


"The Dinosaurs, having the same thoracic structure as the Crocodiles, may be concluded to have possessed a four-chambered heart; and, from their superior adaptation to terrestrial life, to have enjoyed the function of such a highly-organized centre of circulation to a degree more nearly approaching that which now characterizes the warm-blooded Vertebrata." -- Sir Richard Owen, 1842, "British Fossil Reptiles"
BIG QUESTION: Were dinosaurs warm-blooded?
The Hot-Blooded Dinosaurs: Reconstructing Dinosaur Physiology
Among modern vertebrates, some gross generalizations:
Birds and
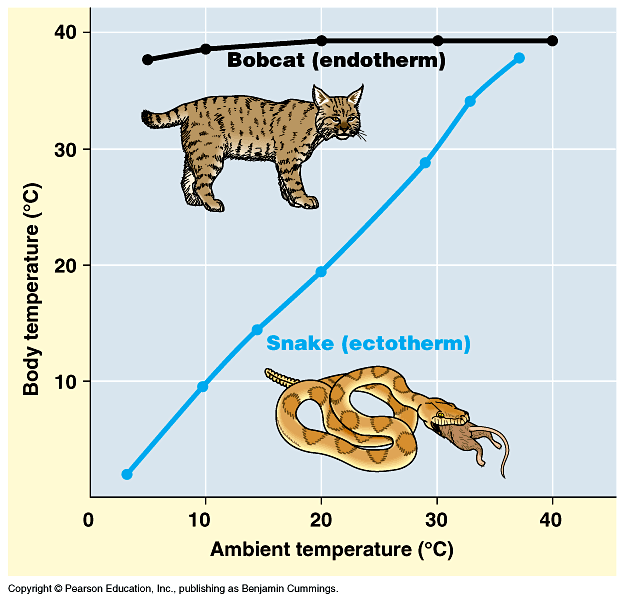
mammals are warm-blooded; that is, they are warmer than the environment around them in typical temperate and colder environments. Crocodilians, lepidosaurs, turtles, amphibians, most fish, and almost all invertebrates are cold-blooded: their bodies are generally only about as warm as the general environment around them, so consequently they feel cool to the touch outside of tropical situations; in contrast, warm-blooded animals have temperatures largely independent of the outside temperature,
so they feel warm to the touch. Need to be precise as to definitions of terms. "Warm-blooded" and "Cold-blooded" actually encompass several different (although related) topics:
A typical cold-blooded animal is an ectothermic bradymetabolic poikilotherm: needs to get its energy from the sun and fluctuates with external environment (but can moderate fluctuations by moving from sunlight to shade and vice versa); however, needs very little food (snakes can go weeks without feeding, for example). Cold blooded animals become torpid at night and in colder weather.
A typical warm-blooded animal is an endothermic tachymetabolic homeotherm: its body temperature is stable and activity levels can remain high for long periods of time, at night, and in colder weather; however, needs a LOT of food or will die (imagine the effects of not feeding a cat or dog for weeks!).
Additional issues to consider:
Why evolve such an expensive trait as endothermy? Some suggestions have included:
Note that it is not just mammals and birds that are "warm-blooded". For example, tunas, billfish (sailfish, swordfish, marlins), lamniform sharks (like great whites and makos), boid snakes (pythons, etc.; but only while brooding), and certain plants (which aren't "blooded" as such, but some can emit internally-generated heat).
When dinosaurs were first discovered, they were interpreted as being no more than gigantic cold-blooded lizards. However, as early as 1842 Owen (in the very paper in which he named "Dinosauria") speculated that dinosaurs may have been warm-blooded like mammals. During most of the 20th Century the model of dinosaurs as cold-blooded returned. Work by John Ostrom (of Yale University) and his colleagues and students (especially Robert Bakker) presented new information that dinosaurs were in fact warm-blooded. This hypothesis generated considerable research (both in support and in attempts to falsify it): this change in thinking about dinosaurs and renewed interest in dinosaurian studies has been termed the "Dinosaur Renaissance".
Among the lines of evidence supporting dinosaurian warm-bloodedness:
So You Want To Be An Endotherm?
Let's consider the equations of life. First, the aerobic respiration equation, the primary means by which animal cells operate:
C6H12O6 + 6O2 yields 6CO2 + 6H2O + Energy
(That is, food (glucose) plus oxygen yields waste carbon dioxide and waste water, plus energy).
If an animal's cells can't get enough oxygen, there is a second way of getting energy: the anaerobic respiration equation:
C6H12O6 yields 2C3H6O3 + Energy
(That is, food yields lactic acid plus energy (although much less than the aerobic respiration.) Lactic acid itself needs oxygen to break down, so you cannot run on anaerobic respiration for very long.
If you want to evolve endothermy, you need to:
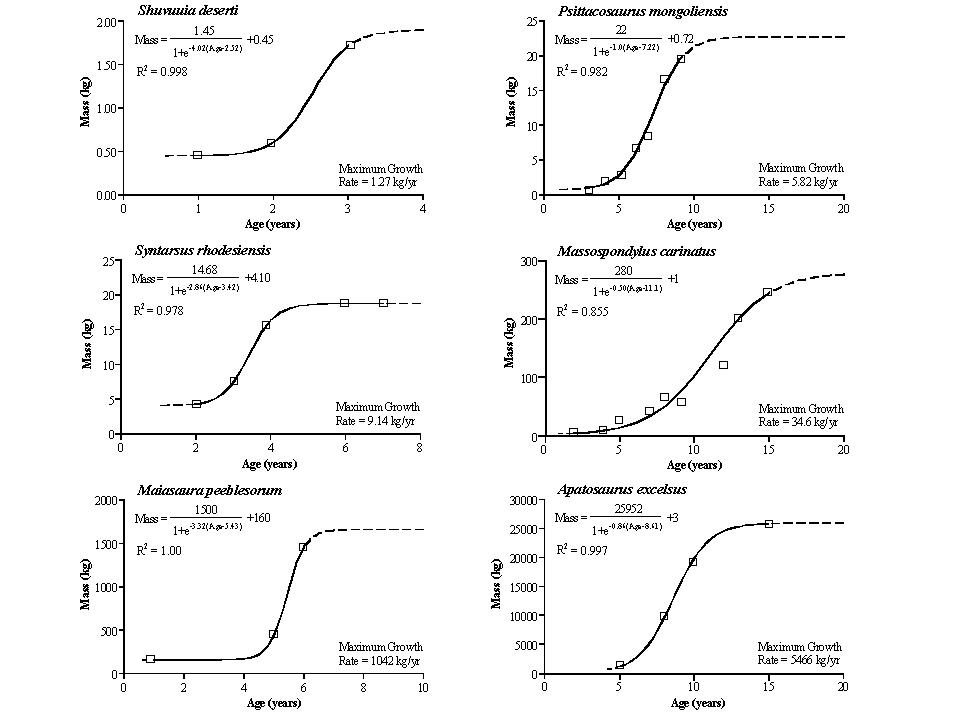
So, where do we stand on dinosaur metabolism?
What would be necessary to justify the above observations?
Is there evidence for these features in dinosaurs? YES!:
Heat enters and exits the body of the surface area, but it is generated internally by the mass of the body (in other worlds, in volume.) So the the ratio of the surface area to the volume is a metric of how quickly (or slowly) heat would leave the body. But from geometry, as linear dimensions double, the surface area goes up by squares, and the volume by cubes:
| Side Length | Surface Area (SA) | Volume (V) | SA/V |
|---|---|---|---|
| 1 | 6 | 1 | 6/1 = 6 |
| 2 | 24 | 8 | 24/8 = 3 |
| 3 | 54 | 27 | 54/27 = 2 |
| 4 | 96 | 64 | 96/64 = 1.5 |
Thus, as size increases, SA/V decreases. Therefore, with bigger body size it takes longer and longer for heat to be lost or gained.
Indeed, very large animals potentially become gigantothermic: homeothermic (stable body temperature) without having the energy costs of endothermy! But did dinosaurs do this? A recent (2020) analysis found that body temperatures of dinosaurs were about the same regardless of whether they were small, medium size, or gigantic. And only the gigantic ones were temperatures that would have been the result of gigantothermy.
Dinosaurs had such a huge size range, that their thermoregulation ranged from needing to keep heat in to needing to dump it:
Keeping the Heat In; Insulation Issues: One problem that small-bodied organisms encounter is the fact that a small organism has a much higher surface area/volume ratio than a large one. Because of this, small animals tend to lose heat much faster than big ones. In contrast, large animals lose heat to or gain heat from the outside world only gradually. This has led some people to suggest the possibility that large dinosaurs exhibited "gigantothermy": effective homeothermy achieved because of large body size. However, this would not apply to small-bodied dinosaurs: either adults of small species or the hatchlings of giants. So how could these keep warm?
There is strong evidence that many--if not most--of the theropods had a fuzzy body insulation over the body: true feathers in the advanced groups, simpler "protofeathers" in the primitive ones. Such fuzz would help keep the warmth in the body. In fact, this is the primary function of the fur of mammals, and one of the functions of body feathers in birds. The recent discovery of 1.4 t Yutyrannus demonstrates that even some giant theropods were fuzzy.
Recent discovery of the early Late Jurassic Chinese ornithischian Tianyulong and the similar aged Kulindadromeus of Siberia showed they too had a fuzzy body covering over at least part of its body! If this is found to be homologous to the protofeathers of tetanurine theropod saurischians it would suggest that the concestor of all dinosaurs was fuzzy, and that dinosaurs were thus fuzzy ancestrally! (In the case of Kulindadromeus, there are also also scales, plates, and additional bizarre tufted plates.) At present, however, there is enough uncertainty to make the homology between Tianyulong's fuzz, Kulindadromeus's diverse integument, and theropod protofeathers suspicious. (But do not be terribly surprised if in the future we discover that most dinosaurs were fuzzy to some degree or another! All we need is a fuzzy primitive sauropodomorph, and it is basically a done deal!)
Dumping Heat: Mechanisms to Remove Heat: Of course, most dinosaurs are not small. Because of the scaling issues discussed previously, large-bodied endotherms have no issue keeping heat in; indeed, they run the risk of overheating because of they cannot lose heat easily. So larger endotherms typically need adaptations to help regulate their body temperature (and most especially for the brain: brains cannot be uncooked.)
Do dinosaurs have any such adaptations. It seems that they do. For instance, some dinosaurs have conspicuous large sails or plates or frills or long necks or long tails that might have been used to dump waste heat.
More generally, there are other structures may have also been used to regulate temperature. The antorbital fenestra (also the promaxillary and maxillary fenestrae of various theropods) housed soft tissue air sacs. These air sacs may have been useful to transport waste heat. Also, many larger dinosaurs have enlarged and/or elaborate nasal regions. The blood vessel-rich tissues of the nose would also have been useful in dumping waste heat.
The enlarged narial regions may support tissues for a different function: recovery of moisture. In living endotherms, rapid rate of respiration would dry out lungs if not for some specialized tissues called nasal turbinates: these are or small in modern ectotherms, but extremely large in mammals, where they support thin tissues which trap moisture going out, and rewets on way back in. Nasal turbinates were fairly large in many birds, but are cartilage rather than bony. (Also, some birds seem to rely on air sac system for this purpose).
Most non-avian dinosaurs do not show much evidence for internal nasal turbinates, but the air sac system and/or tissues in the enlarged narial regions of bigger dinosaurs may have served this function
An secondary advantage of using respiratory turbinates to dump heat is that it helps direct some of the air flow onto the part of the nasal chamber associated with olfaction:
It has just recently been recognized that the expanded frontopareital fossa (an expanded region on the top of the skull of dinosaurs) were filled by vascular tissue and fat rather than by muscle. So these areas may have helped to radiate heat to keep the brains of dinosaurs cool (they seem to have that function in modern crocodilians):
Warm-blooded Protocrocs?: Most studies assume that endothermy evolved sometime after the bird lineage (Ornithodira) and the crocodilian lineage (Pseudosuchia) diverged from each other. This is because crocodilians are ectotherms, as are all the next several outgroups (lepidosaurs, turtles). However, what if crocodilians were not ancestrally ectotherms, but instead reverted to a cold-blooded physiology from warm-blooded ancestors?
There is some evidence that this is the case:
The Facts About Dinosaurs & Feathers:
5 Things You Should Know About Dinosaurs:

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}