The Archosauromorph Stem
John Merck
Archosauromorpha: (Permian - Quaternary) Max ∇ (Passer (sparrow) ~ Lacerta (a squamate)). The node-based group on which Archosauromorpha is anchored is Archosauria, the last common ancestor of crocodilians and birds and all of its descendants - a crown group. As a total-group, Archosauromorpha has no formal synapomorphies.
Nevertheless, known archosauromorphs display evolutionary trends that make them recognizable.
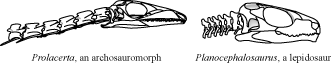
- Overlapping cervical ribs.
- Relatively long rhomboidal cervical vertebrae.
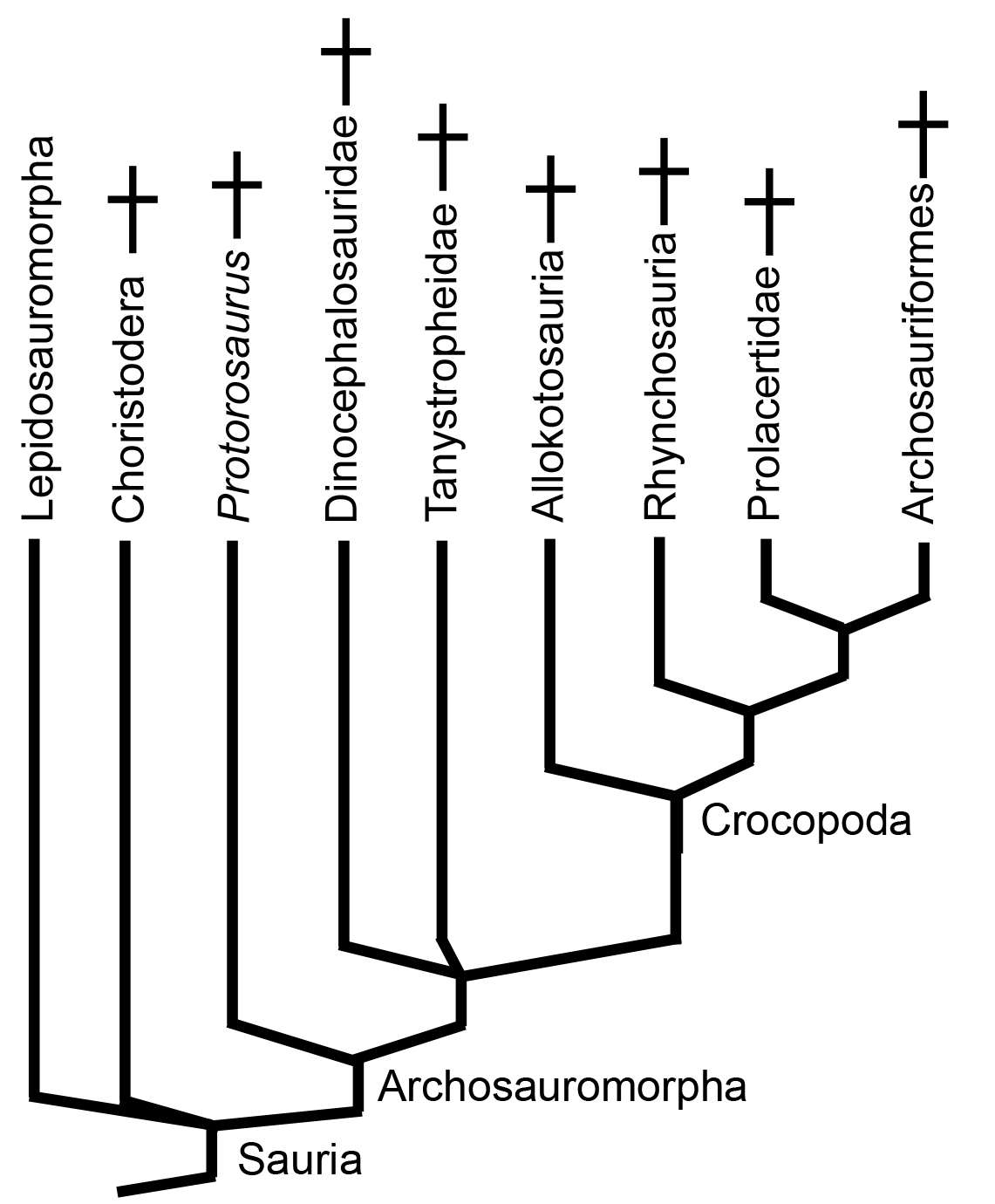
Basal stem archosauromorphs:
Unlike the primitive relatives of lepidosaurs, whose fossil record is sparse, the basal (i.e. non-archosaurian) archosauromorphs present an embarrassment of riches, being common and ecologically diverse during the Triassic, but having their roots in the Permian. They were among the first targets of cladists. (See Gauthier, 1984, Benton, 1985.)
Although there is almost complete agreement on what animals belong on the archosaurian stem, well-studied taxa in the early age of cladistics were typically divergent and ecologically specialized, including:
- Long-necked fishers
- plant chopping herbivores
- plant-grinding herbivores
- aquatic predators
- terrestrial predators

Choristodera:
(Late Triassic - Neogene) Before we start, an intractable phylogenetic headache: Members of Choristodera - a well-diagnosed group - has been found to be:
- basal archosauromorphs
- basal lepidosauromorphs
- stem saurians.
Originally only known to paleontologists as large long-snouted and medium-snouted forms from the Late Cretaceous and Early Paleogene. These were superficially convergent on crocodilians but, when considered in detail, extremely different as well. Eventually, more basal forms were identified going as far back as the late Triassic from fragmentary specimens of nondescript smaller reptiles already housed in museums. Only recently have well-preserved remains of earlier choristoderes become known.
Assorted choristoderes including Simoedosaurus (a) and Champsosaurus (d)
- Posterior expansion of supratemporal fenestrae
- Contact of prefrontals on midline
- Nasals fused
- Nares confluent
- Neomorph (a novel ossification) closes off much of the craniotomy-quadrate passage.
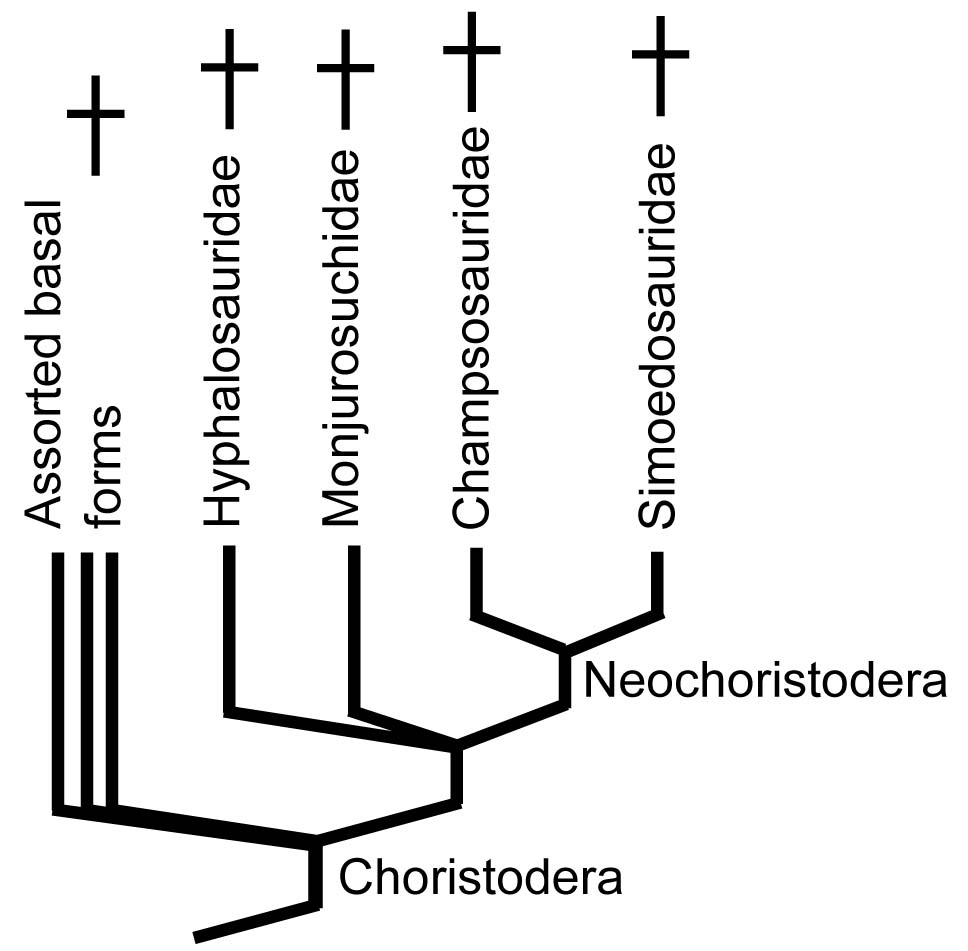
- Monjurosuchidae: (Early Cretaceous) Small terrestrial forms like Monjurosuchus from early Cretaceous.
- Hyphalosauridae: (Early Cretaceous) Small long-necked, small-headed fresh water aquatic forms like Hyphalosaurus from early Cretaceous.
- Simoedosauridae: (Late Cretaceous - Early Paleogene) Croc-sized robust fresh water aquatic forms like Simoedosaurus from early Paleogene.
- Chanmpsosauridae: (Late Cretaceous - Early Paleogene) Croc-sized long-snouted fresh water aquatic forms like Champsosaurus from early Paleogene.

The Base of Archosauromorpha and the Clade that Wasn't:
For the first decade of cladistics, workers like Gauthier, 1984 and Benton, 1985 recovered a clade of longish to long-necked creatures called either Protorosauria or Prolacertiformes that included both terrestrial and somewhat aquatic forms like:- Protorosaurus
- Prolacerta
- Macrocnemus
- Tanystropheus
- Dinocephalosaurus
- etc.
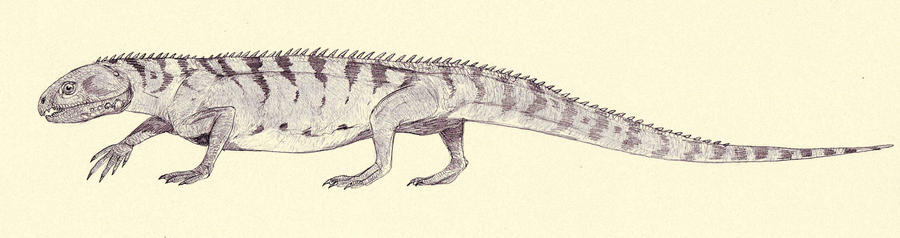
Beginning in the late 1990s, this clade began to fall apart, with forms like Prolacerta jumping toward the crown of the tree (Dilkes, 1998), and others like Protorosaurus (right) falling to the base (Ezcurra, 2016). It now seems that "protorosaurians" represent a relatively basal paraphyletic grade group among archosauromorphs. Indeed, Protorosaurus (Late Permian) is a well-studied close approximation of the ancestral archosauromorph form. It's sister taxon, the long-necked Early Triassic Czatkowiella hare (Borsuk-Bialynicka and Evans, 2009) is similar.
Protorosaurus was fully terrestrial and probably ecologically similar to living varanid lizards. This comparison invites speculation that, like modern varanids, it might have used gular pumping of the hyoid skeleton to ventilate the lungs more efficiently.
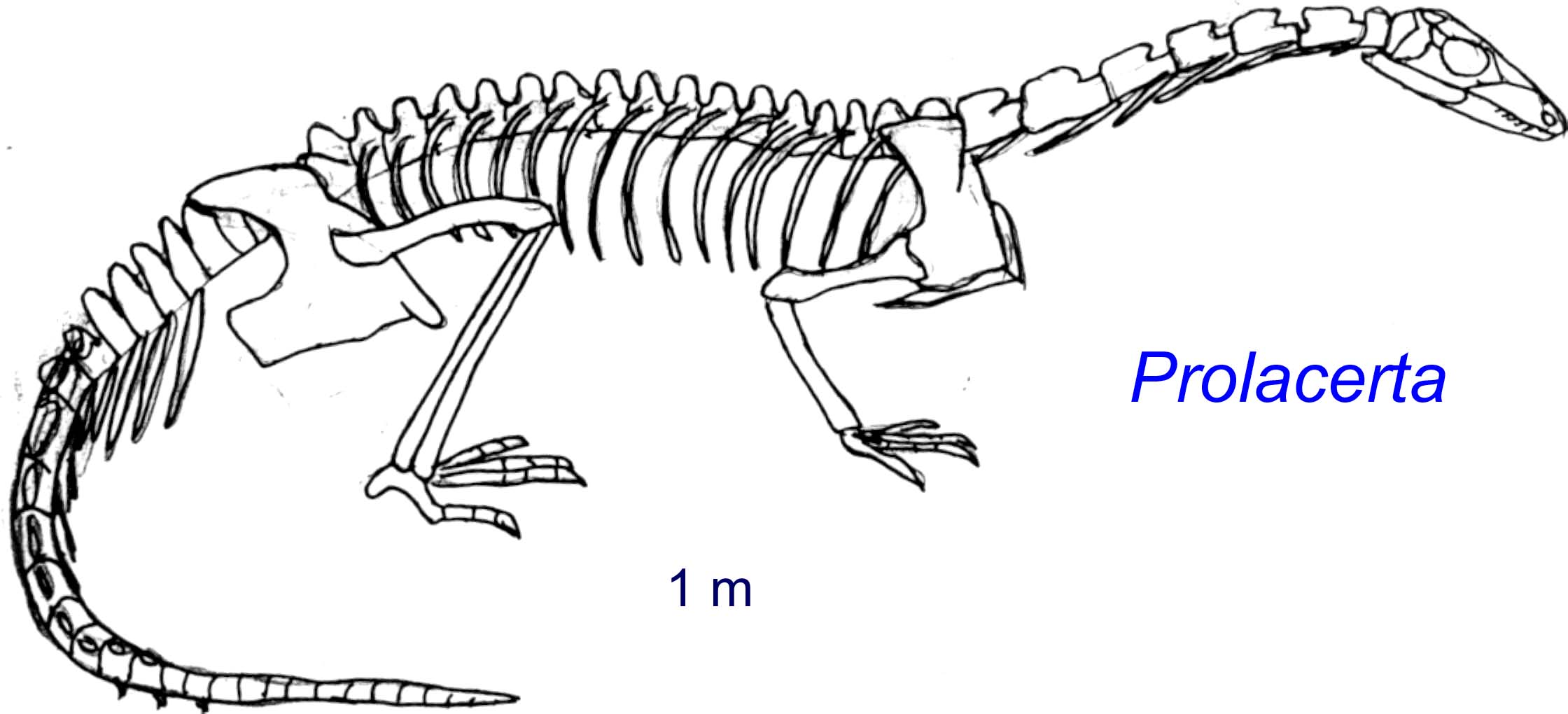
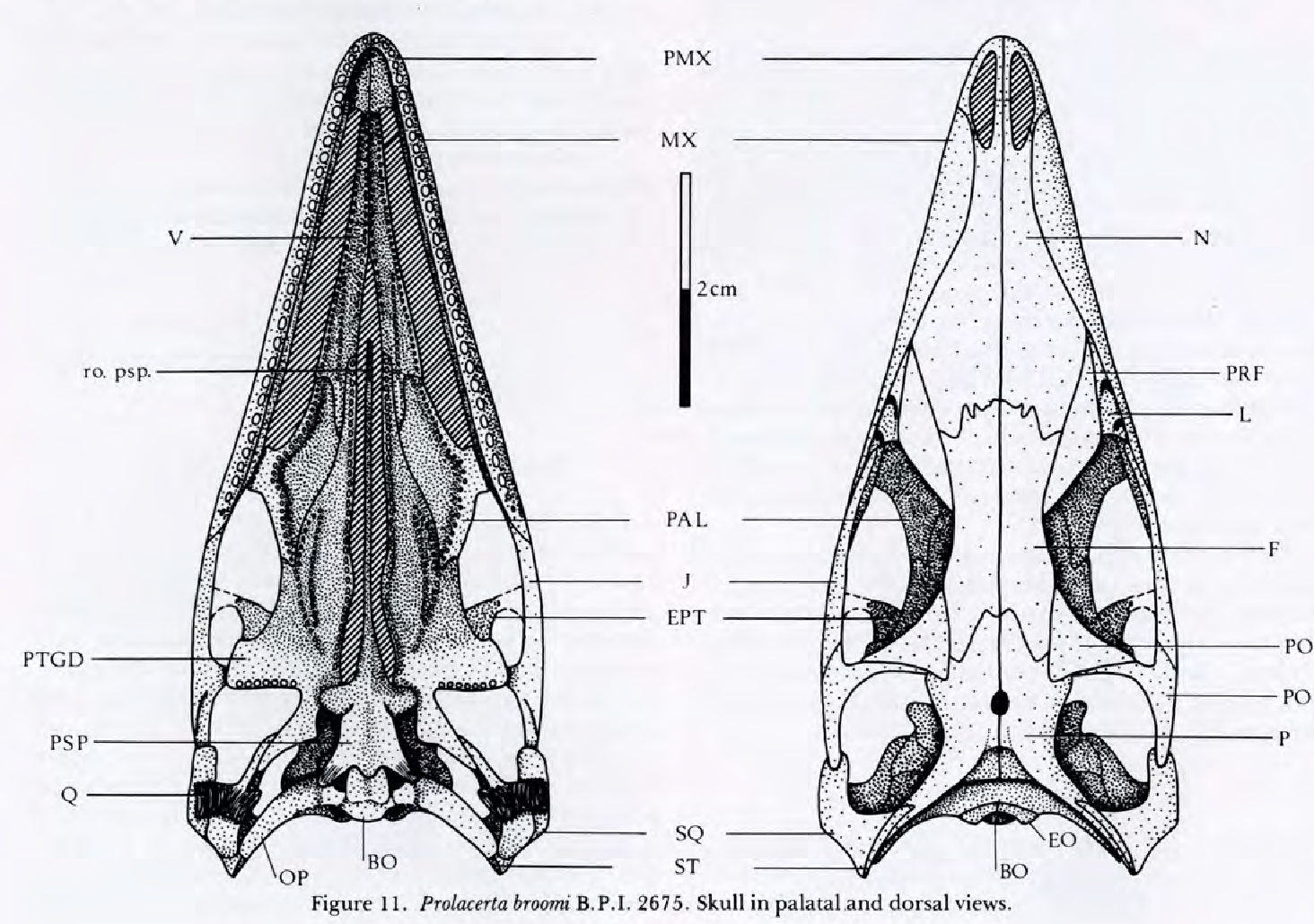
Prolacertidae:
(Early Triassic) An especially well-studied group among "protorosaurians," Prolacertidae has a long history of study. Originally considered a protorosaurian, deemed close to Archosauria by Dilkes, 1998, its current resting place is among other protorosaur-grade creatures (Spiekman et al., 2024). Well known from cranial and postcranial material. Prolacertids, themselves, are diagnosed by:- Parietal-frontal contact that is deeply embayed, with parietals invading the interorbital skull roof.

Dinocephalosauria:
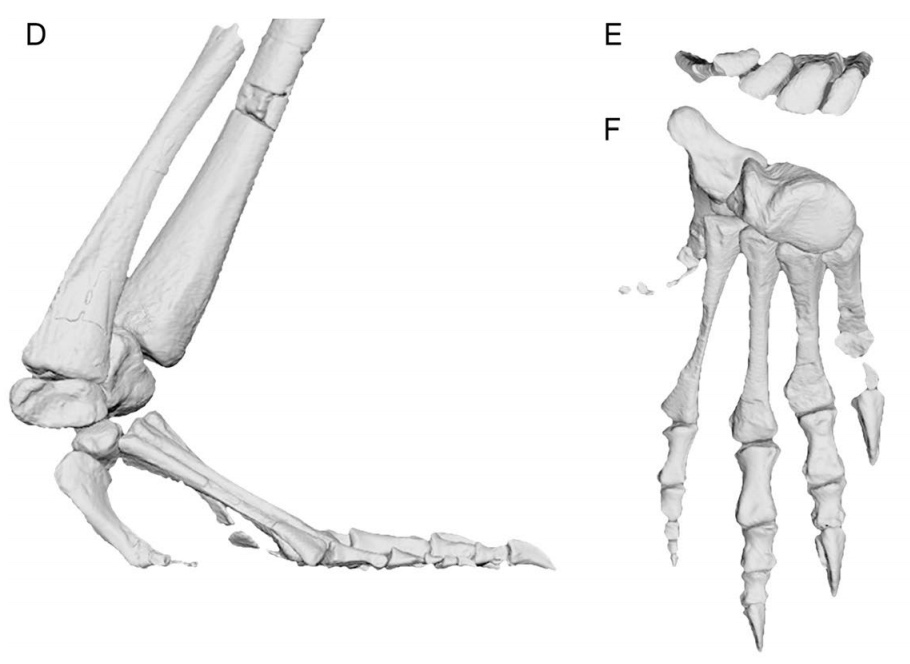
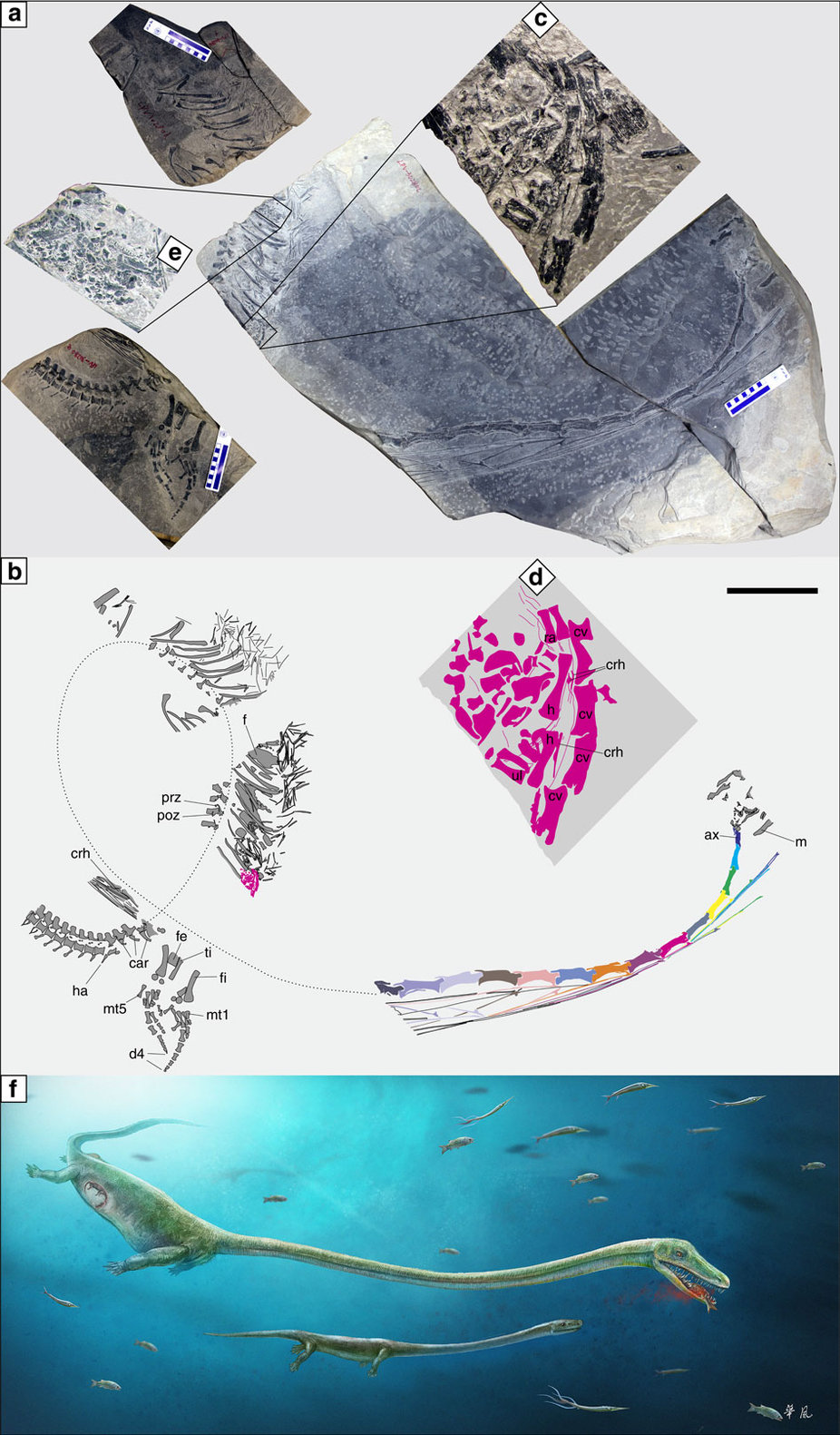
(Middle Triassic) Takes the basic archosauromorph adaptation - a long stiff neck - to extremes through the multiplication of cervical vertebrae, analogous to eosauropterygians. Dinocephalosaurus orientalis, in fact, has 32 cervicals. Dinocephalosaurians resemble other shallow marine long-necked archosauromorphs in general profile but differ in detail. Notably:- The skull is characterized by nares set in long grooves and retracted from the end of the snout.
- The limbs are paddle-like and relatively poorly ossified (in contrast to other archosauromorphs but convergent on eosauropterygians.)
- A specimen preserves a late-term embryo in the abdomen. A radical departure for archosauromorphs, who seem generally to be strongly committed to oviparity.

Tanystropheidae:
(Latest Early Triassic - Late Triassic.) An independent derivation of the long-necked extreme.- Earlier members like Macrocnemus were at most facultatively aquatic, and shows limb proportions similar to cursorial squamates, just with a longer neck. Even so, its remains are found in shallow marine environments, suggesting that it at least lived near water.
- but during the Middle Triassic we pick up fresh-water aquatic forms like Tanytrachelos or shallow marine forms like Tanystropheus. The best known example is Tanystropheus, but others are being discovered Li, et al., 2004. Which (if any) of these swam after prey and which lurked in ambush along the shoreline is debatable.
- Along with their transition to marine life came an elongation of the neck by the evolution of a small number of very long cervical vertebrae.
- The neck was also stiffened by long, slender cervical ribs. And yet, something about large tanystropheids brings out the crazy in illustrators.
- In contrast, the Late Triassic gives us a small radiation of what look like gliding tanystropheids. The best known is Ozimek - small with elongate hindlimb that seem to have supported a gliding membrane. Its relative Sharovipteryx is known from a poorly preserved skeleton but also soft-tissue traces suggesting that the membrane was supported by the hindlimb and tail. This arrangement once provoked some to speculate a connection with pterosaurs, although the preponderance of evidence places them elsewhere in the tree. True enigmas.

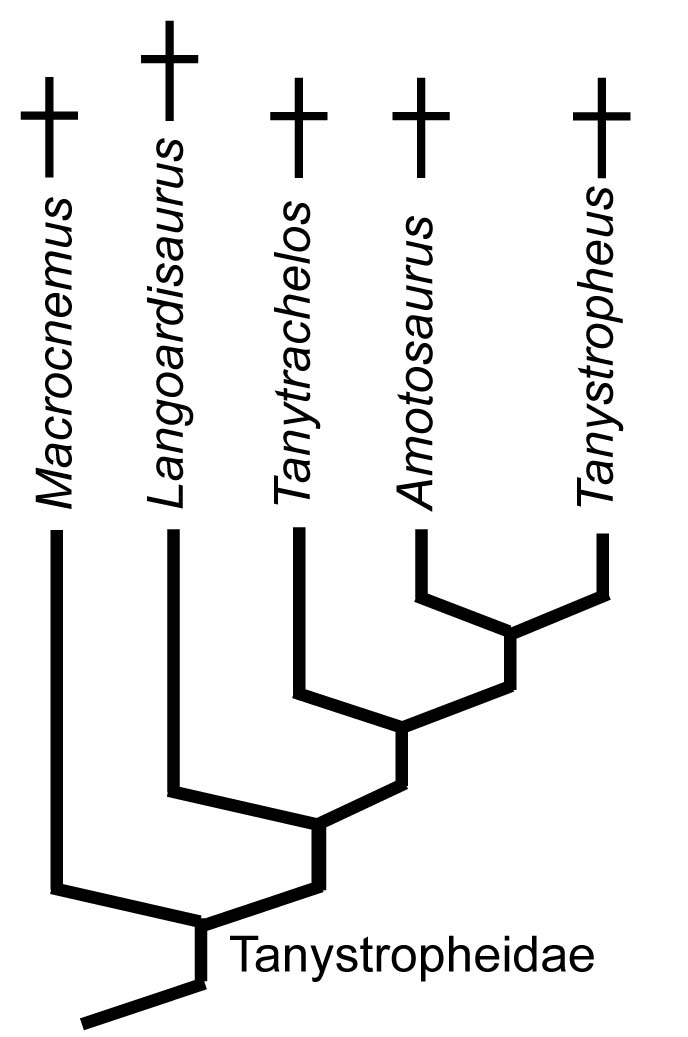
- Elongation of cervical vertebrae
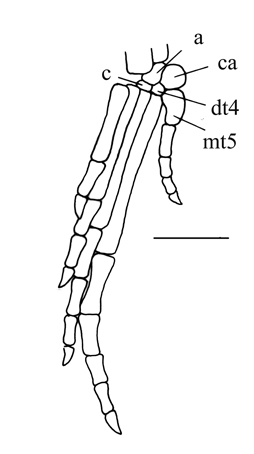
- Shortening of metatarsal V, causing it to resemble a distal tarsal. (In some, the first phalanx of digit V becomes robust and metatarsal-like.)
Synapomorphy of Tanystropheidae + Archosauria:
- Premaxilla with a well-developed postnarial process that excludes the maxilla from the margin of the naris. (Link to skull of Macrocnemus.)
Pros:
- Taphonomy and gut contents indicate they lived in or near marine environments and ate nektonic prey. (Spiekman et al., 2020)
- Their hindlimb were biomechanically poorly suited for striding on land.
- In Tanystropheus hydroides, the nares were retracted posteriorly and positioned on the dorsal surface of the snout.
- The presence of a well-developed embryo in Dinocephalosaurus is strange for an archosauromorph, suggesting some strong selective pressure for ovovivipary. That might be explained by the need to give live birth in the water.
- The long stiff neck presents a real obstacle to agile swimming.
- Their post cranial skeletons lack strong adaptation for swimming.
- They lurked near the shore and ambushed aquatic prey, similarly to a heron.
- Biomechanical analysis of hindlimb suggests they might have swam by simultaneous kicking of hindlimb like a frog, enabling them to lurk on the bottom and ambush prey from below. (Renesto and Saller, 2018)
Crocopoda:
(Later Permian - Quaternary.) Max ∇ (Trilophosaurus ~ Tanystropheus)Named by Ezcurra, 2016, this group encompasses the major terrestrial radiation of archosauromorph herbivores and predators. Significant synapomorphies include:
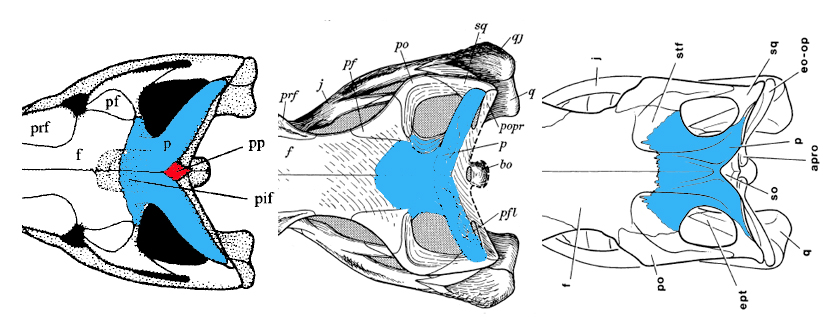
- Changes in the region around the supratemporal fenestra, including:
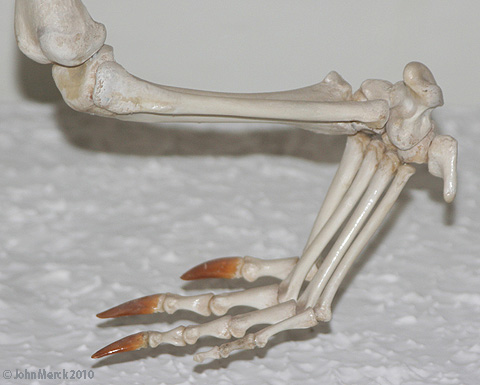
- development of a supratemporal fossa around anterior and lateral margins of fenestra. (See Trilophosaurus - right.)
- transformation of posterolateral parietal into a dorsoventrally deep plate.
- Retroarticular process of jaw upturned.
- Modification of the ankle through the development of a projecting tuber of the calcaneum.
Crocopoda contains:
.jpg)
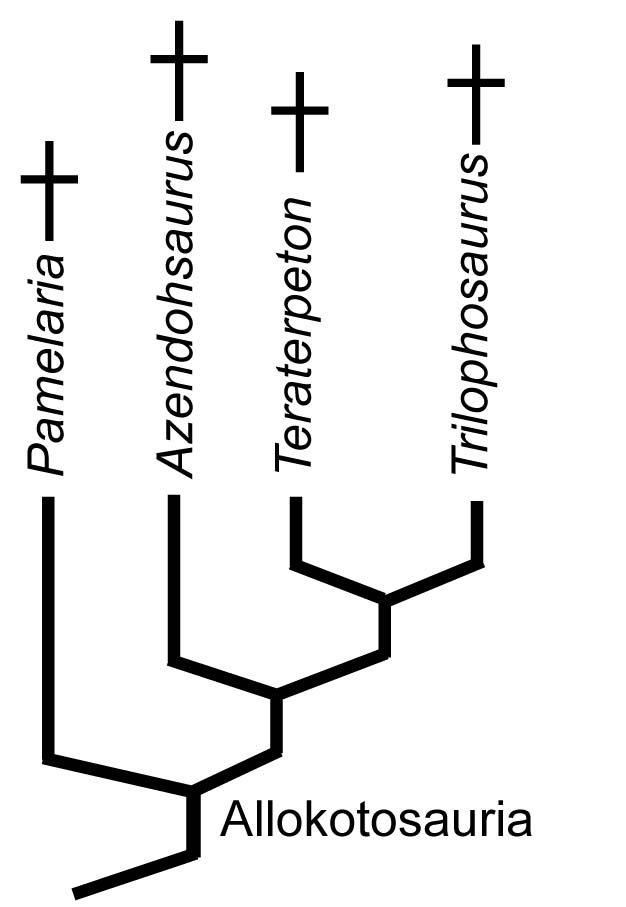
Allokotosauria:
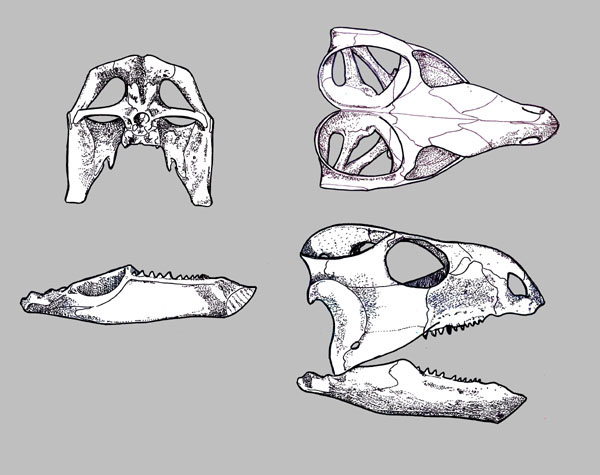
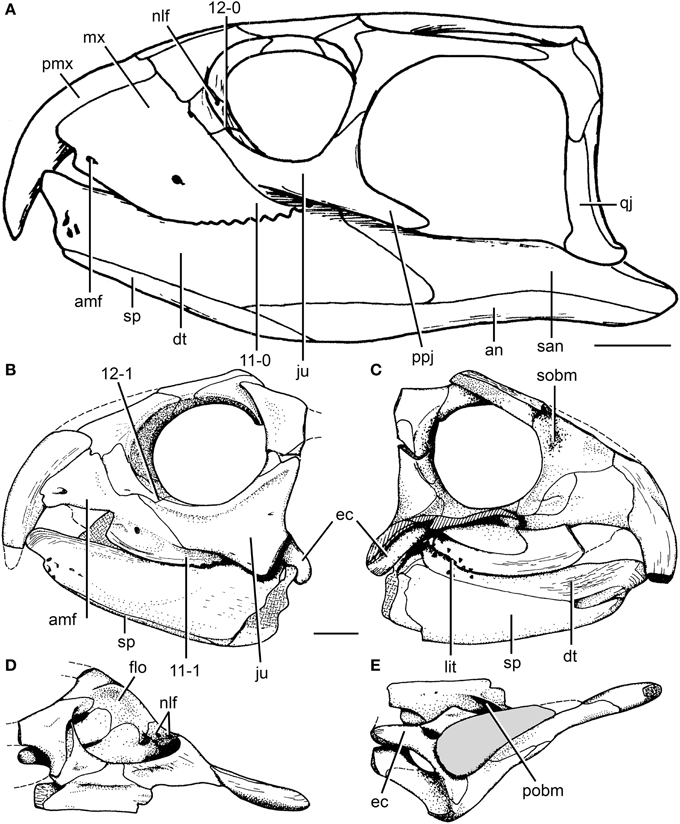
(Middle (Early?) - Late Triassic) Max ∇ (Trilophosaurus, Azendohsaurus ~ other archosauromorphs) (Nesbitt et al., 2015) A herbivorous radiation that was, at first, puzzling because its first studied member was the extremely derived Trilophosaurus buettneri (right) prior to cladistics considered a "euryapsid" because of its temporal fenestration.
Boring on the outside, interesting on the inside. Vaguely resembles a large lizard with a turtle's head, complete with beak in front. Yank its jaws open to admire its wonderfully strange teeth, like a battery of three-cusped parallel razor-blades. Strangest of all, these are maxillary teeth that don't line up with the margin of the skull. Well known from the Late Triassic of North America, but Middle (and possibly Early) Triassic forms are known from Europe (Sues and Schoch, 2023) Freaky. In the last twelve years, related creatures have come to light including:
- The long snouted Teraterpeton Sues, 2003. A long-snouted and slightly less specialized version of Trilophosaurus that retains mediolateraly expanded teeth and a closed infratemporal region.
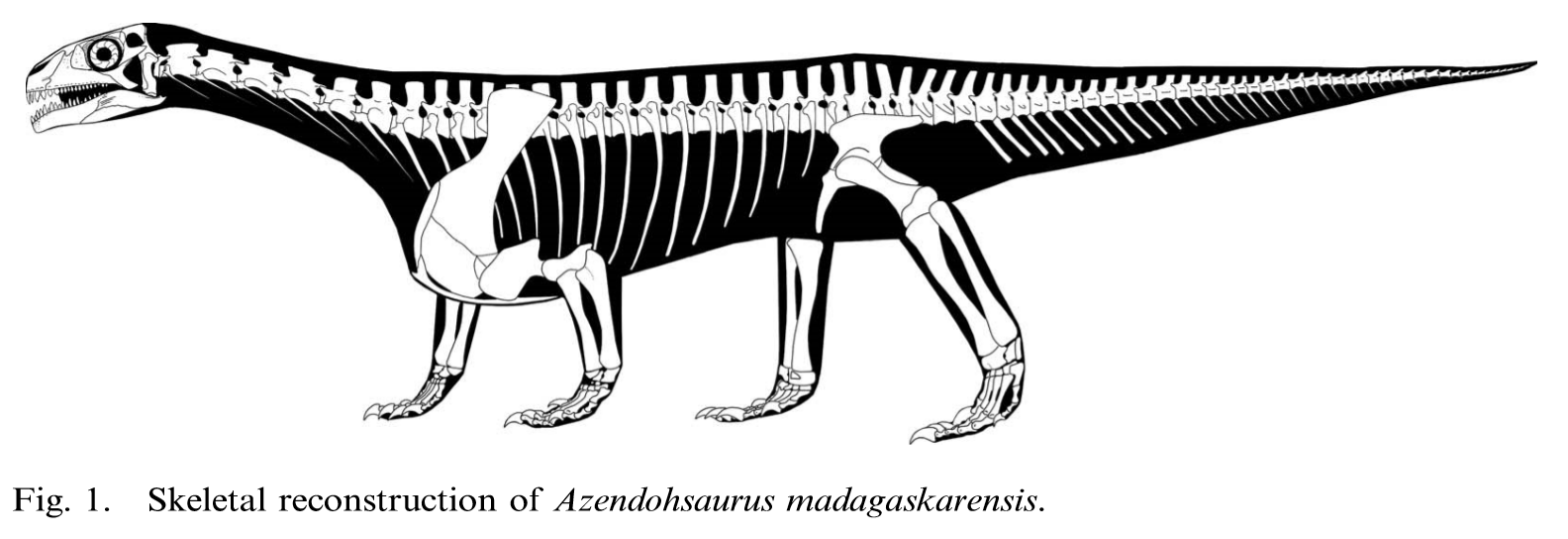
- Azendohsaurus: First described as a sauropodomorph dinosaur, but later revealed to be a stem archosauromorph. Shares adaptations to herbivory with Trilophosaurus (Flynn et al., 2010) although not as extreme, including multi-cusped leaf-shaped teeth and downward deflection of the posterior maxilla.
- Shringasaurus: Cow-sized horned allokotosaur of Middle Triassic India (Sengupta et al., 2017)
- Enlargement of the dorsal process of the jugal
- Development of a rugose lateral border of the prefrontal
- Expansion of the supratemporal fossa onto the posterolateral squamosal

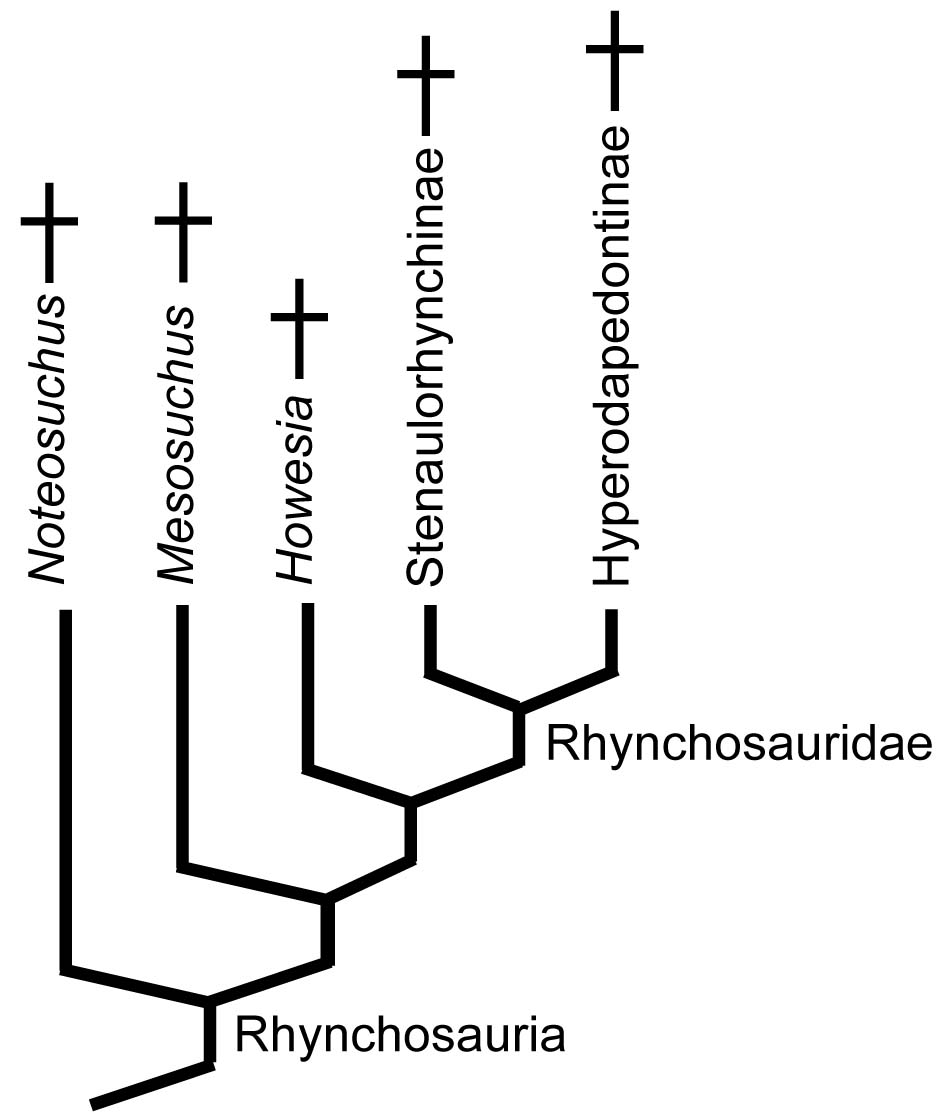
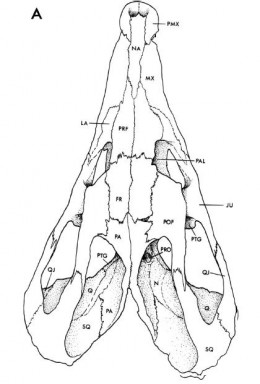
Rhynchosauria:
(Middle - Later Triassic) Min ∇ (Mesosuchus, Howesia) (Dilkes, 1998)Triassic ambulatory scissors for processing tough plant material.
- Ancestral Middle Triassic rhynchosaurs like Mesosuchus seemed normal enough.
- During a brief interval in the Late Triassic, these blossomed into a speciose radiation of the most common plant eating amniotes.
- Proper synapomorphies are very technical, but anatomical trends include:
- Loss of premaxillary and anterior dentary teeth
- Teeth present are permanently fused to bone
- Development of maxillary tooth batteries
- Nares confluent
- Secondary closure of the infratemporal bar
- Reorientation of the iliac blade to project anterodorsally.
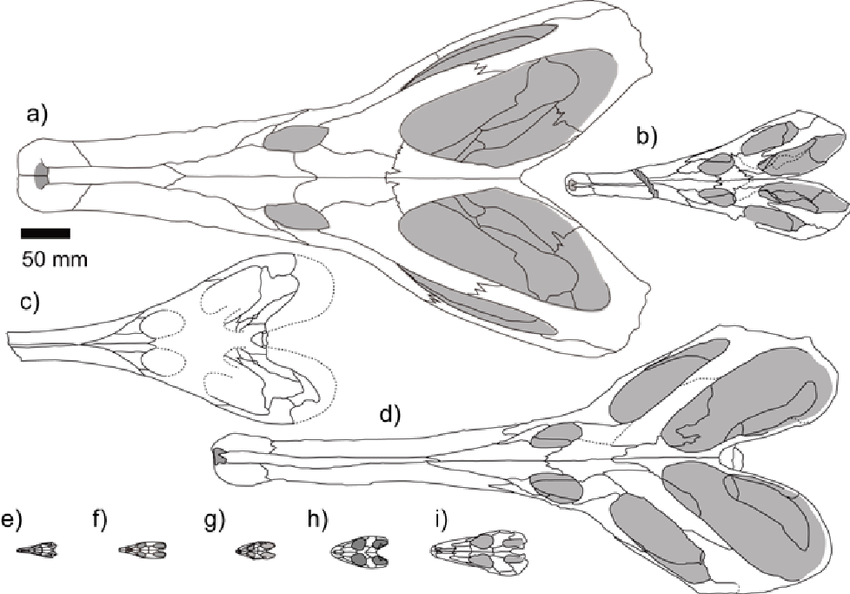
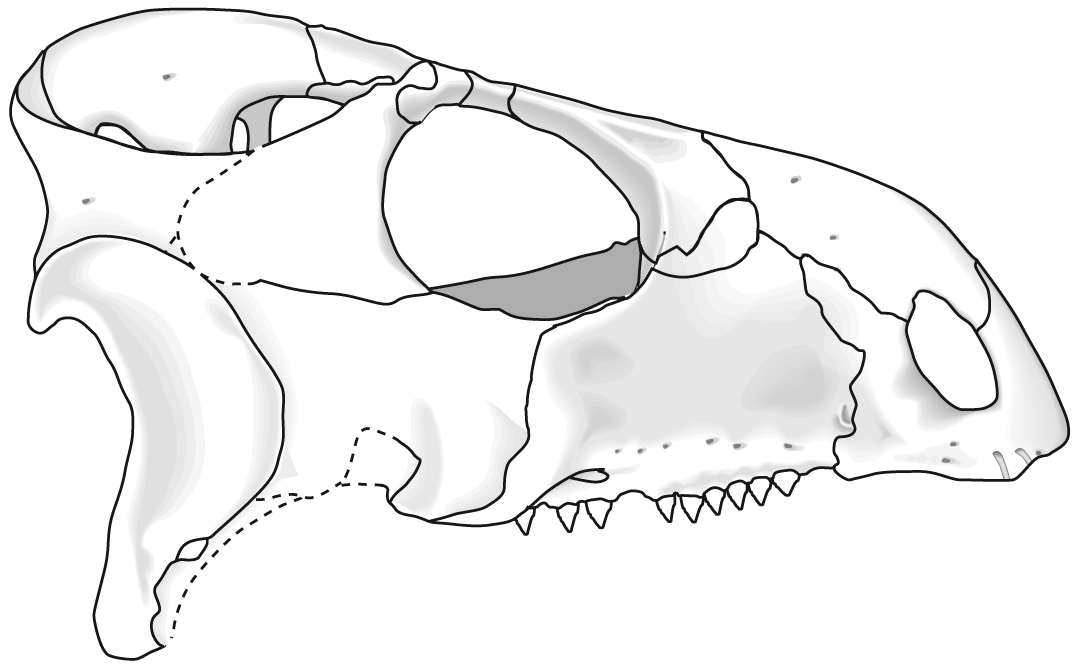
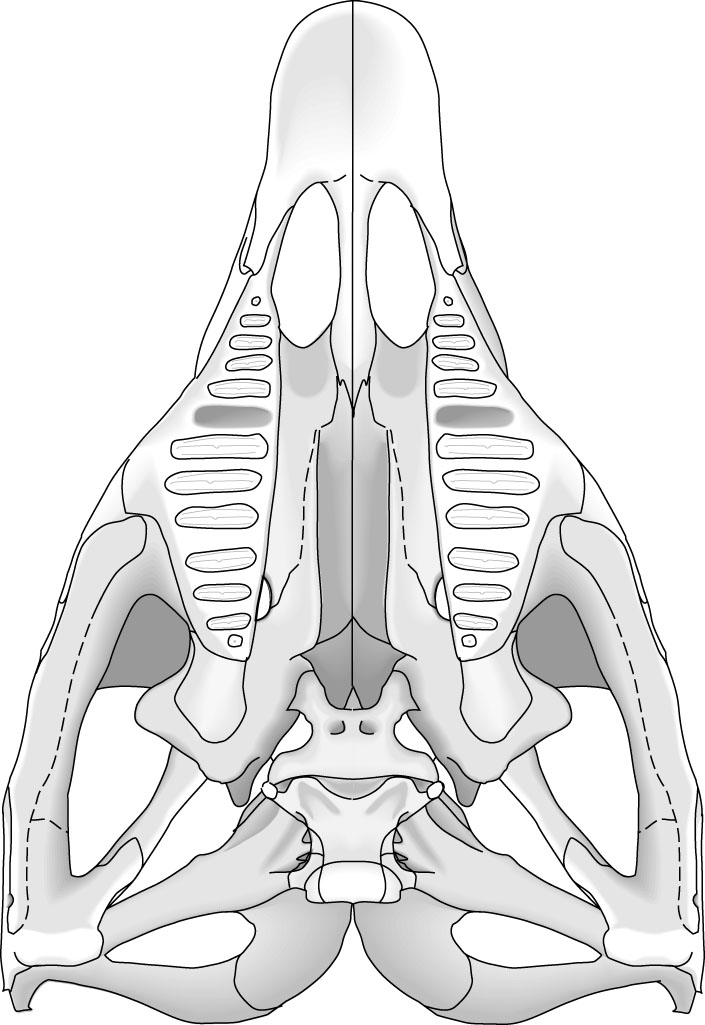
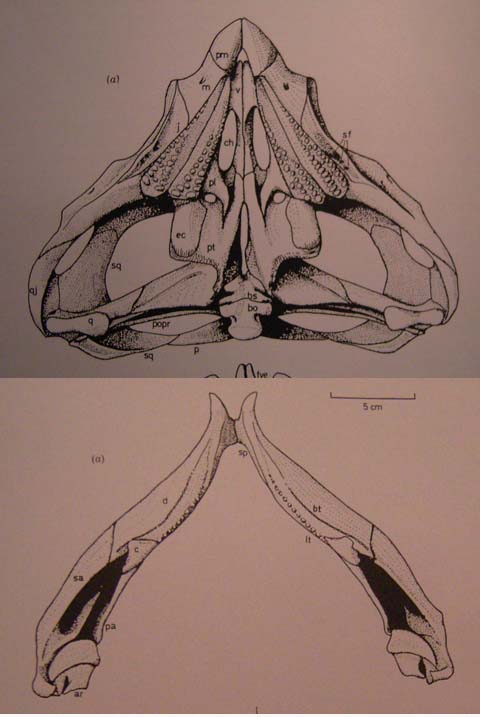
Rhynchosauridae: Min ∇ (Rhynchosaurus, Stenaulorhynchus, Hyperodapedon) (Dilkes, 1998). Skulls of all members were extremely modified for cutting and grinding tough plant material. Cranial specializations ranged from significant in Rhynchosaurus (a stenaulorhynchine) to profound in Hyperodapedon (a hyperodapedontine.) See lateral and ventral view.
Synapomorphies of Rhynchosauria + Archosauria include:
- Subnarial foramen that is not bordered by premaxilla.
- Premaxilla slightly downturned with respect to maxillary tooth row.
- Palate reinforced by interlaced overlap of ectopterygoid and pterygoid, and ectopterygoid contact with jugal.
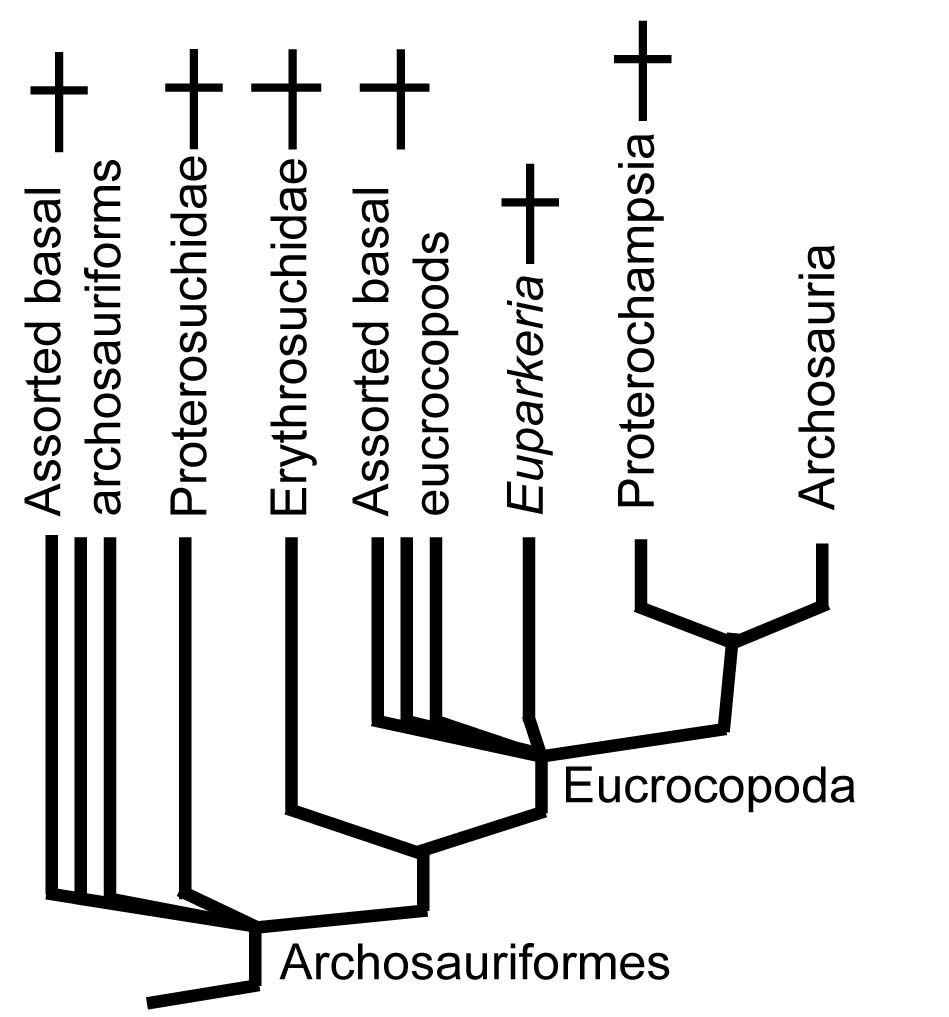
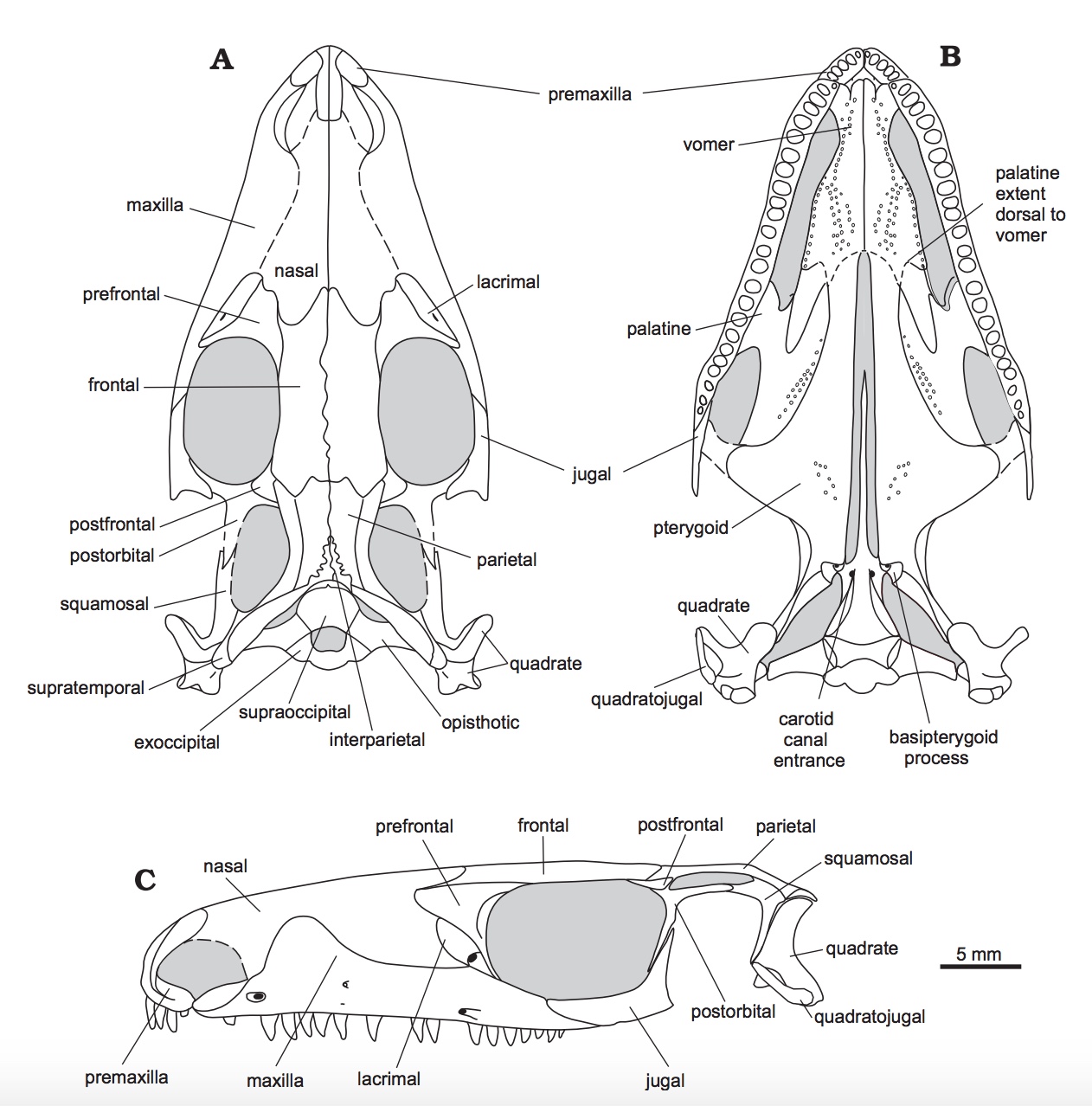
Crown-group Archosauria is nested inside Archosauriformes, a clade containing both Archosauria and critters that would, in the in the pre-Phylogenetic Systematics days, have been called archosaurs based on general similarity, but today are seen to lack some synapomorphy of Archosauria.
Major taxa within Archosauriformes include:
- Proterosuchidae - medium-sized terrestrial predators
- Erythrosuchidae - terrifyingly large terrestrial predators
- Proterochampsia - an odd assortment of medium-small terrestrial and aquatic predators
- Archosauria - The last common ancestor of birds and crocs, and all of its descendants.

Significant synapomorphies of Archosauriformes are not subtle:
- Antorbital fenestra present
- Pineal foramen absent
- Midline postparietal dermal element present. Link to image. See red element at left.
- Teeth serrated
- Premaxilla strongly downturned
- Reestablishment of infratemporal bar
- Mandibular fenestra present
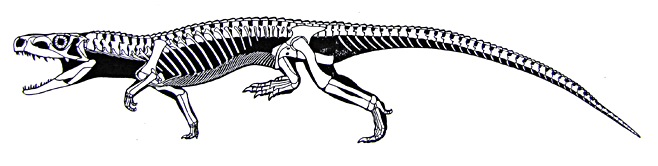
- Paired osteoderms down the midline of the back (See illustration of Euparkeria) provide mechanical strength to trunk.
Beyond this, we see some general tendencies:
- Increased size
- Fore and hindlimbs that assume a semi-sprawling posture, in contrast to other sauropsids considered.
Until recently, a big morphological gulf separated Rhynchosauria from the next branch up - Archosauriformes. These are the classic archosaur-looking animals of the pre-cladistic era. Pinheiro et al., 2017, however, described a lone Early Triassic skull that is midway across this gulf - Teyujagua paradox.

In size and general configuration, it resembles proper archosauriformes, and its presence breaks up what is otherwise a very long list of archosauriform synapomorphies. It has:
- Reduced pineal foramen
- Teeth serrated
- Premaxilla slightly downturned
- Infratemporal bar is just barely incomplete
- Mandibular fenestra present
It definitely lacks:
- Antorbital fenestra
- Complete closure of pineal foramen
- Midline postparietal dermal element.
- Complete infratemporal bar

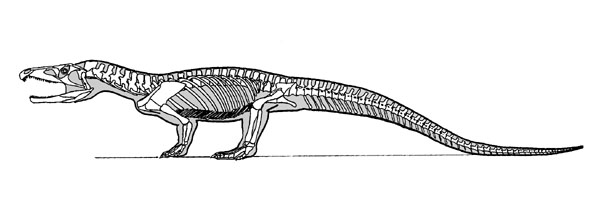
Proterosuchidae:
(Late Permian to Early Triassic) Arguably the most basal: Proterosuchidae, including Proterosuchus and Chasmatosaurus. Present prior to the Permo-Triassic extinction and common immediately after it - a classic "disaster taxon." (Cf. the dicynodont Lystrosaurus.) Probably similar in life style to crocodilians, but without strong specializations for aquatic life.
Synapomorphies of Proterosuchidae:
- Diastema between premaxillary and maxillary tooth rows.
- Maxilla extends past posterior orbit border

Erythrosuchidae:
(Early to Middle Triassic) The first archosauromorph apex predators. During the Middle Triassic they held the role that would be taken by proper archosaurs during the Late Triassic, and parallel some of their derived features. If you're thinking that the head is improbably large, you are not wrong. Uniquely among archosauromorphs, the heads of erythrosuchids have been shown by Bestwick et al., 2022 to scale with positive allometry.
Synapomorphies of Erythrosuchidae:
- Anterior third of maxillary tooth row abruptly upturned.
- Maxilla extends past posterior orbit border
- Loss of palatal teeth
Synapomorphy with Archosauria:
- A "triradiate" pelvis in which the pubes and ischia project anteroventrally and posteroventrally.
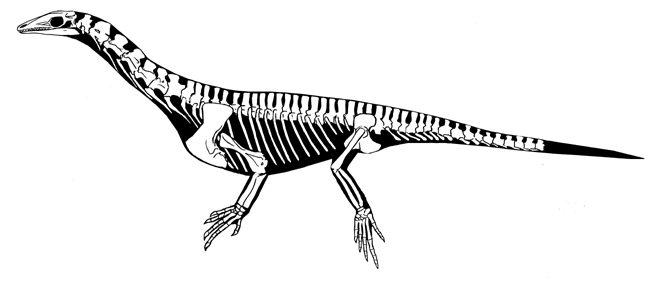
Eucrocopoda:
(Late Early Triassic to Quaternary) Max ∇ (Passer ~ Erythrosuchus). Derived features are concentrated in the femur, which has:- Inturned head
- Fourth trochanter - a crest for insertion of caudofemoralis muscles.
- A facultative biped
- capable of an "improved" stance.
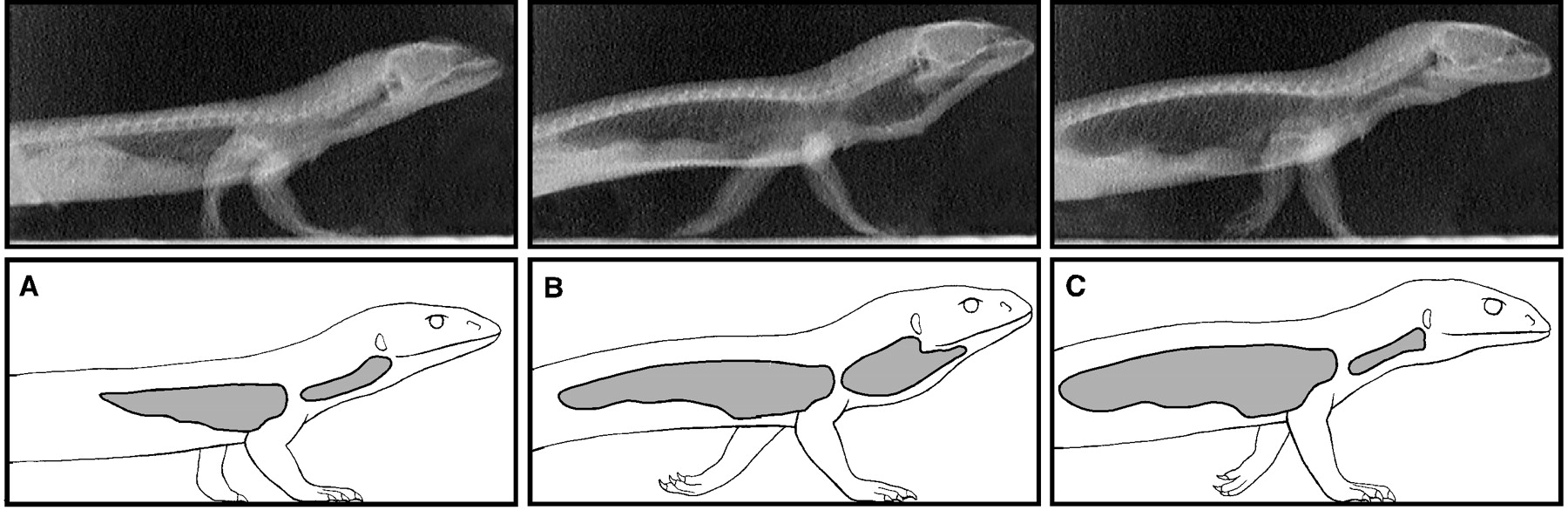
A quantitative analysis of Euparkeria's hip joint mobility by Demuth et al., 2020 suggests that it is capable of a wide range of postures ranging from an intermediate semi-upright to fully upright. It's obliquely angled ankle joint, however, suggests that it didn't typically assume a full upright stance.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
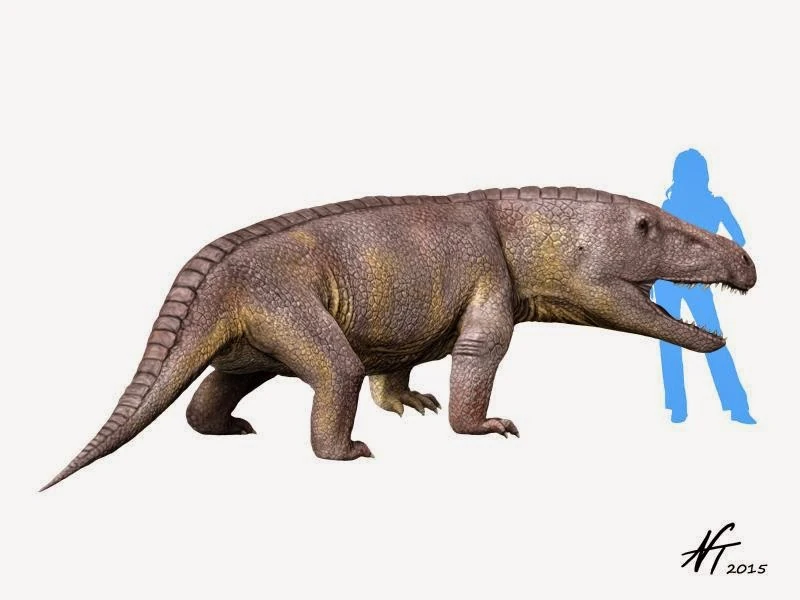{kind=link}
{kind=link}
{kind=link}
{kind=link}
Proterochampsia:
(Middle Triassic to Late Triassic) Max ∇ (Proterochampsa ~ Euparkeria, Erythrosuchus, Passer, Crocodylus) (Nesbitt, 2011)An ecologically diverse assortment of smallish eucrocopods. Synapomorphies include:
- Nares face dorsally
- Transverse profile of snout between nares and orbits is gently rounded (as opposed to the oreinorhinal configuration of other archosauriformes.)
- Maxilla contacts prefrontal
- Tooth-bearing part of dentary ventrally curved.
- Metatarsal V lacks a hooked proximal end (reversal).
- Proterochampsidae: (Early Late Triassic) Medium-sized fresh-water aquatic predators with flattened skulls that arguably lived as modern crocodilians do. (See Proterochampsa right)
- Dosweliidae: (Late Middle Triassic to Late Triassic) Medium-sized predators occupying a range of ecologies. Although united by technical synapomorphies like:
- Maxilla extends posteriorly past orbit
- Squamosal form posterior border of supratemporal fenestra,
- Extensive square scutes, including on limbs.
- Doswellia: A gracile terrestrial predator with an oddly-configured, boxy trunk region and long slender limbs.
- Vancleavea: An angilliform fresh water aquatic predator with predator whose body is covered with imbricating scutes. Its skull is strongly laterally compressed and it has secondarily closed its antorbital and supratemporal fenestrae.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Proterochampsia and Archosauria are united by subtle synapomorphies of the vertebral column and coracoid.
Of the groups referred to in this lecture, only two:
- Choristodera
- Archosauria
Additional reading:
- Michael Benton, 1985. Classification and phylogeny of the diapsid reptiles. Zoological Journal of the Linnean Society 84, 97-164.
- Jordan Bestwick, Pedro L. Godoy, Susannah C. R. Maidment, Martin D. Ezcurra, Mia Wroe, Thomas J. Raven, Joseph A. Bonsor, Richard J. Butler 2022. Relative skull size evolution in Mesozoic archosauromorphs: potential drivers and morphological uniqueness of erythrosuchid archosauriforms. Palaeontology 65(3) e12599.
- M. Borsuk-Bialynicka and S. Evans, 2009. A long-necked archosauromorph from the Early Triassic of Poland. Palaeontologica Polonica 65: 203-234.
- Oliver E. Demuth, Emily J. Rayfeld, and John R. Hutchinson, 2020. 3D hindlimb joint mobility of the stem-archosaur Euparkeria capensis with implications for postural evolution within Archosauria. Ntureresearch Scientific Reports.
- David Dilkes, 1998. The early Triassic rhynchosaur Mesosuchus browni and the interrelationships of basal archosauromorph reptiles. Philosophical Transactions of the Royal Society of London B Biological Sciences 353(1368): 501Đ541.
- Martin Ezcurra, 2016. The phylogenetic relationships of basal archosauromorphs, with an emphasis on the systematics of proterosuchian archosauriforms. PeerJ April 28, 2016 PubMed 27162705.
- John Flynn, Sterling Nesbitt, Michael Parrish, Lovasda Ranivoharimana, Andre Wyss, 2010. A new species of Azendohsaurus (Diapsida: Archosauromorpha) from the Triassic Isalo Group of southwestern Madagascar: cranium and mandible. Palaeontology 53(3), 669Đ688.
- Jacques Gauthier, 1984. A cladistic analysis of the higher systematic categories of the Diapsida. [PhD dissertation]. Available from University Microfilms International, Ann Arbor, #85-12825, vii + 564 pp.
- Chun Li, Olivier Rieppel, Michael C. LaBarbera, 2004. A Triassic Aquatic Protorosaur with an Extremely Long Neck. Science 305(5692) 1931.
- Sterling Nesbitt, 2011. The early evolution of archosaurs : relationships and the origin of major clades.. Bulletin of the American Museum of Natural History. 352.
- Sterling Nesbitt, John J. Flynn, Adam C. Pritchard, J. Michael Parrish, Lovasoa Ranivoharimanana, Andre R. Wyss, 2015. Postcranial osteology of Azendohsaurus madagaskarensis (?Middle to Upper Triassic Isalo Group, Madagascar) and its systematic position among stem archosaur reptiles. Bulletin of the American Museum of Natural History. 398: 1Đ126.
- Pinheiro, F., FranŤa, M., Lacerda, M. et al. 2017. An exceptional fossil skull from South America and the origins of the archosauriform radiation. Sci Rep 6, 22817 (2016). https://doi.org/10.1038/srep22817
- Silvio Renesto and Franco Saller 2018. Evidences for an aquatic lifestyle in the Triassic diapsid reptile Tanystropheus. Rivista Italiana di Paleontologia e Stratigrafia vol. 124(1): 23-34
- Sengupta, S.; Ezcurra, M.D.; Bandyopadhyay, S. 2017. A new horned and long-necked herbivorous stem-archosaur from the Middle Triassic of India. Nature: Scientific Reports. 7: 8366.
- Tiago Simões, Oksana Vernygora, Michael W. Caldwell & Stephanie E. Pierce . 2020. Megaevolutionary dynamics and the timing of evolutionary innovation in reptiles. Nature Communications volume 11, Article number: 3322 (2020).
- Stephan N. F Spiekman, James M. Neenan, Nicholas C. Fraser, Vincent Fernandez, Olivier Rieppel, Stafania Nosotti, Torsten Scheyer, 2020. The cranial morphology of Tanystropheus hydroides (Tanystropheidae, Archosauromorpha) as revealed by synchrotron microtomography. PeerJ 8:e10299.
- Stephan N. F Spiekman, Nicholas C. Fraser, Torsten M. Scheyer, 2021. A new phylogenetic hypothesis of Tanystropheidae (Diapsida, Archosauromorpha) and other "protorosaurs", and its implications for the early evolution of stem archosaurs. PeerJ 9:e11143.
- Spiekman SNF, Wang W, Zhao L, Rieppel O, Fraser NC, Li C. 2024. Dinocephalosaurus orientalis Li, 2003: a remarkable marine archosauromorph from the Middle Triassic of southwestern China. Earth and Environmental Science Transactions of the Royal Society of Edinburgh. 2023;114(3-4):218-250. doi:10.1017/S175569102400001X
- Hans-Dieter Sues, 2003. An unusual archosauromorph reptile from the Upper Triassic Wolfville Formation of Nova Scotia. Canadian Journal of Earth Science 40: 634-649.
- Hans-Dieter Sues and Rainer Schoch, 2023. A new Middle Triassic (Ladinian) trilophosaurid stem-archosaur from Germany increases diversity and temporal range of this clade. Royal Society Open Science March, 2023.
- Jordan Bestwick, Pedro L. Godoy, Susannah C. R. Maidment, Martin D. Ezcurra, Mia Wroe, Thomas J. Raven, Joseph A. Bonsor, Richard J. Butler 2022. Relative skull size evolution in Mesozoic archosauromorphs: potential drivers and morphological uniqueness of erythrosuchid archosauriforms. Palaeontology 65(3) e12599.