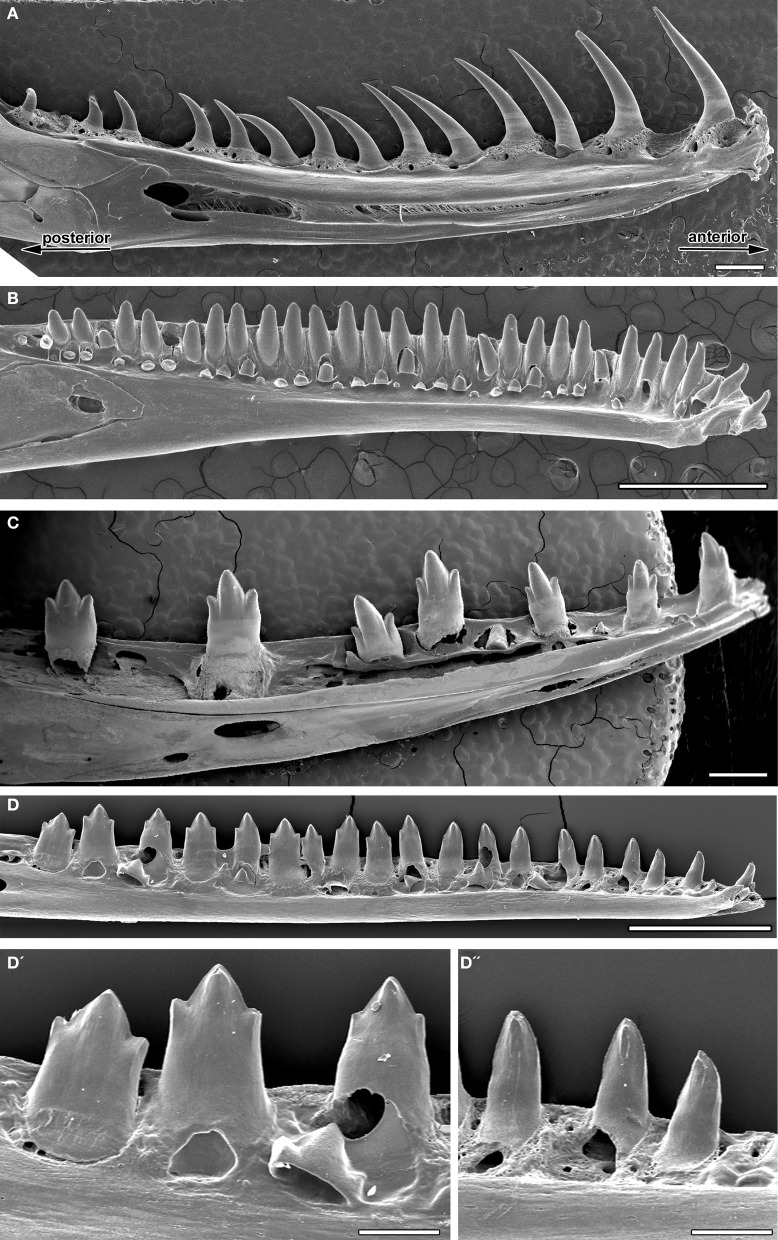
- The teeth become fully pleurodont - i.e. rather than being set in shallow sockets (subthecodont) they occupy a shelf and are buttressed laterally by bone.(Link to Sophineta maxilla.)
Lepidosauria: (Triassic - Quaternary)
Synapomorphies:- The quadrate flares laterally into a conch for support of the tympanum
- Keratinous overlapping scales: (contrast mojave rattlesnakes with false gavial, an archosaur)
- Modified mid-dorsal scale row.
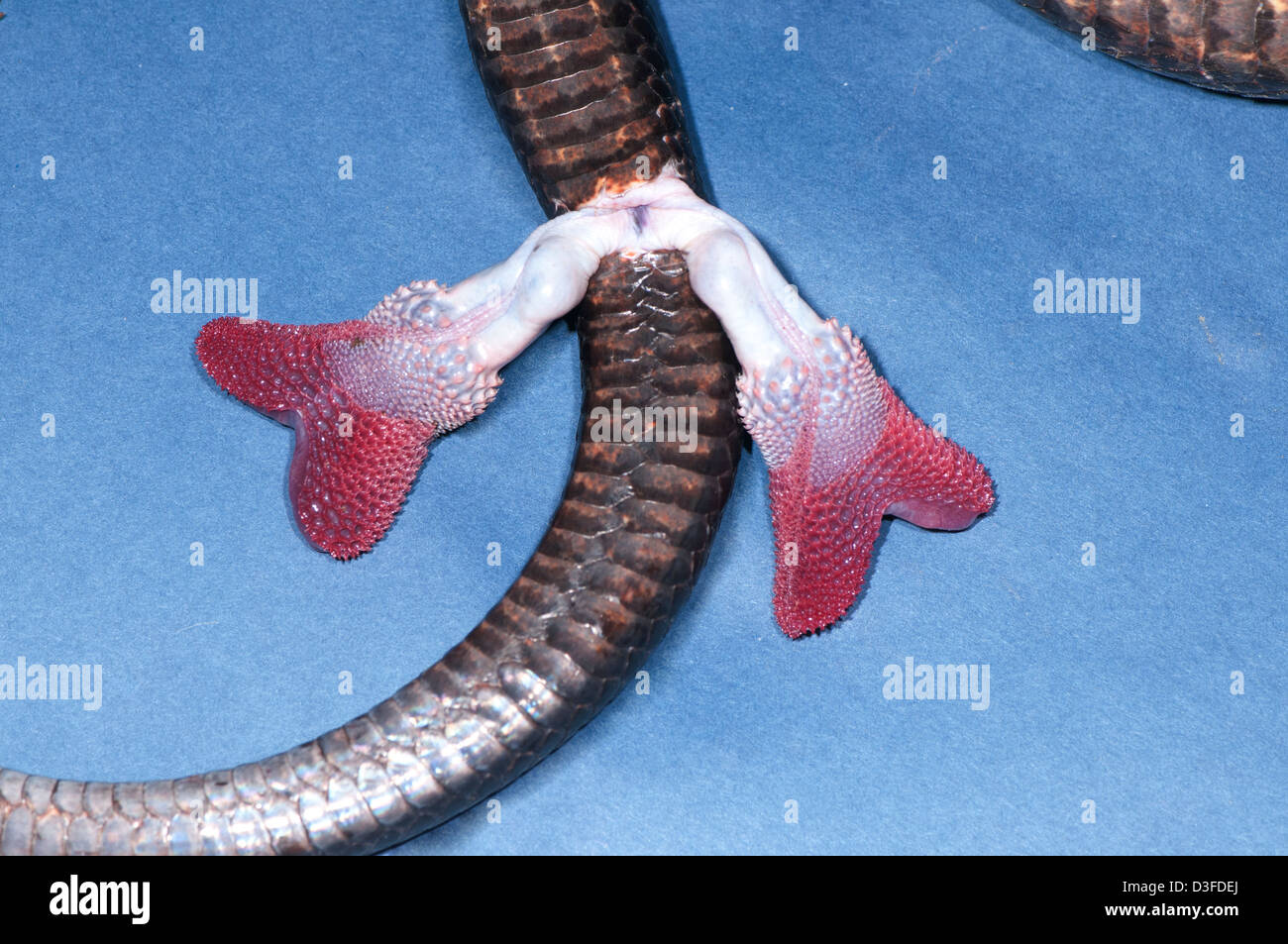
- Paired hemipenis (pl: hemipenes): Unique intromittant organs derived from midline amniote penis. As of 2022, also paired hemiclitores in snakes at least. (Folwell, et al., 2022)
- Caudal autotomy is commonly used. Autotomy planes are first observed in Captorhinus in the amniote stem, so are probably plesiomorphic for lepidosaurs, but the extensive use of caudal autotomy as an escape strategy characterizes Lepidosauria. (Note: this feature is lost among some derived members.)
- The mandibular glenoid (the articular surface of the jaw) is formed exclusively from the articular. (In other saurians, the dermal angular contributes.)
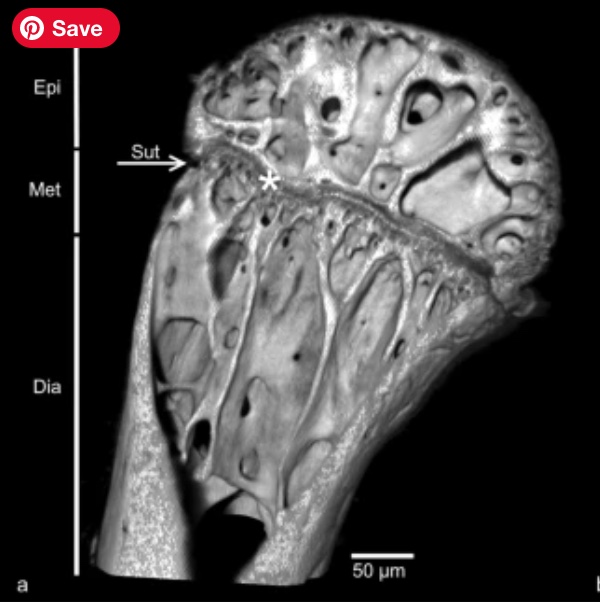
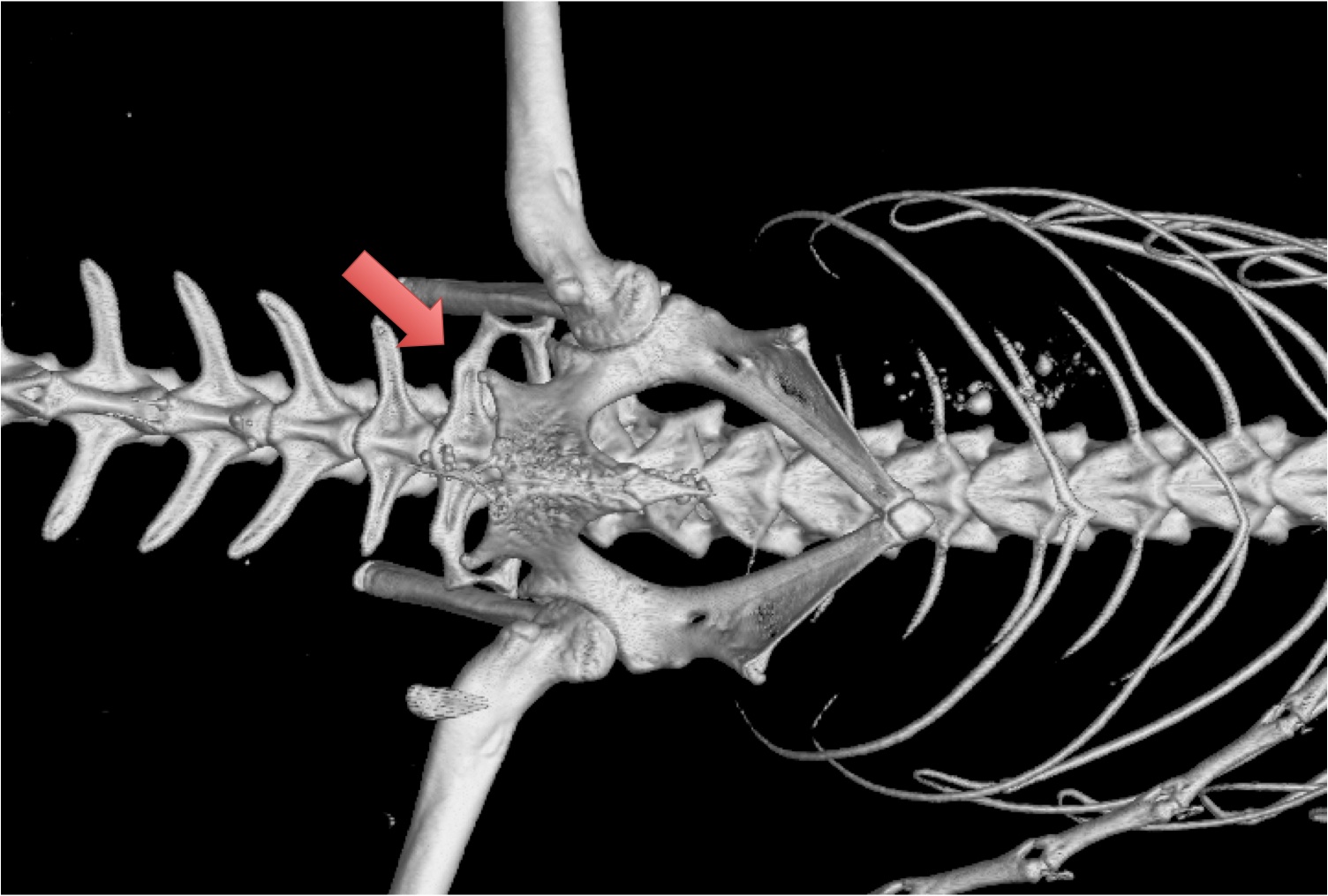
- Long bones feature distinct epiphyses and diaphyses separated by cartilaginous epiphyseal plates (Link to CT scan of Uromastix epiphyseal plate. Epiphyses and diaphyses fuse as the epiphyseal plate is resorbed upon full adulthood. Convergent with Mammalia. This feature typically indicated determinate growth. Indeed, Frydlova et al. 2020 regard determinate growth as plesiomorphic for Squamata.
Rhynchocephalia:
(Triassic - Quaternary) Total group - everything closer to Sphenodon than to Squamata. The single genus Sphenodon, the New Zealand tuatara is the sad remnant of a large group of lepidosaurs that was abundant and speciose during the Triassic and Jurassic. Ancestrally they were terrestrial, and basal forms like Gephyrosaurus (Early Jurassic Evans, 1980) were barely distinct from stem lepidosauromorphs and yet they are distinct in having: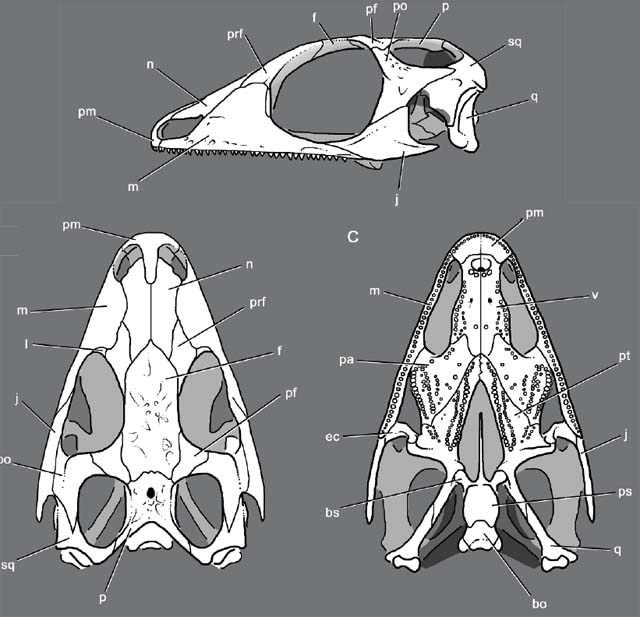
- A posterior process of the dentary that laps over the angular
- Loss of the splenial
- Supratemporal lost
- Organization of palatine teeth into rows that parallel marginal teeth
- The differentiation of the tooth row
- Acrodont dentition, in which teeth are fused to the bone of the jaw
- Transformation of premaxillary teeth into chisel-shaped incisors.
- Loss of the lacrimal
- The re-evolution, in some, of a fully ossified lower temporal bar.
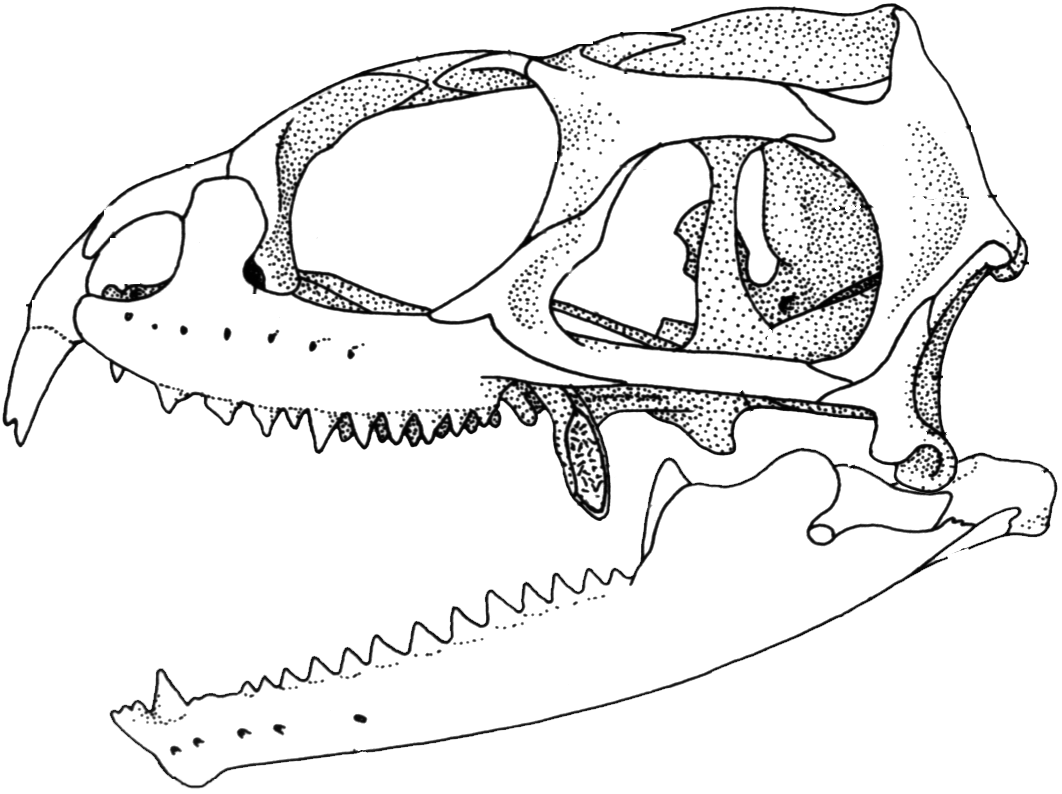
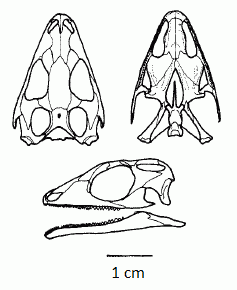
Sphenodon punctatus
- Pleurosauridae: (Jurassic) aquatic, with long, eel-like bodies and short limbs.
- Sphenodontidae: (Early Jurassic - Quaternary - right) The living Sphenodon and its closest fossil relatives are characterized by a return to the strong, slow bite we saw in ancestral amniotes.
- Sphenodon:(rec) lives on a few islands off the shore of New Zealand. It feeds on small vertebrates (which it bites pieces off of). Noteworthy for being able to remain active at low temperatures (roughly 50 degrees) thus being literally cold-blooded.
- Eilenodontinae: (Late Triassic - Late Cretaceous) Specialized herbivores, including the largest terrestrial rhynchocephalian on record Priosphenodon (Late Cretaceous), reached body lengths of one meter.
Despite their derived features, rhynchocephalians are a conservative group that, in many aspects of their biology, resemble stem-lepidosaurs.
Squamata:
Lizards and snakes. (Late Triassic - Rec.) Thousands of species. Many interesting adaptations. There is no way to cover exhaustively if we had an entire semester.
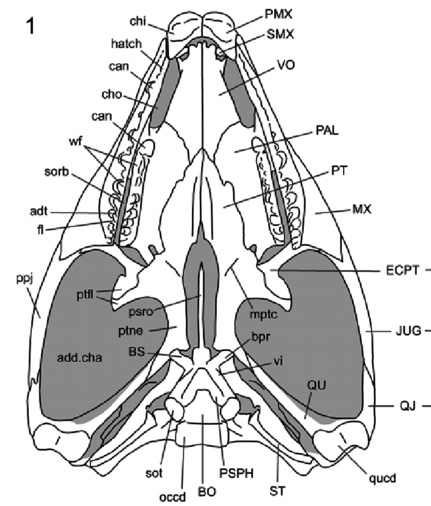
Sphenodon punctatus a rhynchocephalian compared to Tupinambis teguixin, a squamate.
The synapomorphies of Squamata are legion. We focus on a conspicuous subset:
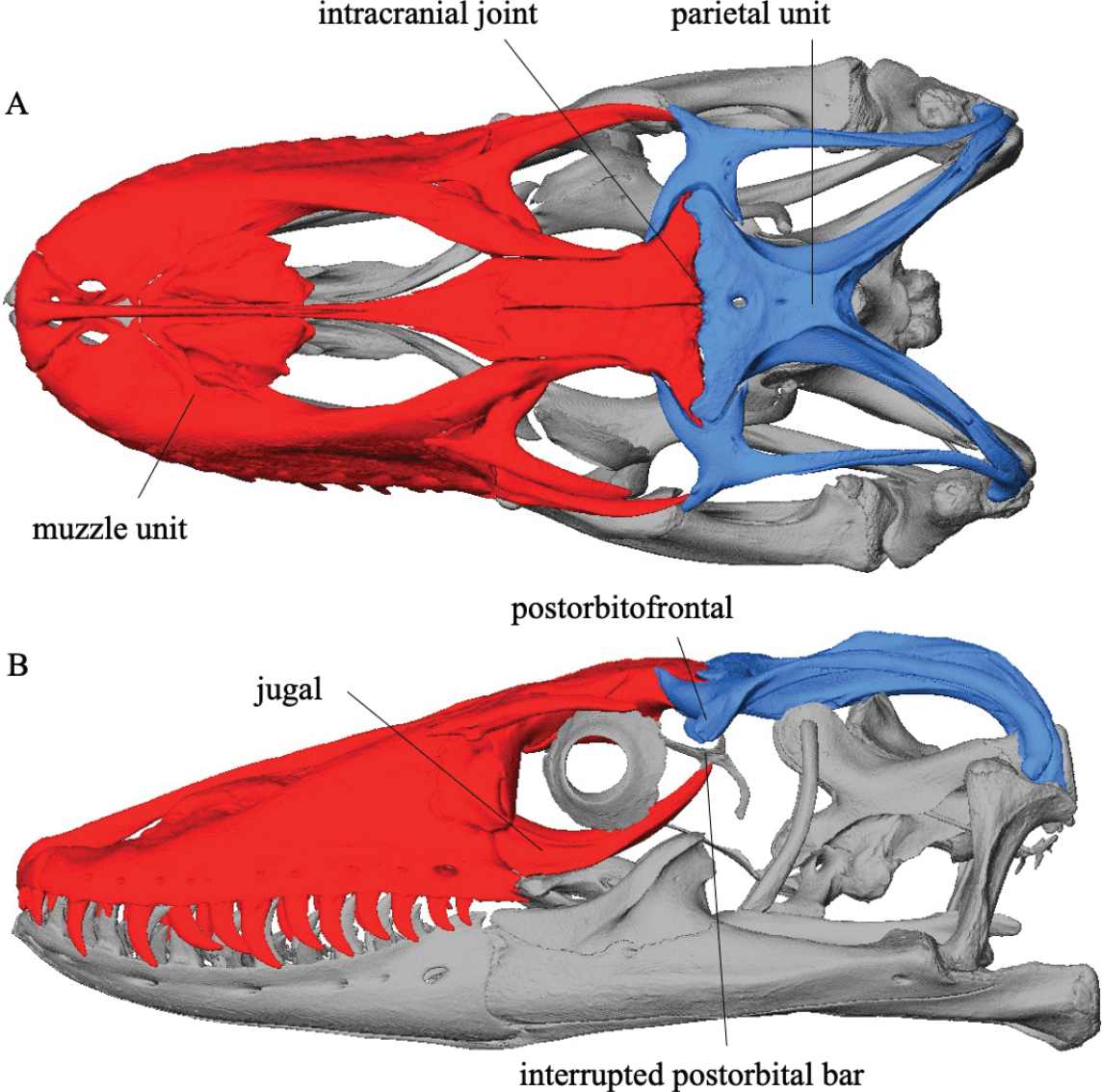
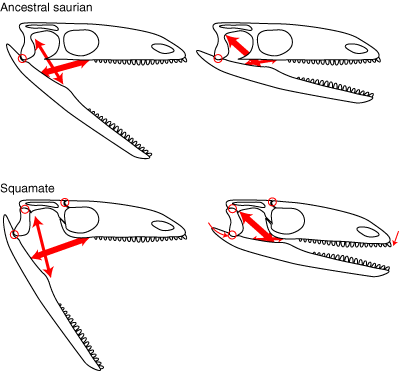
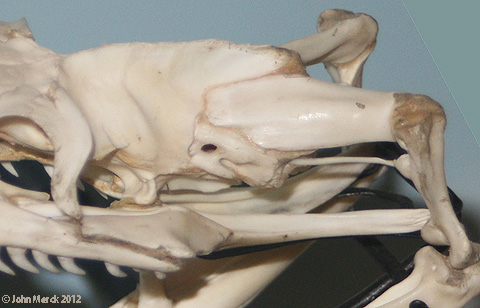
- Hinged quadrate articulation to the skull roof, allowing the quadrate to rock back and forth. Compare Sphenodon, a rhynchocephalian ( above left) with Tupinambis (right from The Reptile Database), the tegu lizard. Associated synapomorphies:
- Overlap between the quadrate and pterygoid is minimal, allowing the ventral end of the quadrate to swing freely.
- Squamosal lacks a ventral process
- Quadratojugal is lost
- Jugal extends anteriorly in orbit margin
- Epipterygoid is reduced to a narrow column
- Angular is short (not reaching the jaw articulation posteriorly)
- Coronoid is complex and forms a tall coronoid process.
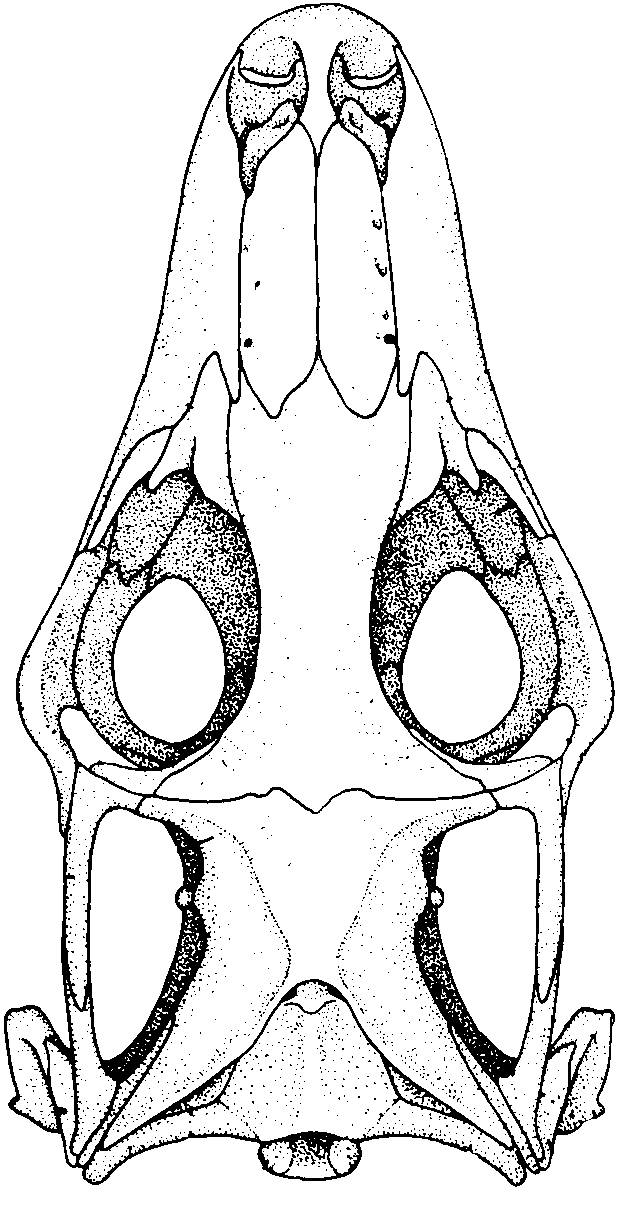
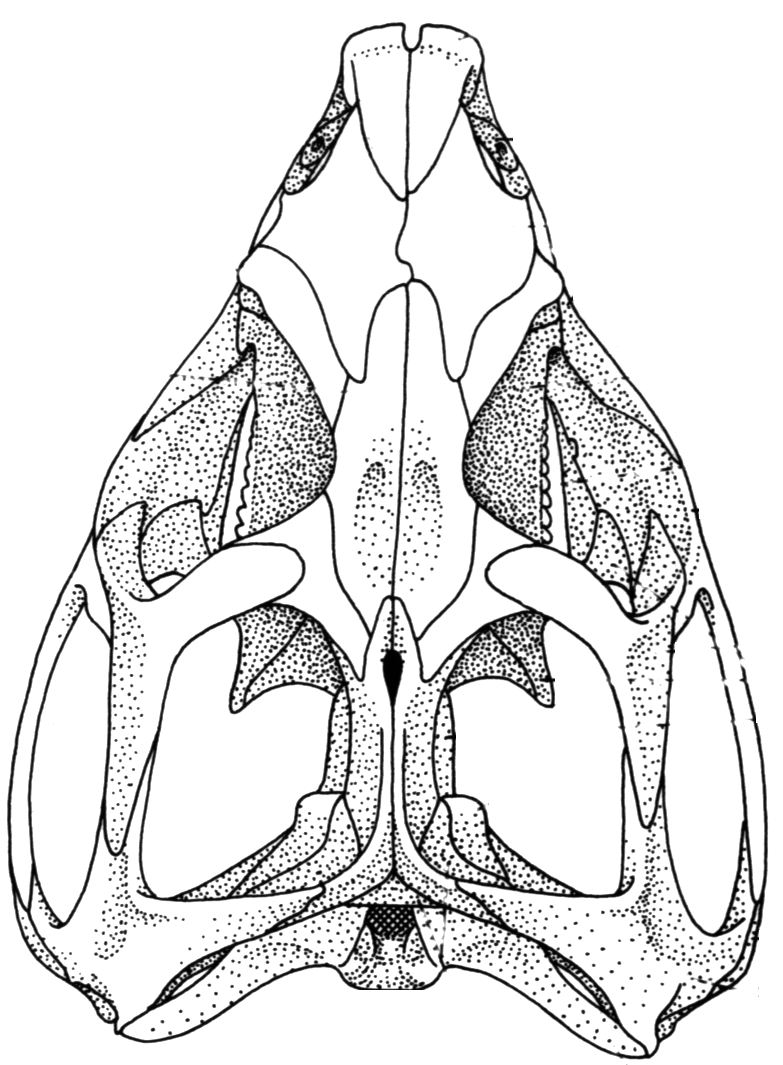
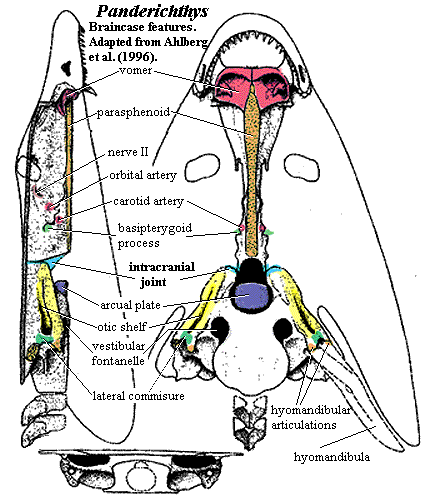
Sphenodon punctatus a rhynchocephalian compared to Tupinambis teguixin, a squamate. - Frontals and parietals meet in a straight line that forms a hinge. See illustration from Kubota et al., 2024. This is functionally similar to the intracranial hinge of basal sarcopterygians. Together with the mobile quadrate, squamate skulls enjoyed a degree of cranial kinesis that enabled them to optimize the leverage of the temporalis and petrygoideus muscles of the jaw.
- Pineal foramen moves to the fronto-parietal contact (but is often absent)
- Nasals reduced
- Premaxillae fused
- Frontals fused
- Parietals fused
- Parietals anteroposteriorly short, such that the occipital region is exposed in dorsal view.
- Opisthotics and exoccipitals fused
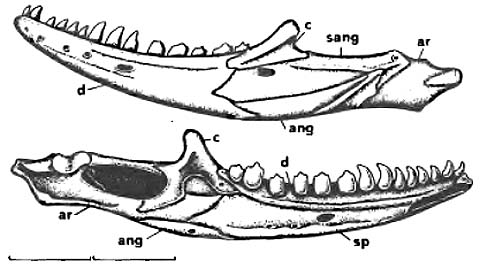
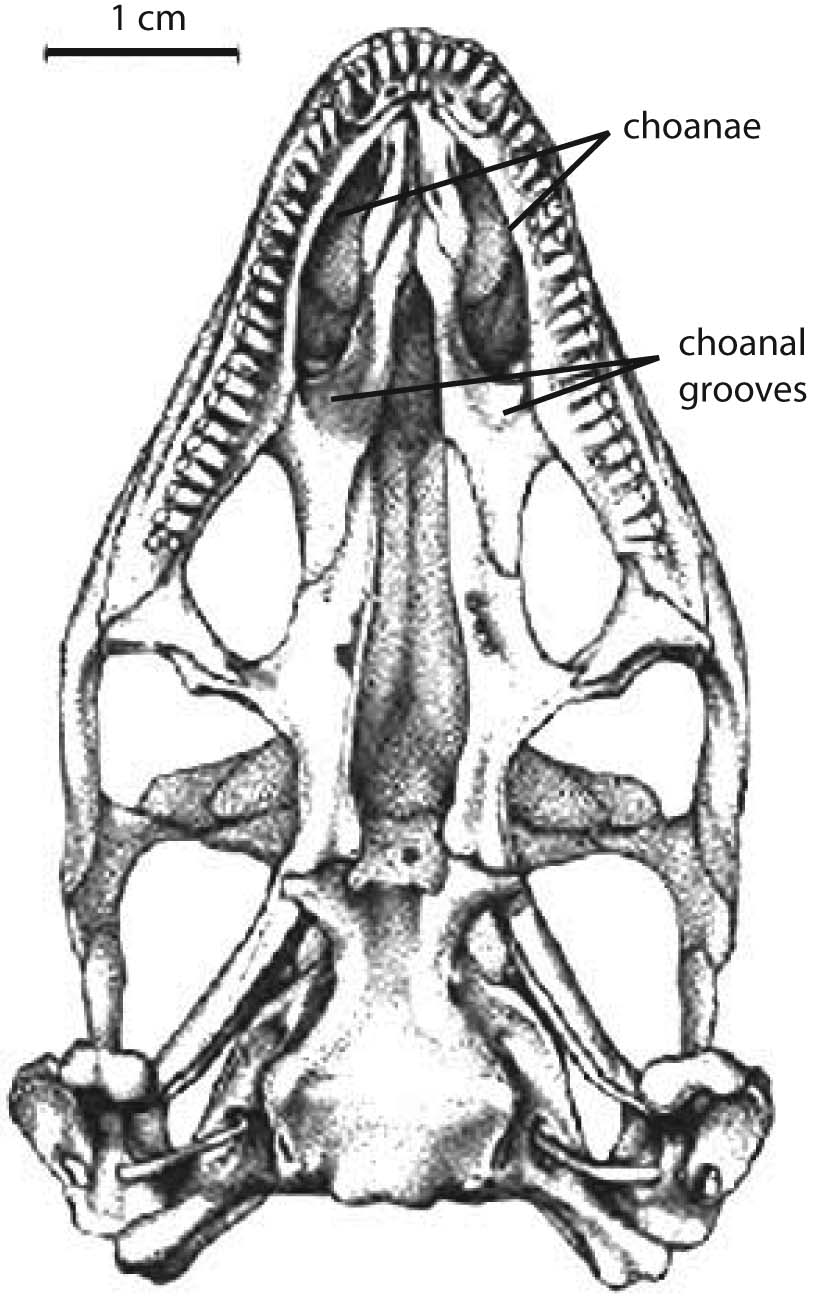
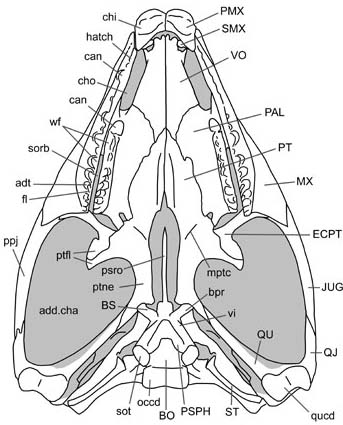
- Palatal teeth reduced
- Choanae recessed in choanal grooves
- Suborbital fenestrae expanded such that pterygoids form part of their medial border
- Enlarged interpterygoid vacuity, such that pterygoids barely touch, if at all
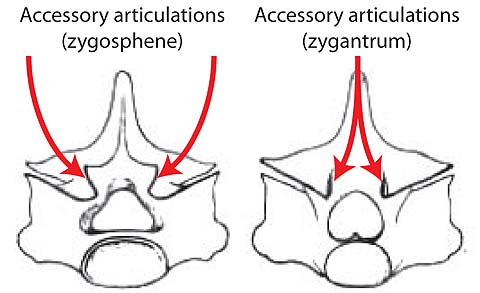
- Vertebral articulations procoelous (concave anteriorly)
- Accessory articulations of neural arches present
- Dorsal intercentra absent
- Coracoid emargination present
- Ossified sternum present, facilitating rotation of coracoids.
- Thyroid fenestra in pelvis
- Astragalus and calcaneum fused to form astragalocalcaneum.
- Distal tarsals 1 and 2 are lost.
This is a bewildering list, however some major adaptations shine though:
- In contrast to rhynchocephalians, whose strong jaws enable them to bite pieces off of their prey, the anatomy of the kinetic squamate skull is adapted to seizing and holding prey, then swallowing it whole. In some, this is developed into an astonishing ability to wrap their heads around their prey.
- Squamate locomotion is improved, with increased rotation of forelimbs and robust consolidation of ankle bones. Link to video.
Diversity:
This huge list of synapomorphies suggests a very long squamate stem, however we have only one definite well-known stem squamate:
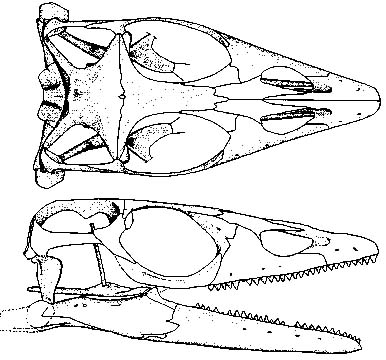
- Huehuecuetzpalli mixtecus, from the Early Cretaceous. Displays most of the synapomorphies listed above, except that:
- Premaxillae remain paired
- Vertebrae are amphicoelous
- Distal tarsal 2 is still present
Living Crown Squamata
Diversity: Since the early days of cladistics, a short list of major monophyletic groups appears has typically appeared:
- Iguania: (Cretaceous - Quaternary) iguanas, anoles, fence lizards, horned lizards,
agamids; chameleons, etc.

Phelsuma quadriocellata - Gekkota: (Late Jurassic - Quaternary) Paedomorphic forms including Geckonidae and Pygopodidae.

Varanus panoptes - Anguiformes: (Early Cretaceous - Quaternary) Anguidae, Helodermatidae, Varanoidea.

Tiliqua scincoides - Scincidae: (Jurassic - Quaternary) Skinks, with broad smooth scales and a tendency toward limb reduction.
- Cordylidae: (Cretaceous - Quaternary) Girdled lizards, with heavy dermal armor.
- Lacertidae: (Paleogene - Quaternary) Old World wall lizards.
- Serpentes: (Jurassic - Quaternary) Snakes. "More subtle than any beast."
- Teiidae: (Cretaceous - Quaternary) New World whip-tails, race-runners, tegus.
- Amphisbaenia: (Early Cretaceous - Quaternary) The "worm-lizards." Limbless (with one exception) burrowers with extremely strong skulls modified for burrowing.
- Dibamidae: (Quaternary) Nearly limbless burrowers in soil and leaf-litter.

Crotaphytus collaris






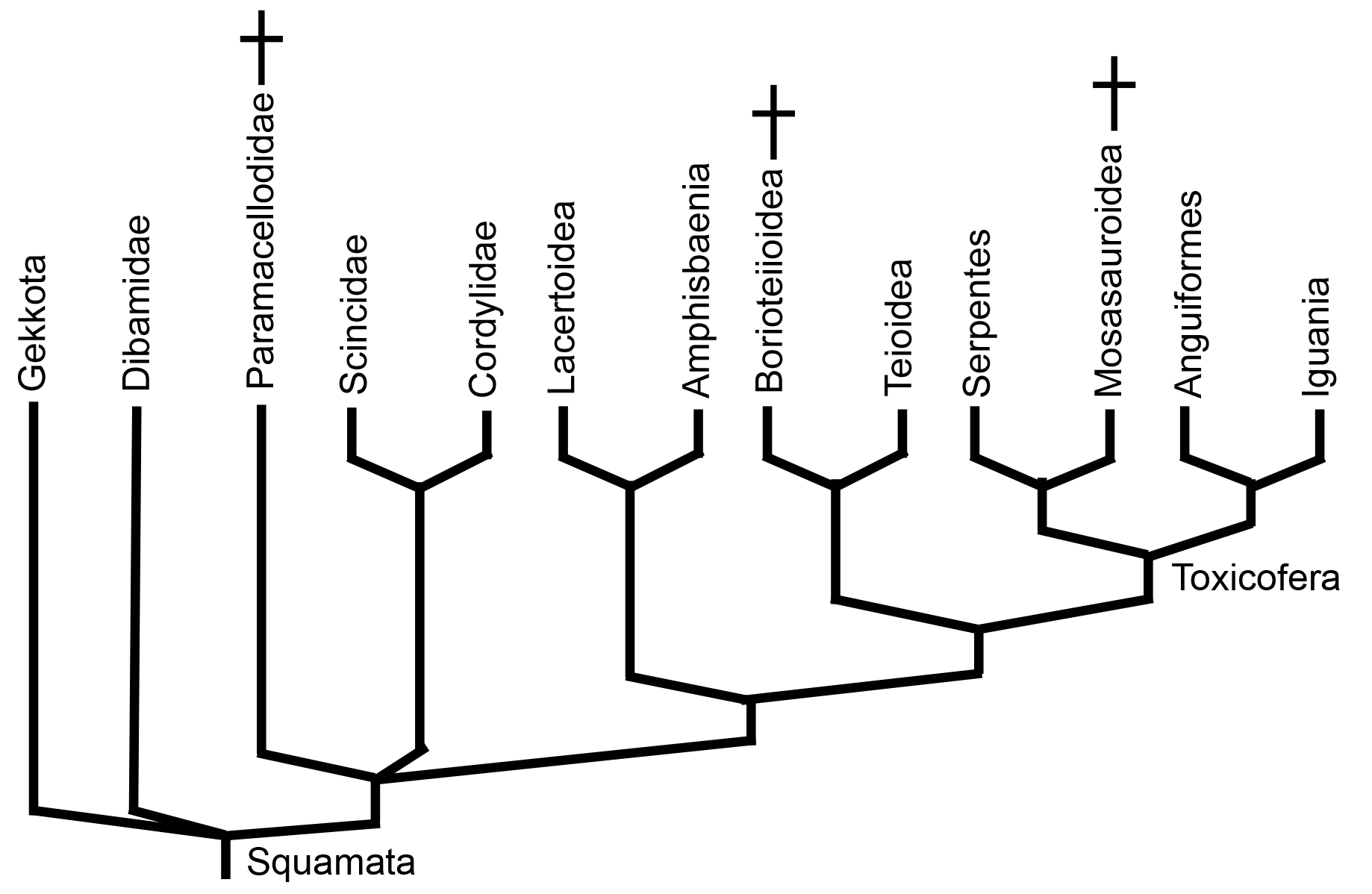
Fossil Crown Squamata
- Paramacellodidae: (Jurassic - Cretaceous) Known from fragmentary remains, but with distinctive multicusped teeth. Usually appear in phylogenetic analyses near Scincidae.

- Borioteiioidae: (Cretaceous - Paleogene) AKA "Polyglyphanodontia." Largeish herbivores with transversely expanded teeth and robust skulls. (Indeed, Polyglyphanodon has reestablished a complete infratemporal bar.

- Mosasauroidea: (Cretaceous) Diverse radiation of semiaquatic - fully marine squamates, including marine apex predators of the Late Cretaceous. (See below.)
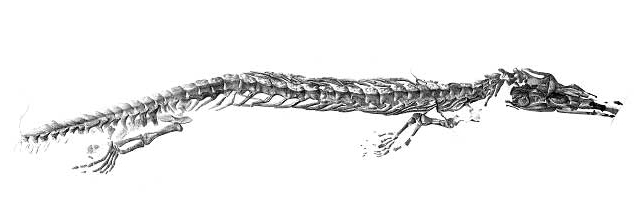

Cryptovaranoides microlanius
- Brownstein et al., 2023. Refute the squamate affinities of Cryptovaranoides, insisting that it is an archosauromorph.
- But this is rebutted by Whiteside et al., 2024, who assert that Cryptovaranoides is absolutely a squamate.
- an archosauromorph masquerading as a squamate
- a crown squamate with a time-machine or...
- an incorrectly dated rock unit.
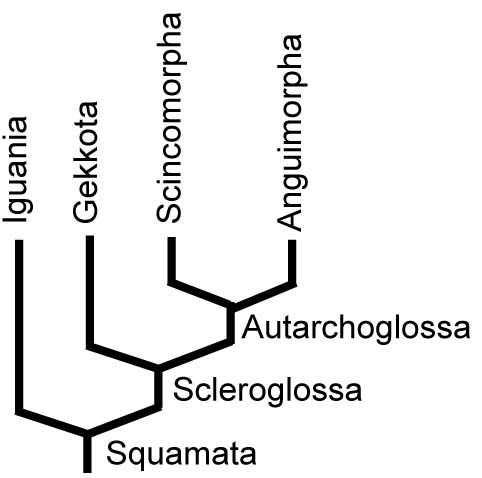
Squamate Phlogeny 1986 - the naive version:
Morphology-based analyses have, since the early days of cladistics, recovered the pattern at right where Squamata is divided into two large groups:- Iguania: whose members:
- use the tongue in prey capture
- Emphasize limb-propelled locomotion with large limbs (plesiomorphy?)
- Scleroglossa: characterized by:
- Scaly tongues
- Loss of the lepidosaurian modified mid-dorsal scale row.
- Autarchoglossa, specialize the anterior tongue as a sensory structure - the classic forked tongue.
- Scincomorpha: Scincidae, Cordylidae, and Lacertidae.
- Anguimorpha: Anguiformes and Serpentes.
Scleroglossans show a tendencies toward:
- the reduction of the limbs and contain several limbless groups
- the use of venom (or caustic saliva) in prey capture. (Parallel to a small handful of iguanians.)
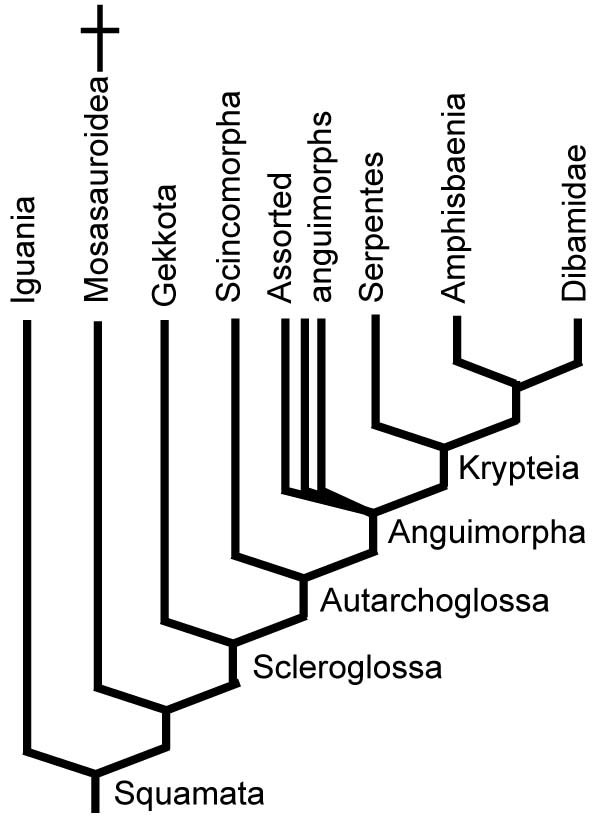
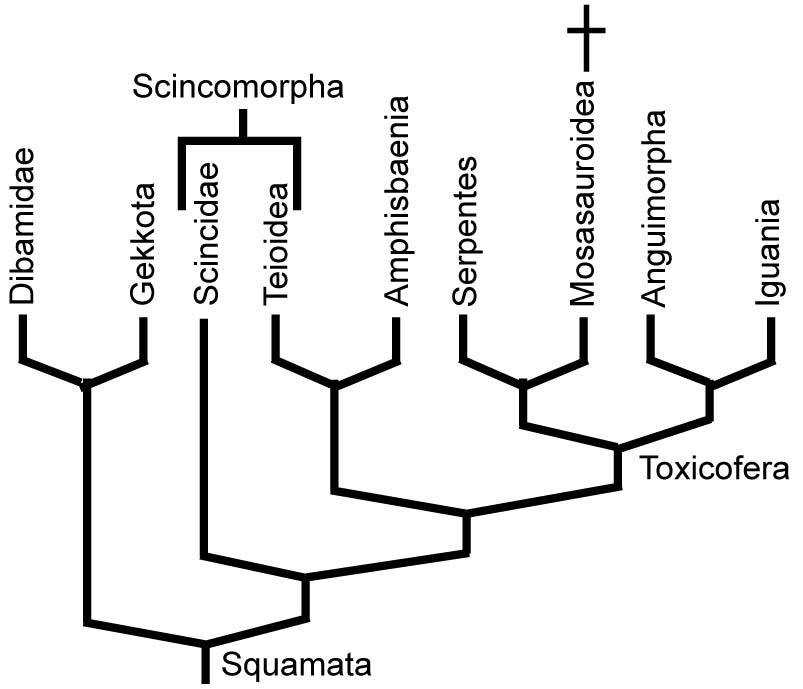
But forget it! This consensus has evaporated with the inclusion of greater taxon samples and molecular data, with the first fifteen years of the 21st century being a time of competing hypotheses and the last ten years seeing a slow march toward a new consensus. The biggest points of contention have been:
- The most basal squamate group: The 1986 version put Iguania in this position, but more recent analyses place Iguania near the squamate crown. Gekkota and Dibamidae vie for the "most basal branch" prize.
- The position of mosasauroids
- The identity of the sister taxon of snakes
Squamate superlatives I: Mosasauroidea: (Cretaceous)

Plotosaurus bennisoni
A look at the quadrate of Tylosaurus (right) clearly shows the apomorphic squamate quadrate. Other interesting mosasaurian features include:
- Elongation of the snout
- Retraction of the nares
- Development of a distinct hinge joint at the mid-length of the jaw and corresponding weakening of the jaw symphysis.
- Organization of the palatal teeth into a single stout row on the pterygoid.
Mosasaur ecology: Despite their long history of study, mosasaurs continue to surprise us.
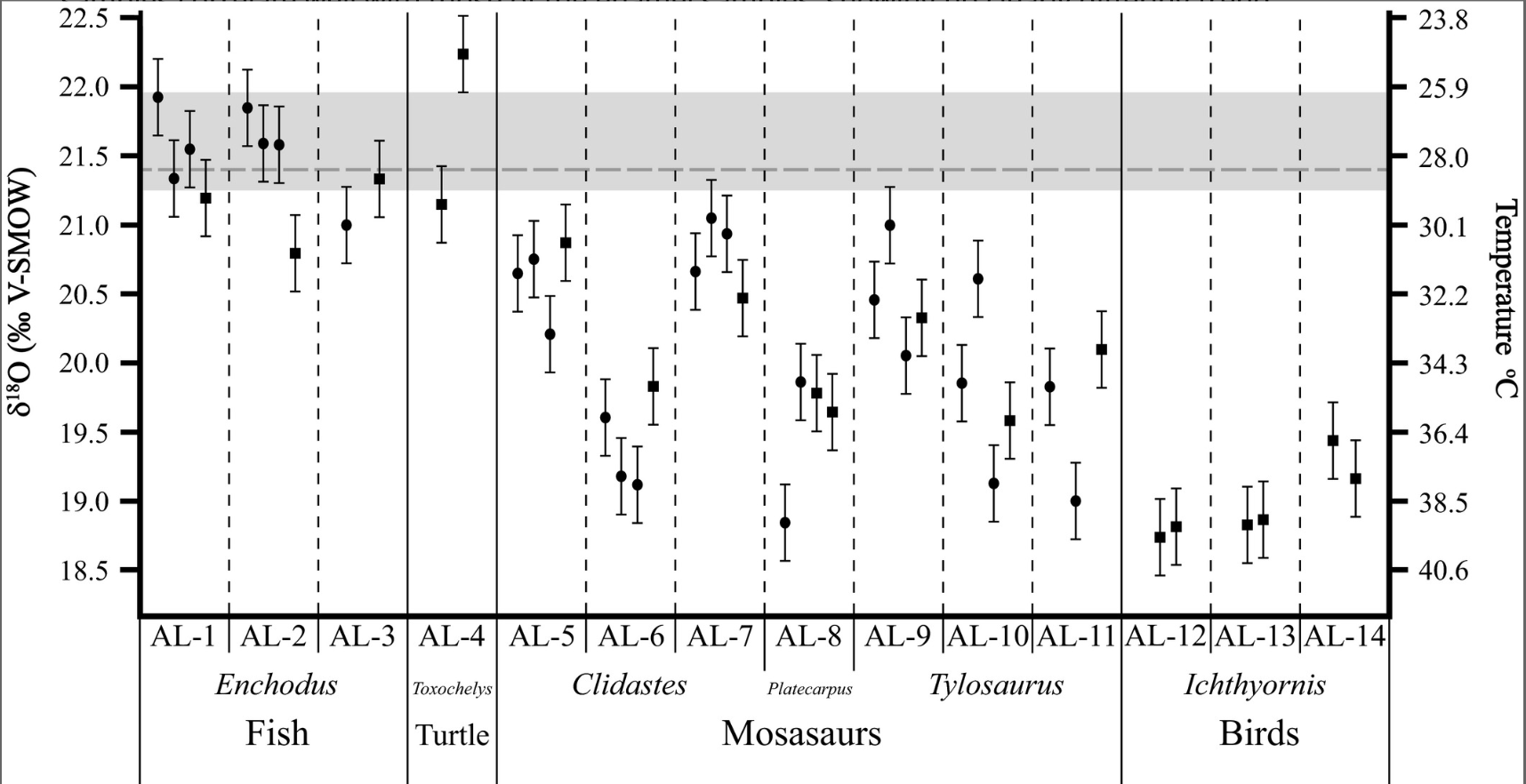
- Endothermy: Although more common in low-mid latitudes, mosasaurs are found at most paleolatitudes raising the question of whether they could have been endothermic. Harrell et al. 2016 addressed the question by comparing δ18O values (a proxy for temperature) in the bones and enamel of fish, turtles, toothed-birds, and mosasaurs from two locations/rock-units in Alabama controlling for paleoenvironment and diagenesis. Results indicate that, in contrast to the actinopterygian Enchodus, mosasaurs displayed a range of elevated temperatures, in some case approaching those of sea birds. Results varied among individuals and taxa, however.
- Reproduction: Until recently, mosasaurs' mode of reproduction was mysterious. Did they come onto the beach to lay eggs or give live birth at sea? According to Legendre et al. 2020, neither. They describe a 29 cm. long leathery egg-shell from Antarctica that could only be from a mosasaur. It was apparently laid at sea then immediately hatched - in effect, a live birth with a vestigial egg-shell.
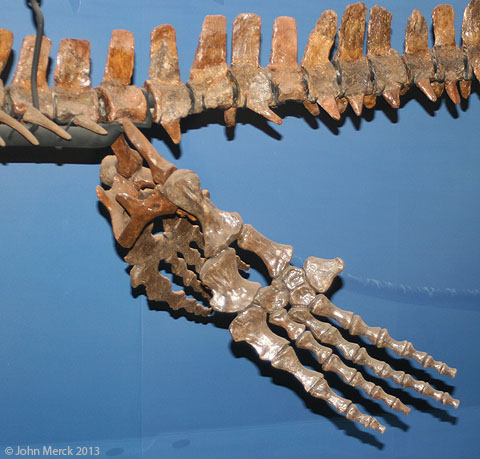
- But could they swim? Well, yes, but the exact roster and timing of transformations associated with swimming remains incomplete. Mosasaurids - the big aquatic forms - are derived from basal mosasauroids that resemble living squamates with some aptitude for swimming, including the water monitor Varanus Salvator. Derived forms, however, possess apomorphies including:
- Fore and hindlimb transition from ancestral ("plesiopedal") to derived aquatic form ("hydropedal") through shortening of the stylopodium and zeugopodium and elongation of the autopodium. In very derived forms, this is accompanied by hyperphalangy - the increase in the number of phalanges.
- The pelvis loses its attachment to sacral ribs, becoming "hydropelvic."
- The mosasaur tail has always been reconstructed as laterally flattened but similar to other lizard tails. Lundgren et al. 2013 demonstrated that the mosasaur Prognathodon, at least, had a full-blown tail fin. Could other mosasaurs have been that different?
- Squamation: When preserved, mosasaur scales are fine, rhomboidal, keeled, and less than 5 mm in length.

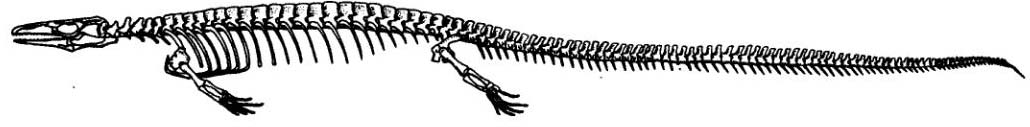
Opetiosaurus buccichi, an "aigialosaur"
Relationships: All agree that Mosasauroidea insists of:
- Mosasauridae - proper marine mosasaurs
- Dolichosauridae - elongate forms with especially long necks
- "Aigialosaurs," a paraphyletic group of semi-aquatic medium-sized squamates. While aigialosaurs might have spent time in the water, they were not strongly specialized for marine life.

We can't get into the weeds on the systematics of Mosasauridae - a speciose and diverse group, but one question depends our attention that has been prominent in recent literature: How many times did mosasaurs become fully aquatic? This question stems from the presence of taxa like Dallassaurus (right) that seem to nest within the tree of derived aquatic mosasaurs but which display plesiopedal and plesiopelvic anatomy. Does their presence indicate that fully aquatic life was acquired more than once or that some mosasaurs reverted to semi-terrestrial living?
- Bell and Polcyn, 2005 describing Dallassaurus noted that basal members of otherwise marine clades Mosasaurinae and Halisauromorpha contain basal members - either Dallasaurus or taxa previously regarded as "aigialosaurus" whose presence suggested that the full marine habits was independently acquired.
- Simões et al., 2017 employed a more sophisticated range of phylogeny reconstruction methods to develop an alternate hypothesis where creatures like Dallasaurus represent reversals, not the plesiomorphic condition.

Squamate superlatives II: Serpentes
(Late Jurassic - Rec.) The most subtle of beasts.
Rather than giving a synapomorphy list, we will note major evolutionary trends:
- Limbs reduced or absent.
- Lidless eyes protected by a transparent scale.
- A unique pattern of rods and cones in the retina resulting in a reduction of color vision (typically well-developed in sauropsids.)
- External ear opening lost.
- The temporal region is long in comparison to the snout (a condition found in other "head hunters" like aistopods.)
- Braincase reinforced anteriorly by laterosphenoid ossification.
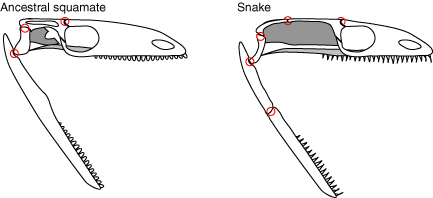
- Bones of the dermal skull roof and braincase fuse into a solid cylindrical unit.
- The quadrates, palate bones, maxillae and premaxillae become very loose and mobile, allowing manipulation and swallowing of large food items.
- The supratemporal, the skull bone to which the quadrate attaches, also becomes mobile with respect to the braincase.
- The teeth of the palate are organized into a pair of rows.
- The jaws are jointed at their mid-length and flexible, and in most cases, the two sides do not connect in front, allowing swallowing of even larger objects.
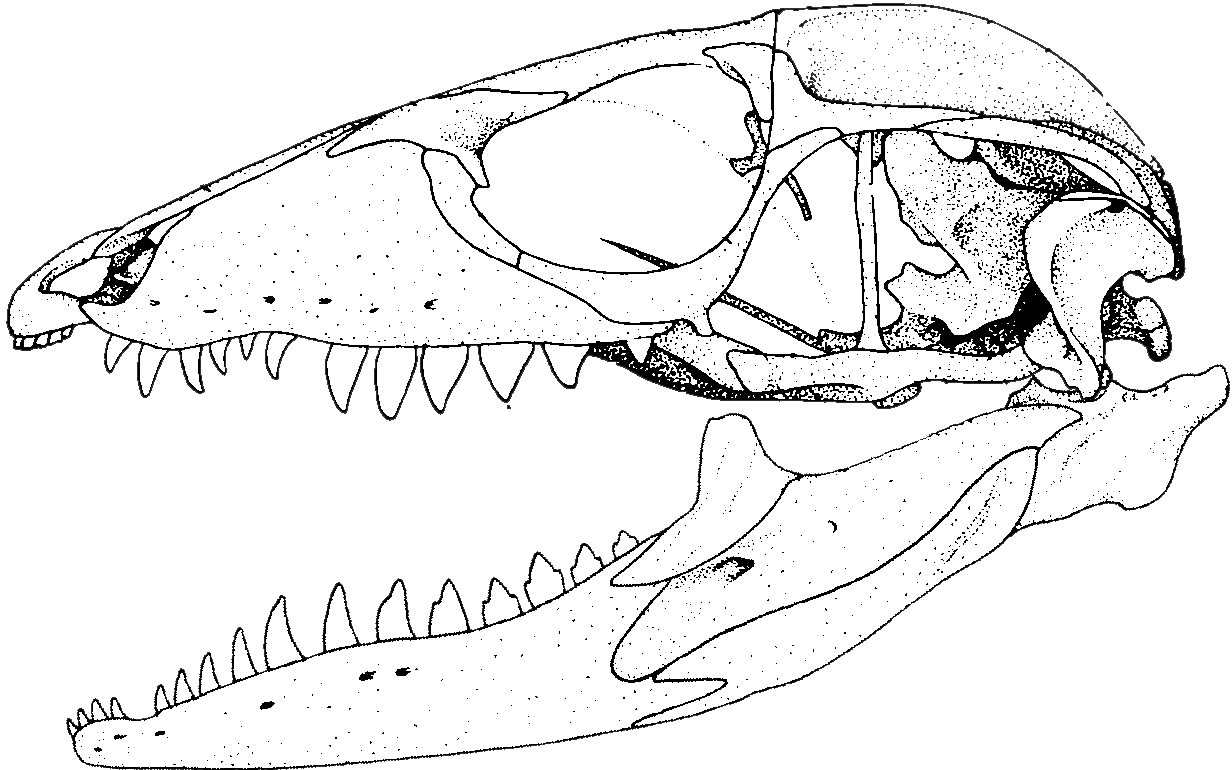
The comparison of the skulls of the Nile monitor lizard and reticulated python (a snake showing a relatively ancestral serpentine morphotype) underscore these differences.
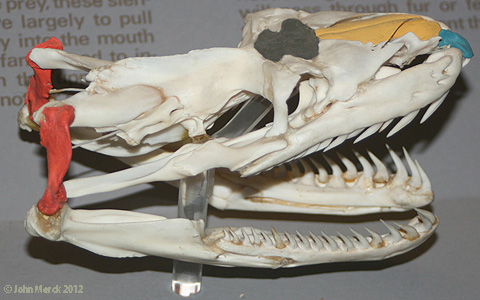
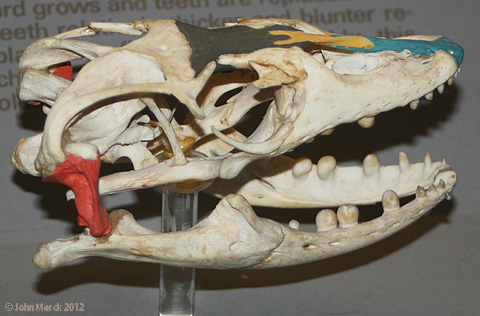
Nile monitor (left) and reticulated python (right) with premaxillae (blue), nasals (yellow), frontals (brown) and quadrates (red) highlighted.
These trends are even more pronounced in derived snakes like vipers.
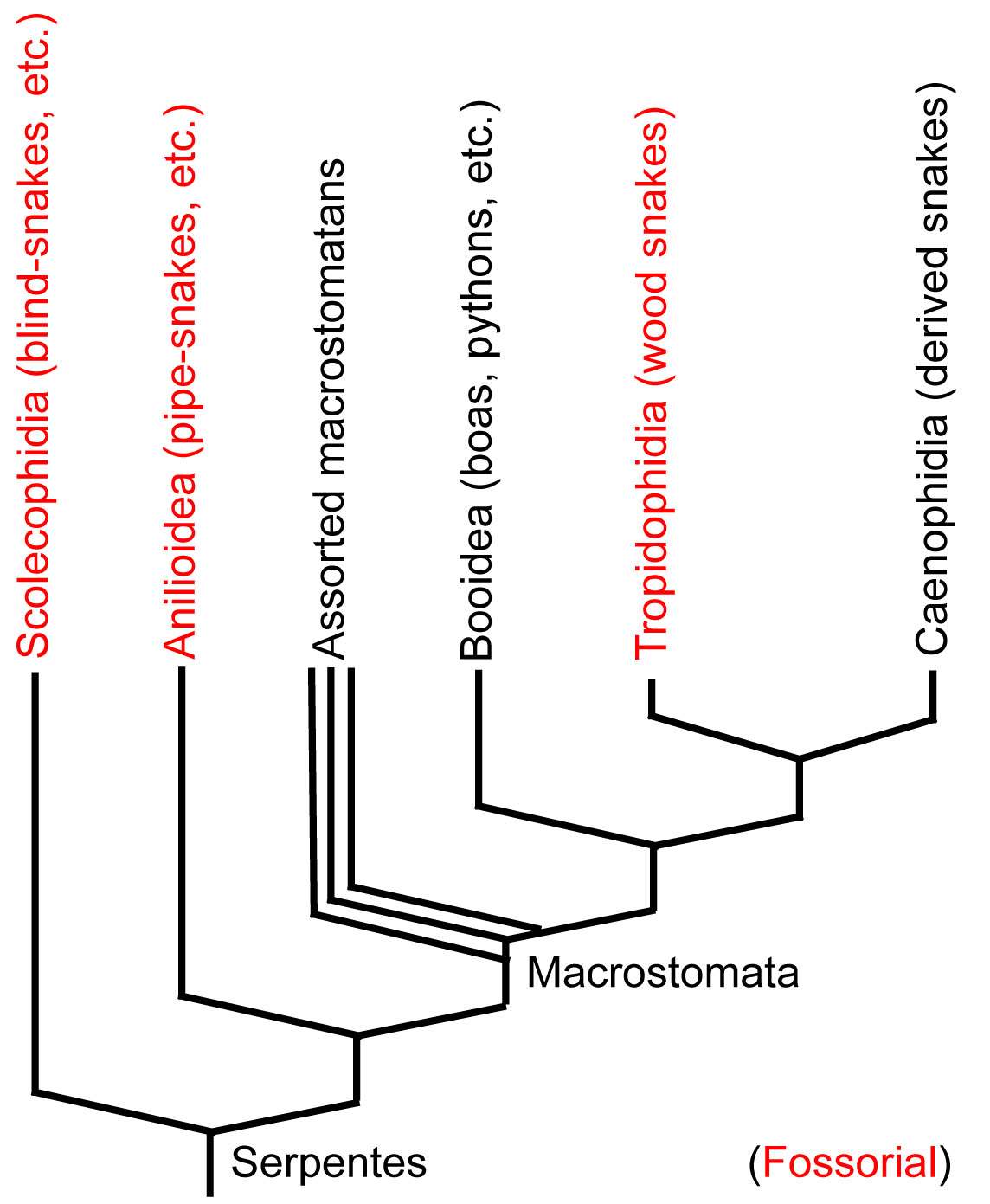
What kind of animal was the first snake?
Two hypotheses have been proposed, both of which address snakes odd sensory systems:- The aquatic hypothesis: The ancestral snake's visual system was modified for the lower lighting underwater. Loss of the tympanum reflects lack of need for impedance-matching ear underwater.
- The fossorial (burrowing) hypothesis: The ancestral snake's visual system was reduced in an ancestor that mostly lived underground and had little to see. Reduction of tympanum reflects advantage, underground, of picking up vibrations with jaw.
Basal snakes: Proponents of the fossorial hypothesis derive comfort from the cladogram of living snakes.
- The basal branches of the snake tree, including Leptotyphlops, the blind snake and Cylindrophis, the pipe-snake are definitely fossorial.
- Although their skulls definitely display the derived features of snakes, they definitely lack the extreme modification of more derived snakes. (See Cylindrophis)
- These snakes lack the large flexible jaws and jaw-articulations characteristic of the "big-mouthed" macrostomatan snakes.
Systematists are drawn into this debate because the discovery that the sister taxon of Serpentes was either aquatic or fossorial would weigh heavily in larger disputes.
Hypotheses of Snake Relationships
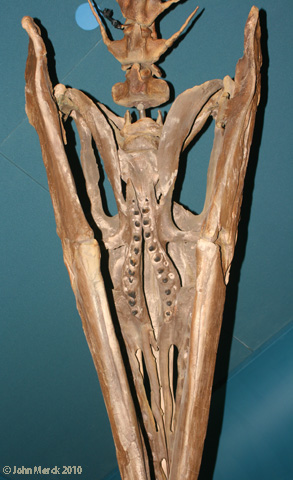

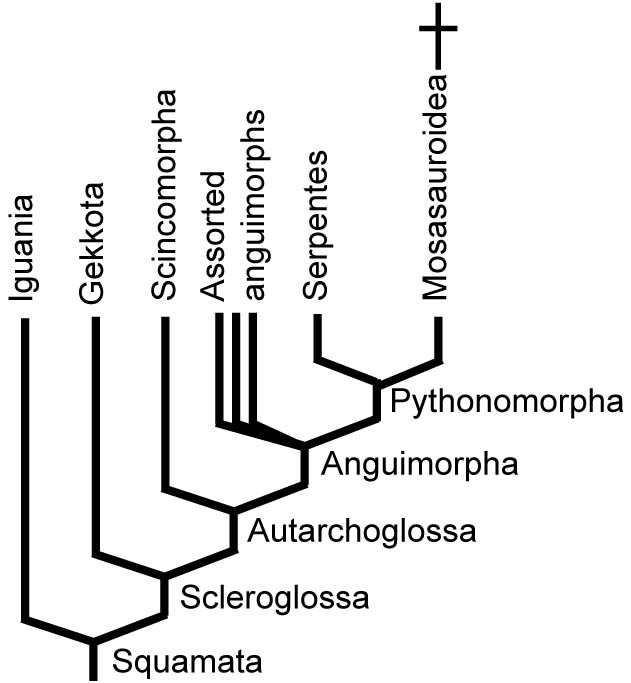
- A single row of stout palatal teeth
- Mobile joint at the mid-length of the jaw.
- Reduction of limbs and limb girdles.

Pachyrachus problematicus
- they were definitely marine
- they retained small but functional hindlimbs.
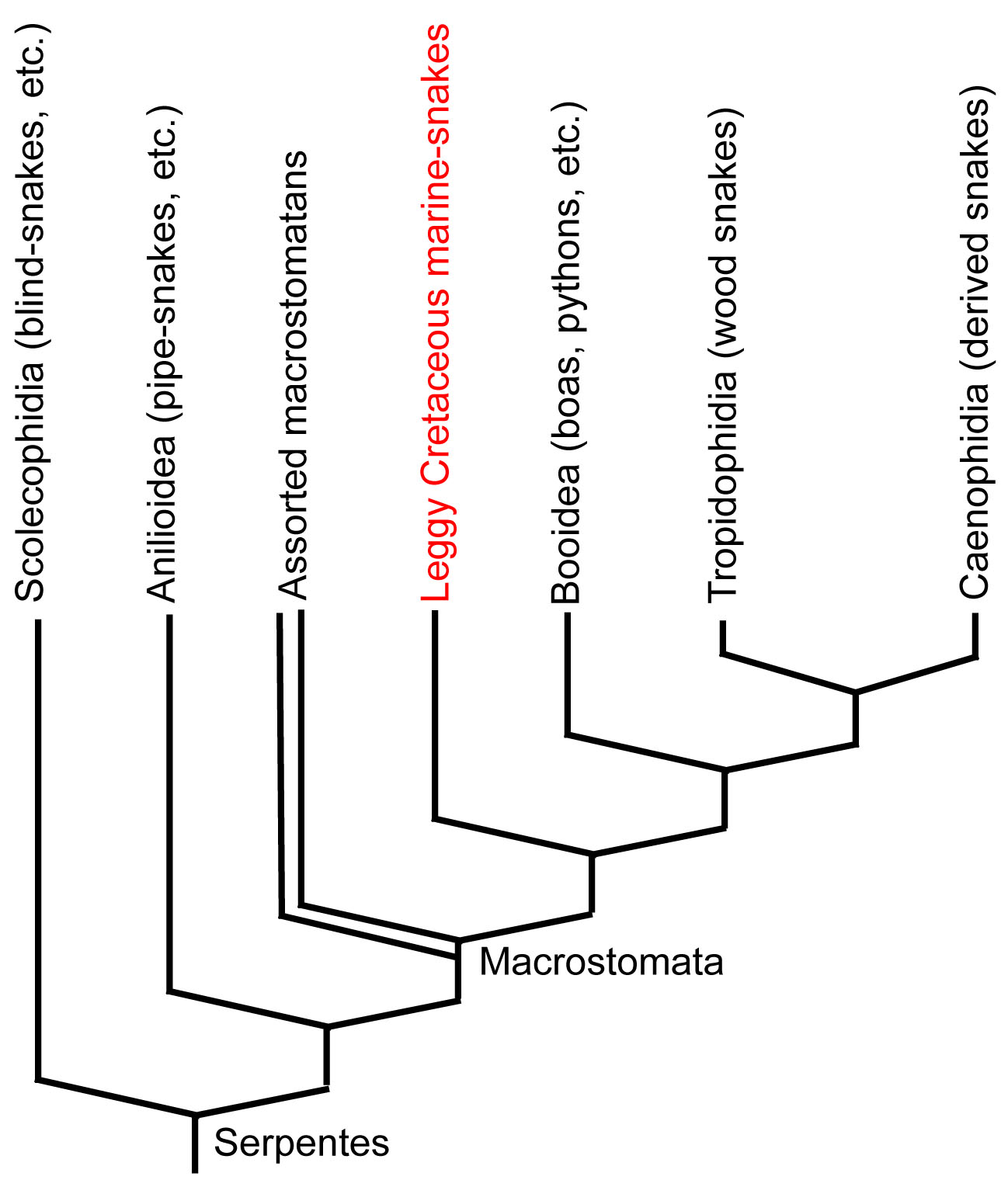 Their skulls proved to have the derived characters of macrostomatan snakes. Thus, despite the presence of legs, most cladistic analyses continue to place them well within the crown of living snakes. Apparently the presence of legs in them is a reversal. Strange! Of course, if they are derived snakes then they do not represent the ancestral condition.
Their skulls proved to have the derived characters of macrostomatan snakes. Thus, despite the presence of legs, most cladistic analyses continue to place them well within the crown of living snakes. Apparently the presence of legs in them is a reversal. Strange! Of course, if they are derived snakes then they do not represent the ancestral condition.
More recently, Caldwell et al., 2015 have revealed leggy snakes from the Late Jurassic. These, in contrast, to Pachyrhachis, occupy a basal position in Serpentes.
The fact that for over a year, the four-legged Early Cretaceous Tetrapodophis amplectus was regarded as a basal snake (Martill et al., 2015) only later to be identified as a dolichosaur (Caldwell et al., 2016) emphasizes the strong similarities between snakes and mosasauroids.
Thesis, Antithesis, and Synthesis
The debate on the phylogenetic position of snakes has fed larger controversies about the relative importance of morphological and molecular data in phylogenetic analysis.Some landmarks in the unfolding debate:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}