The Pattern of Evolution - Phylogenetic Systematics Review
John Merck
The Phylogenetic System of Taxonomy: Any system of taxonomy, scientific or folk, has:
- An organizing principle
- An internested hierarchy of categories
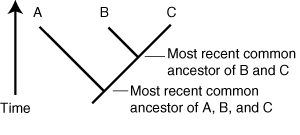
Graphic representation of a the tree
of evolution of three taxa, A, B, and C.
- Lineages propagating through time are shown as line segments.
- Speciation events are Y-shaped bifurcations
- Terminal taxa for which we have actual information are indicated at the ends of branches.
Definitions:
Monophyletic groups: In phylogenetic systematics, taxonomic groups are defined strictly in terms of the non-arbitrary criterion of descent from a common ancestor. Such taxa are called monophyletic groups.
-
Memorize this definition: A monophyletic group is an ancestor and ALL of its descendants. E.G. in the cladogram at right, the last common ancestor of B and C and all of its descendants would be monophyletic.
- Paraphyletic groups: An ancestor and some but not all of its descendants. In the cladogram at right, the last common ancestor of A and B and all of its descendants except for C and the branch leading to it would be paraphyletic.
- Polyphyletic groups: A group of organisms which fails to include at least some of their common ancestors. In the cladogram at right, the a group that contained A and C but omitted their last common ancestor would be polyphyletic.
Note: You may be familiar with two types of non-monophyletic groups:
Note carefully: Only monophyletic groups are based exclusively on natural, non-arbitrary criteria. When we define a paraphyletic group, we must arbitrarily decide which descendants to exclude. In the case of polyphyletic groups, we must decide which ancestors to leave out.
Hypotheses of Phylogeny
If God were to hand us the true phylogeny, and our only task were to read it and construct taxonomic system accordingly, our lives would be easy. Instead, we must somehow reconstruct phylogeny by making observations and testing hypotheses. This is where the "modification" side of "descent with modification" comes in. As lineages evolve, the characters of their members change. I.e. they go from ancestral to derived states.
- Sometimes we refer to cladograms as "trees." It helps to think of them that way. Imagine marker dye injected into branch. In the tree of evolution, the derived character states play the role of the marker dye.
- There is a particular type of derived character state that we are particularly interested in. Synapomorphy = Derived character state shared by different lineages.
Note: Just as we have disphonious cladobabble describing different types of taxonomic groups, we have it for characters, too:
- Plesiomorphy: The ancestral character state - inherited from distant ancestors. E.G. From the point of view of Primates, having five fingers is a plesiomorphy. (Symplesiomorphies are "shared ancestral states.")
- Apomorphies: A derived (evolved) character state. E.G. From the point of view of Primates, having an opposable thumb is an apomorphy.
- Synapomorphies: are "shared derived states." E.G. the presence of an opposable thumb is a synapomorphy of humans, chimpanzees, monkeys, lemurs, and other lineages of primate.
- Autapomorphies: Unique derived character state. E.G. The enlarged braincase of humans is an autapomorphy, not shared with other animals.
Let's see how this works in a simple cladistic analysis of some imaginary beetles. We assume that they are related somehow, but we don't know if B shares a more recent common ancestor with C or A, or if C and D are more closely related to one another than to B.
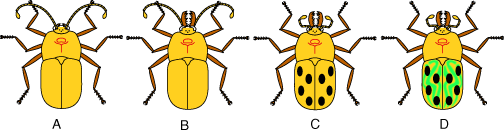
- The first thing we do is notice how they are different.
- Three have big jaws while one does not.
- Two have long antennae while the others have short ones.
- Two have plain wing covers while the others have spotted ones.
- One has stripes
- We compile this information in a table called a taxon-character matrix.
Character
1. Large jaws present
2. Small antennae present
3. Spots present
4. Stripes present
A
0
0
0
0
B
1
0
0
0
C
1
1
1
0
D
1
1
1
1
Outgroup taxon: This matrix records whether the observed state for each taxon is ancestral or derived. How do we know? You may have noticed that we we haven't had much to say about taxon A. In this analysis, A is the outgroup taxon. This is a beetle that, on the basis of some prior information, we can assume is more distantly related to beetles B, C, and D than any of them are to one another. Maybe we found it fossilized in amber. The outgroup is our standard for what is derived and what isn't, in that anything we see in it, we assume to be the ancestral state. Incidentally, because it has smaller mandibles than the others, I've included a "large mandible" characteristic in the matrix.
Tree 1: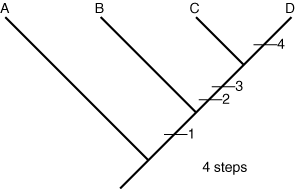
Tree 2: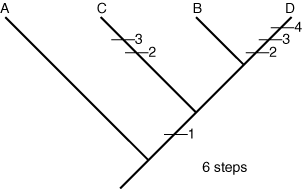
Tree 3: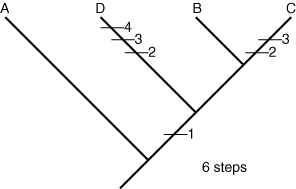
- We now compare every possible evolutionary tree arrangement by mapping onto them the simplest possible distribution of each character state change. Because A is presumed to be the outgroup, its ancestral lineage always branches off first, no matter what.
The evolutionary novelty of large mandibles, for example, can be explained most simply assuming that it appeared once before the last common ancestor of A, B. and C, who simply inherited it. Short antennae are a different story. The outgroup has long antennae, so we assume this is the ancestral state, but beetle C retains them also. In some tree arrangements, this implies that short antennae they evolved independently in A and B. After mapping the character changes onto the trees, we count them up. Tree 1 implies a minimum of six changes, tree 2 suggests a minimum of eight and tree 3 would require a minimum of seven. Each of these arrangements is a plausible hypothesis of evolutionary history. Now, we must choose the best hypothesis. To do that, we use the principle of parsimony, which holds that the simplest solution is usually the best. For us, the simplest hypothesis is the one that invokes the fewest character state changes. That is tree 1, with only six changes. Is this the true tree? God only knows. The best we can say is that it is our best approximation based on current knowledge.
- Most reasonable hypothesis of phylogeny based on available data, not necessarily truth.
- Model of character evolution: We can identify likely points of character state change, and distinguish convergent characters from unidirectionally evolving ones. Note: Not all characters evolve concordantly. Some experience reversals or convergences. The phylogenetic analysis helps us identify these homoplastic characters.
Feeling vulnerable? For more review see:
What Phylogenies Tell Us
Hypotheses of the pattern of evolution: Duh. But it does not stop here. For all that the pattern of evolution is a worthwhile thing to know, it is not an end in itself. Reconstructing Missing Information: A robust phylogeny can enable us to develop hypotheses about the state of characters in extinct organisms for which we have no direct evidence.
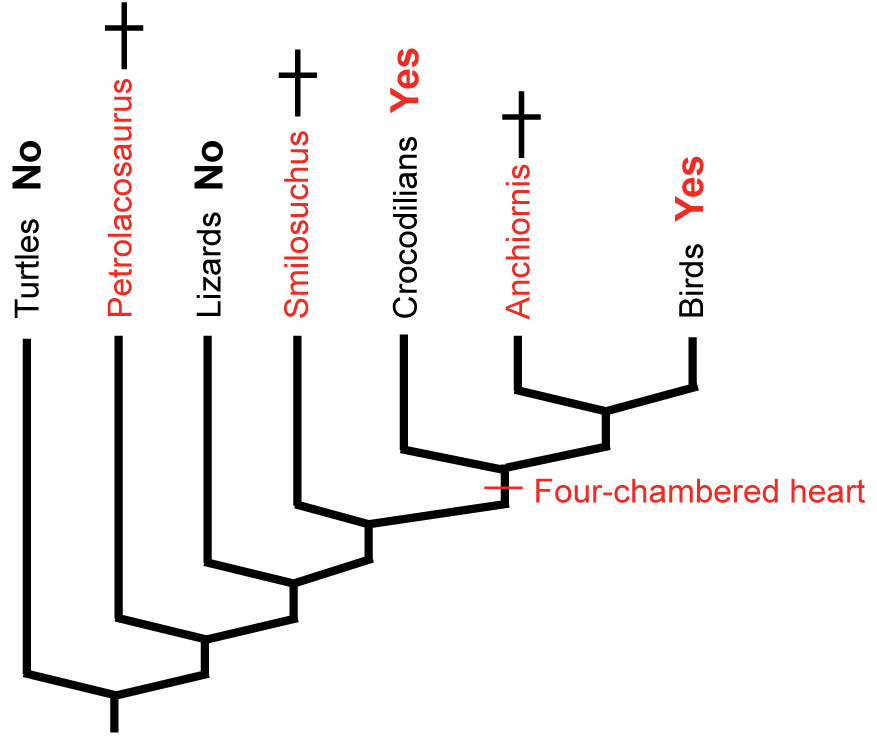
- Type I Inference: the bird-like theropod Anchiornis is bracketed by birds and crocodilians, both of which have the derived character. With no contrary positive evidence, the simplest assumption is that Anchiornis had it also.
- Type II Inference: The archosauriform Smilosuchus, not quite an archosaur, is bracketed by crocodilians and birds above, and squamates below. Crocs and birds have the derived character, squamates don't. Thus, we are less secure than above in inferring it in Smilosuchus, and even less secure the farther from the Archosaurian node we get. Still, the presence of some sort of hard tissue correlate of that trait, or a strong biomechanical argument for it might increase our confidence.
- Type III Inference: The basal diapsid Petrolacosaurus is bracketed by squamates and turtles, neither of which have the derived character. Our confidence in
its presence in the extinct form is very low. We would need strong positive fossil evidence to argue for its presence.



Phylogenies meet the Rock Record
Phylogenies and Biostratigraphy: Traditional biostratigraphers - Geologists who use the fossil record to date sedimentary rock units tend to be literal-minded souls who think that either fossils of a taxon are e present at a particular time, or they aren't. What if a group of organisms is known only from relatively rare specimens. I we reconstruct a robust phylogeny, it might help us identify ghost lineages intervals of time in which a lineage ought to have been present but for which no fossils have been found. This:
- tells us when we ought to be cautious about literal interpretations of the rock record.
- enables us to approximate minimum divergence ages for evolving lineages.
Ghost Lineages and minimum divergence ages: When we know that two taxa are sister taxa (descendants of the same recent common ancestor), we in essence know that they originated at the same point in geologic time - the time of their last common ancestor and the speciation event that gave rise to them. Say we know one of these taxa from 100 million year old rocks, and the other from 90 million year old rocks. Even without seeing a fossil, we know that the second group must have representatives dating back at least to 100 million years, simply from its sister-taxon relationship with the other. 100 million years is the minimum divergence age of the monophyletic group formed by the two total groups. A lineage whose existence can be inferred from the cladogram, but which is not known from actual fossils is called a ghost lineage. The examination of ghost lineages should allow biostratigraphers to refine their models of the stratigraphic ages of organisms.

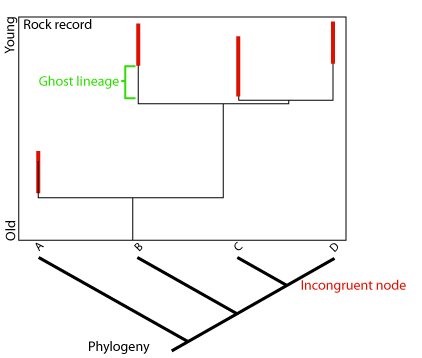
- the creatures had poor fossilization potential
- that their fossils have been misidentified
- that our reconstruction of phylogeny is wrong.
Crown-group definitions: A special case of the node based definitions in which the anchor taxa are living. Thus:
- "the last common ancestor of of Homo sapiens and Homo habilis and all of its descendants" is node-based but not a crown group, whereas
- "the last common ancestor of humans and gorillas and all of its descendants" is node-based and also a crown group.
Utility:
- Utility of node-based definitions: The set of synapomorphies we use to diagnose (i.e. identify) a node-based group is acquired during evolutionary change in the lineage leading up to the group's last common ancestor. Thus, we can be certain that "doodads," the synapomorphy of Doodadichthyes, were present in that common ancestor.
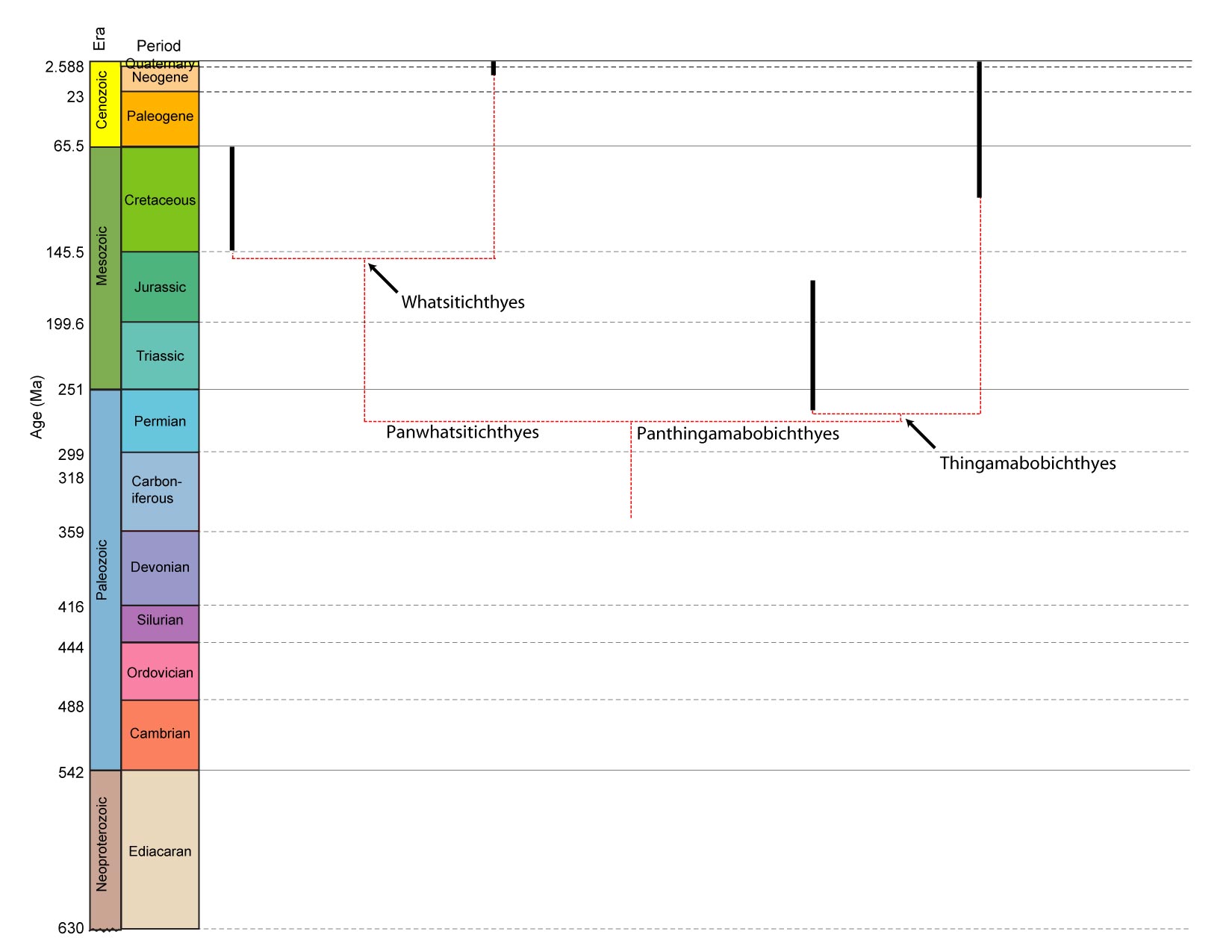
- Weakness of node-based definitions: We represent relative time on the cladogram, however it is impossible precisely to compare the sequence of events on different branches. Thus, if we wanted to know whether the last common ancestor of Thingamabobichthyes or Whatsitichthyes lived first, we would be out of luck. We can only say that both lived after the last common ancestor of Doodadichthyes.
Utility:
- Utility of Total-group definitions: Unlike with node-based sister taxa, we can readily compare the ages of total-group sister taxa. Panthingamabobichthyes and Panwhatsitichthyes both arose from a single event - the speciation event that split the last common ancestor of Doodadichthyes into two lineages. The two groups are, by definition, of equal age. This fact can become very convenient if one group has a copious fossil record and the other doesn't.
- Weakness of total-group definitions: But now we have a problem. The synapomorphy of Thingamabobichthyes is "thingamabobs." But what about Panthingamabobichthyes? We know that somewhere in the lineage leading to the last common ancestor of Thingamabobichthyes, thingamabobs appear, but where? That we don't know. In fact, it is most probable that if we could look at the very first panthingamabobichthyians, they would not yet have acquired thingamabobs. In effect, total-groups can't be diagnosed with synapomorphies.