GEOL 204 Dinosaurs, Early Humans, Ancestors & Evolution:
The Fossil Record of Vanished Worlds of the Prehistoric Past
Spring Semester 2017
Ancestors, "Missing Links", and Transitions: The Fossil Record of Speciation and Macroevolution
"Nothing makes sense in biology except in the light of evolution.", Theodosius Dobzhansky, 1963. "Biology, Molecular and Organismic" American Zoologist: 4: 443-452.
and
"Why then is not every geological formation and every stratum full of such intermediate links? Geology assuredly does not reveal any such finely-graduated organic chain; and this, perhaps, is the most obvious and serious objection which can be urged against the theory." -- Charles Darwin, 1859, Chapter 9 "On the Imperfection of the Geological Record", On the Origin of Species by Means of Natural Selection.
and
"Since we proposed punctuated equilibria to explain trends, it is infuriating to be quoted again and again by creationists -- whether through design or stupidity, I do not know -- as admitting that the fossil record includes no transitional forms. The punctuations occur at the level of species; directional trends (on the staircase model) are rife at the higher level of transitions within major groups."
, Stephen Jay Gould, 1980, The Panda's Thumb.
Sources of Variation
When looking at samples (fossils, modern specimens, etc.) of individuals, we will find similarities and differences. Do these similarities and differences represent simply variation? Or do they represent differences that significant taxonomically (that is, that are representative of distinct species, genera, etc.)?
There are several potential sources of variation:
Sexual dimorphism: different sexes are different sizes and shapes and have different structures
Ontogenetic (growth): babies look different from juveniles look different from subadults look different from adults (can be even more
extreme in animals that undergo metamorphosis, like amphibians and many insects)
Geographic: populations in different regions might have slightly different sizes, color patterns, proportions, behaviors, etc. For example, some biologists consider the populations of orangutans, tigers, African elephants, etc. as distinct species; others simply regard them as regional variants
Stratigraphic: lineages may shift in some traits or characteristics over time
Individual: one of the great "discoveries" of Darwin and Wallace, the recognition that no two individuals in a population are identical! (Before them, many people thought that there existed the perfect "type" of each kind of organism, and all variation is degeneration from that perfection. Darwin and Wallace showed that the variation is the reality)
The Species Problem
What, really, is a species? How do we recognize their boundaries? What is speciation (the literal "origin of a species)?
For most of the 20th century, the Biological Species Concept (BSC) of Ernst Mayr held sway: "an array of populations which are actually or potentially interbreeding, and which are reproductively isolated from other such arrays under natural conditions". (Almost certainly you learned some version of this in high school and BSCI classes.) It works pretty well for the first pass: it emphasizes isolation, and thus species would represent pools of shared genes which do not get mixed with their closest relatives.
But there are some problems:
Impossible to define for asexual species, unless each clone lineage is a separate species, despite being otherwise no different from each other than are two individuals are within sexually reproducing species
Extremely difficult to test in most wild populations, and (worst for paleontology) IMPOSSIBLE to test for fossils!
Existence of rare but real natural hybrids show that such isolation is not complete by any means
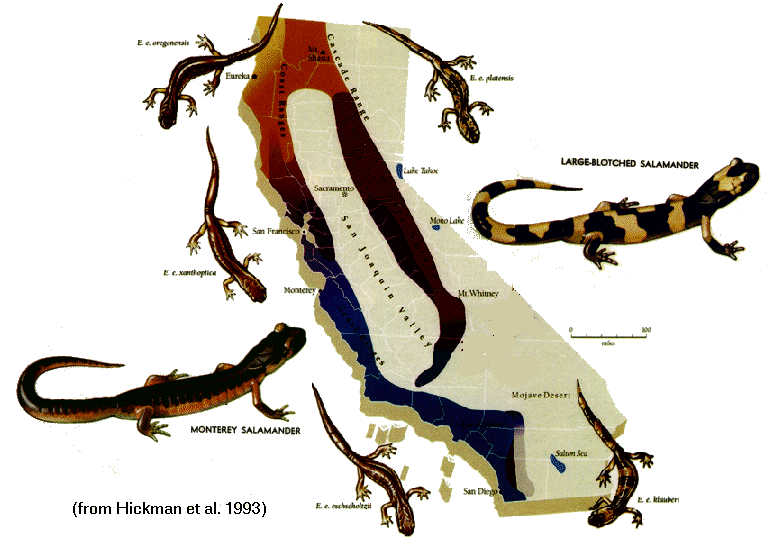
Existence of ring species shows that you can go through a continuum with no boundaries and yet reach conditions of genetic incompatibility between end members.
This is part of what is called the species problem: it is damned difficult to find a good, useful criterion that can be broadly applied to our ideas of what species are and how we recognize their boundaries. Biologists have tried to come up with hard-and-fast rules about how to recognize species, and although they have named many such "species concepts", these tend to cluster around two major different ideas:
Species are recognized by some shared aspect or attribute held in common by members of the species (similarity criterion)
Species are distinguished from other species by some attribute that makes them different from other (difference criterion)
Although related, they really aren't talking about the same things necessarily.
In real life, species do seem to have "fuzzy boundaries", and the distinction between different closely related species on the one hand and clusters of variation within a species are nearly impossible to tell. In fact, biologists go through shifts of fashion towards increasing splitting (the former idea) and lumping (the latter) over time. Currently the fashion is towards splitting: consequently, whereas in much of the 20th Century we recognized only one species each of African elephant, gorilla, orangutan, Nile crocodile, and orca, early 21st Century taxonomists recognize two or more. (On the flip side, dinosaur paleontologists seem to be following the opposite trend, lumping once-separate species and genera into each other).
As with many things, we run into problem with typological thinking: the idea that there are ideal types of things, and that we judge a specimens membership in a group by how well it conforms from that type. Instead, we find that variation is the reality. So we need to use population-based thinking. (Next lecture we will add tree-based thinking.)
Darwin's species concept is worth revisiting:
"I look at the term species as one arbitrarily given, for the sake of convenience, to a set of individuals closely resembling each other, and that it does not essentially differ from the term variety, which is given to less distinct and more fluctuating forms. The term variety, again, in comparison with mere individual differences, is also applied arbitrarily, for convenience sake." Chap. 2 of The Origin
"Species are only well-marked varieties" Chap. 4 of The Origin
"The only distinction between species and well-marked varieties is, that the later are known, or believed, to be connected at the present day by intermediate gradations, whereas species were formerly thus connected" Chap. 14 of The Origin
An important issue which is commonly forgotten comes out here: descendants are descendants of only a small part of any ancestral group! That is, entire species do not evolve into entire other species. Instead, only some small subset of any given species population is the ancestral group leading to a particular descendant. This points to several different aspects:
It is entirely possible for the ancestral species to still be around even though some subpopulation might give rise to a later species
There is no reason why for a given ancestral group that only a single subpopulation gives rise to descendants. Thus, we have a reason for divergence of multiple descendants from a single ancestral species
Physical geographic isolation of a subgroup and a change of environmental condition favoring evolution of distinct traits would greatly increase the chance in some small subgroup on the fringes (a peripheral isolate) to be most easily split off as a potential ancestor of a new species
Note: this relates to a common anti-evolutionary rant, which goes a long the lines of "if people evolved from monkeys, how come there are still monkeys". Ignoring lots of other problems with this statement (such as the fact humans didn't evolve from any living monkey species; that "monkeys" aren't one thing, but are a vast number of species; etc.), this misses important aspects of how evolution works! Just because some monkeys evolved into apes which evolved into humans does not require that ALL monkeys evolved into apes and ALL apes evolved into humans. Plus, it doesn't mean that monkeys were TRYING to evolve into humans, or DESTINED to do so.
(Here's a way to restate an analogy to this anti-evolution argument: "If [for example] your ancestor came from Ireland (or Norway, or India, or whatever), why are there still Irish/Norwegians/Indians/etc.?")
Speciation
Speciation is the process of the origin of a species. It doesn't happen immediately or instantaneously: it is indeed a process rather than an instantaneous event. (In fact, except in rare cases, it is unlikely that it you there during it that you would recognize it as such.)
Some aspects of the origin of species to consider:
Area of origin: Did the new species arise within the main range of the ancestral species (sympatric, "same homeland"); alongside the ancestral range, with no major barriers to gene flow (parapatric, "parallel homeland"); on the edges of the species range, with some substantial (but not necessarily total) barrier to gene flow (peripatric, "edge of the homeland"); or by either subdividing the original population or by isolating a part of it (allopatric, "other homeland")?
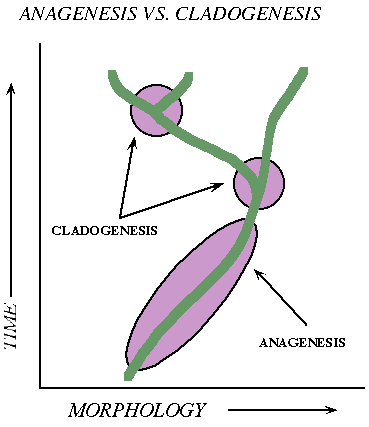
Trends or branches: Did the the new species arise by the main population itself shifting as a group (anagenesis, "no origin") or by splitting/subdivision of the lineage (cladogenesis, "branching origin")
During the 20th Century (especially during the first half), evolutionary biologists assumed the dominant trends were sympatry and anagenesis. However, as a better understanding of genetics was developed, some (including Mayr) argued that allopatry, peripatry, and parapatry (which all require cladogenesis) were actually more common.
The problem, of course, is that speciation takes time, and field biologists are unlikely to observe it. If only there were some sort of record of changes over time. Say, for example, a fossil record...
Creeps or Jerks: Phyletic gradualism vs. Punctuated Equilibrium
Prior to the 1970s most paleontologists considered a model which became known as phyletic gradualism:
Speciation is mostly sympatric: change occurs mostly within the ancestral range
Speciation is mostly anagenetic: the whole population changes together
Evolutionary change is largely continuous
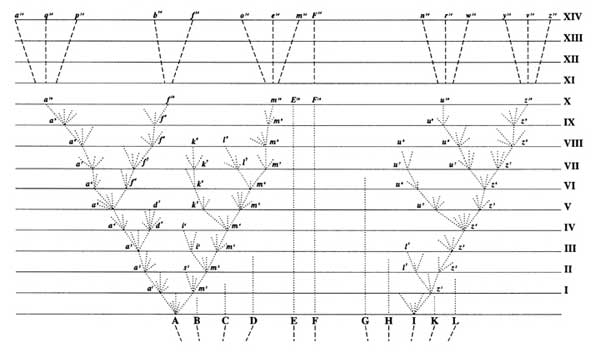
As a consequence, fossil species were all simply chronospecies: arbitrary sections of a lineage with no particular justification for dividing the lineage up one way or the other
Speciation is mostly para/peri/allopatric: change occurs mostly by peripheral isolation
Speciation is very dominantly cladogenic: branching (either subdivision of the whole population, or the origin of a peripheral isolate) is the most common
Evolutionary change is not continuous: instead, during most of the history of a species it remains relatively stable (no major directional change) (the "equilibrium" part), and most of the change is restricted to a small portion of its history (the punctuation event)
As a consequence, fossil species have a fairly distinct beginning (the punctuation)
The punctuated equilibrium model helped explain some aspects of paleontology. As Darwin noted (see quote at top), we do not see an endless series of slight gradations, each stratum with a slightly different version. Instead, species remain largely unchanged for most of their duration, with new closely-related species appearing suddenly in the fossil record. In fact, if it weren't like this, biostratigraphy would not really work! As the punctuated equilibrium supporters argued, "stasis is data".
During the 1970s and 1980s (and continuing today, but at with much less rancor), the debate over "evolution by creeps" vs. "evolution by jerks" continued. At least in the fossil record, punctuated equilibrium seems
How long are punctuation events? In a rare case, Smithsonian paleontologists Gene Hunt, Michael Bell, and Matthew Travis found that a population of the stickleback species Gasterosteus doryssus got isolated in a lake in Nevada in the Miocene Epoch. In this particular case, there were annual layers, allowing them to measure an excellent sample over time and document its change. They found the period of transition from the ancestral form to the descendent took only about 2000 generations (about 4000 years), after which the population was mostly stable. Events on the 103 year scale are unlikely to show up in the fossil record except in such situations (high sample size, restricted location, annual record), as the fossil record is much better at picking up events at the scale of 104, 105, or greater.
Supporters of the punctuated equilibrium model had to wonder how equilibrium was maintained. Evolutionary stable scenarios seem to be at least part of the reason.
Why the punctuations? A likely cause is that environmental changes are rather quick on the geologic time scale, with stable conditions in between. Rapid shifts in climates will result in shifting population ranges, shifts in habitat availability, etc. This leads to the prediction that we should see evolutionary shifts (speciations, extinctions, etc.) concentrated at moments of environment change: the so-called "Turnover-Pulse Model".
In summary, punctuated equilibrium may well be due to the following combination of aspects:
Abrupt environmental change, resulting in shifting selective conditions
Evo-devo-type evolution (see below), allowing for a few genetic changes to result in more significant changes of form
Evolutionary stable scenarios maintaining equilibrium between punctuation events
A historical note: a close read of The Origin shows that Darwin did consider cladogenesis and parapatry/peripatry as critically important in most speciation, and that anagenesis of the main part of the ancestral population was almost never the case.
"Missing Links" As An Outdated Concept
One of the most problematic terms in popular discussion of evolution is "missing link". Coined by Lyell, it was used in the 19th Century for a potential but as-of-then undiscovered link between humans and other primates. Subsequently is has been used as the idea of an as-yet undiscovered intermediate form between two particular species.
The term is problematic for a couple reasons:
Once you actually find the specimen, it isn't missing any more!
More importantly, we are unlikely to find any individual that is a direct ancestor of the later forms
The chances that any individual making it in to the fossil record is very, very small. And even the chance that every population, or even species, will show up in the fossil record is small.
An ancestor is a very specific thing: not merely an earlier relative, but specifically an individual who was in direct line of descent with the later form. (Your great great great great uncle is NOT your ancestor, while your great great grandfather is.)
So it has a very small chance that given fossils are the ancestors of the species you are interested in. However, it might well be a more general early relative, and that could be useful indeed.
Transitional Forms
Finding a direct ancestor might be hard, but finding an early relative is easier. And since relatives share many of their same traits, finding early relatives can help establish the anatomical (and ecological, and so forth) transitions involved in that part of the tree of life.
Darwin pointed out that there is not the continuous series of transitions that might expected from a gradualistic model of evolution in the fossil record, but noted that the fossil record was great for higher-level transitions. And this record is vastly better now than in the 1850s!
Here are a handful of interesting transitions recorded in the fossil record:
The various taxa in there might not be direct ancestors of the later branches, but they do show the set of traits associated with a given phase of that evolutionary transition
You don't always have to look at the whole organism: you can concentrate on the transitional states of a particular part of the body
Although not shown here, you can actually use contemporary organisms to look at the transitional phases of a particular organ system or behavior or whatever, with the understanding that you are not looking at the actual ancestors.
"Intermediate forms" is another related term used in the field. Basically, however, every taxon is intermediate between its closest relative and the groups more distantly related.
An important thing to revisit before we move on: the specter of typological thinking. Our minds like to think of discrete types of things. However, when dealing with evolution, there is a continuum of form from one to another. Remember: at no time did a mother of one species give birth to a daughter of another species! It is only from a distance in time do we see the accumulation of changes.
This applies for groups above the species as well. At not moment in the history of life would you witness a population of one major group giving rise to a population belonging to another major group. It would always look just like ordinary speciation. It is only from a distance that we see the Tree of Life.
Major Patterns of Macroevolution Macroevolution: term for evolutionary patterns at and above species level. Since "species-level" is a difficult thing to define in a consistent way that actually applies to Nature, macroevolution can be thought of as higher-level effects of evolutionary change.
With the discovery of evolution by natural selection, biologists from Darwin and Wallace's time onward have documented
many different patterns and processes in evolution. Sometimes they refer to "microevolution" (changes within an species) and
"macroevolution" (patterns on the larger scale; changes from one species to another, or between different lineages of
ancestors and descendants). It is important to remember that "micro-" vs "macro-" is just a matter of scale and perception:
at the level of individuals and populations, there is just variability, heritability, and superfecundity.
Two (or more) distinct variations in an ancestral population convey their own advantage against the rest of the population
Over time, these two (or more) variations will become more distinct from each other
If they diverge enough, they will no longer be able to mate with each other: will be different species
Divergence can also occur (perhaps more commonly!) if an ancestral population is divided into two or more by changes in geography: because natural selection works by chance survivals, it is unlikely that exactly the same variations of the ancestral population will survival in the two or more separated populations. Over time, if the populations meet again, the accumulation of variations may be significant enough that they are distinct species.
Common Ancestry
Closely related species are close because their common ancestor diverged relatively recently in Earth history
Other species are more distantly related because of divergences of THEIR common ancestors even farther back in time
No separate origins for different groups; instead, patterns of common ancestry and diverging descendants
Thus, the basic pattern of the history of living things is a Tree of Life, where the trunk and stems are lineages of ancestors, the branching points representing divergences between lineages, and the tips of the branches living species (or extinct species that died without descendants).
Other important patterns and processes:
Sexual Selection, a variation of Natural Selection recognized by Darwin, where the variation is "being more sexy" (and thus have better than average chance of breeding, and thus passing on "sexiness", compared to other members of the population [increased reproductive success]). Explains many extravagant display structures and behaviors (such as peacock tails, bird song, lion manes, etc.)
Correlated Progression: Ancestor and descendants form a lineage (historical line). Sometimes a particular life habit favors the slight increase in multiple different traits (e.g., longer and longer legs, more compact body, more efficient heart and respiration for fast running; longer and longer necks, longer legs, better cropping teeth and/or grasping tongue for browsing in trees; more and more streamlined body profile, more paddle like legs, more dorsal nostrils, etc. in swimmers; etc.) Traits that go against the general trend will be selected against; traits that go with the general trend will be selected for. (For many people, this series of trends in adaptations represents the totality of evolution)
Adaptive Radiation: If a population evolves some significant new adaptation, or colonizes a region without competitors, or is present when competitors die off, many different variations from that common ancestral population might survive (fill new or unoccupied "niches" (ways of life) in environment). Over a geologically short period time, a common ancestor can radiate into many very different descendant lineages.
Niche Partitioning: during an adaptive radiation, the early members of the divergence will (naturally) still be relatively similar to each other (and to their common ancestor) in terms of size, shape, behavior, etc. Over time, those variations in each lineage that are least like their relatives will more likely survive, because they will have less competition. Consequently, the different species will "partition" (divide up) the niches and the resources.
Convergence: Some adaptations are mechanically advantageous and easy to produce developmentally. Different lineages of organisms can independently develop some of the same features, even though ancestors were quite different (i.e., streamlining in sharks, tunas, ichthyosaurs & dolphins).
Living Fossils: Species do not have a fixed duration, but will persist until the evolve into something else and/or go extinct. In some cases, species (or genera) may persist for extremely long periods of time with no major changes.
Exaptation: Formerly called "preadaptation", the co-option of a structure that previously had some entirely different function for a new use. Seems to be the more common pattern of evolution than the appearance of entirely novel structures. For example, the wings of birds and bats were initially arms and hands; the mouthparts of various arthropods were legs; etc.
Heterochrony: Evolution by changes in rate of development from embryo to adulthood. Two major forms of heterochrony:
Paedomorphosis: descendant populations will retain some juvenile features into adulthood
Peramorphosis: descendant populations will develop structures beyond the adult form of ancestor
Extinction, Mass Extinction, & the Game of Life Extinction is the termination of a lineage. (If a species "dies out" by evolving into another species, this is more properly called a pseudoextinction). Extinctions occur throughout Earth History. What is more remarkable is mass extinction: the geologically-sudden
disappearance of many diverse groups of organisms, which are not immediately replaced by ecological equivalents. Some mass extinction events seem to correlate with major disruptions of Earth's environment.
We will explore mass extinctions in far more detail later this semester. But for now, it is worth pointing out that mass extinctions may be bad for the taxa that die out, they are great opportunity for the survivors. Certain niches (apex predators, reef makers, etc.) have been occupied by very different groups at different phases of Earth's history.
We can think of the large scale interactions within the evolving biosphere as a series of different Games of Life. Each particular Game represents some extremely broad set of interactions (rules). Introduction of a new Game into the "game room" (i.e., the totality of Earth's ecosphere) requires some major change: perhaps a new adaptation for interacting in a novel way, or the development of an entirely new way of life. In each game there are different roles: i.e., different ecological niches such as "apex predator" or "largest forest tree" or "reef framework builder." The organisms in the different roles interact with each other according to the rules of the game. But the players in each role change through time: sometimes one species may inherit its role from its direct ancestor, but sometimes the new player might come from an ancestor in a very different role. Mass extinctions represent times when many roles might be "up for grabs"; many niches are vacated, and entirely different players might take up that role. (Similarly, when new games start, it is an opportunity for players from very different ancestors to take their spot.).
The Selfish Gene: Evolution from the Gene's Eye View
When discussing evolution, we often concentrate on physical attributes (traits), or the populations, or the lineages of organisms, or even ecosystems and the like. But evolutionary biologist Richard Dawkins is right in pointing out that the only thing that is actually passed on from one generation (and thus the "material" that is being selected for or against with each selection event), is really just the gene (or coalitions of genes). By shifting emphasis from the macroscopic scale to the gene itself, natural selection be be considered to favor those traits which maximize copies of that allele into the next generation. This became the main focus of his book The Selfish Gene (which he regrets not calling "The Immortal Gene", which gets the point across better: genes out living the body of each generation.)
Selfish gene approaches -- that is, trying to understanding evolutionary events from the point of view of alleles in competition rather than bodies -- gives an important set of explanations for certain aspects of animal behavior. For example, why should animals have gregarious behaviors: that is, live together cooperatively in groups? After all, individuals within the same species have the greatest amount of overlap in requirements for resources, and would this be each others greatest competitors. So why (and when) would natural selection favor living together cooperatively? Two main reasons that--in some circumstances--cooperative group living might be favored:
Kin Selection: or, "Blood is Thicker Than Water." From a gene's perspective, protecting or nurturing close relatives with the same genes as you can be as effective in spreading additional copies of that gene. So behaviors that favor cooperation between kin might be selected for if those behaviors wind up promoting the transmission of those genes (in comparison to populations which do not have the cooperative behaviors).
Reciprocal Altruism: or, "You Watch My Back, and I'll Watch Yours." In some circumstances behaviors can be selective where individuals
keep a look out for each other (or similar type of altruism (helpfulness)) so long as they get the same benefit from other members of that population.
For example, if there are "cheaters" (ones that take advantage of others looking out for them, but that don't waste their own energy keeping a lookout)
than those populations may get additional predation (because of the predators who attacked when the cheaters could have seen them and warned others.)
If there are other populations of the same species in which the reciprocal altruistic behaviors are present, those populations will have fewer losses
and so prosper relative to the populations with cheaters.
And what about sex? Sexual strategies of the two sexes are very different: male and female animals have different priorities in terms of reproduction. Males can in principle fertilize many many individuals, while females typically have fewer sex cells (eggs) available at any given time. With less cells to use, females often are "choosier" in terms of mates. So many species evolve displays in which males somehow "show off" (in terms of physical features, ritual motions, combat between rivals, etc.) and females evaluate the display.
For example:
Evolutionary Stable Strategies (ESS): Combination of game theory and behavioral ecology. An ESS is a strategy which, if adopted by a population of players, cannot be invaded by any alternative strategy. A Nash equilibrium which is "evolutionarily" stable meaning that once it is fixed in a population, natural selection alone is sufficient to prevent alternative (mutant) strategies from successfully invading:
"A population is said to be in an evolutionarily stable state if its genetic composition is restored by selection after a disturbance, provided the disturbance is not too large. Such a population can be genetically monomorphic or polymorphic." -- Maynard Smith (1982).
Evo-Devo & Building Bodies
Since the mid-1990s, emphasis on the interrelationships between development (as studied in embryology), genetics, and whole organism biology
(esp. paleontology as record of Life's changes). Name given to this field (at a conference at the University of Maryland!): Evo-Devo (evolution & development).
It compares the developmental processes of different organisms in an attempt to determine the ancestral relationship between organisms and how developmental processes evolved. Evo-devo addresses:
The origin and evolution of embryonic development
How modifications of development and developmental processes lead to the production of novel features
The role of developmental plasticity in evolution
How ecology impacts in development and evolutionary change
The developmental basis of homoplasy and homology.
Evo-devo reflects the discovery that there are developmental genetic toolkits (such as the Hox genes of animals) that control the timing, sequence,
rate, and duration of embryological changes. Modification of these genes (first seen in homeotic mutants) can produce both minor and major
morphological variations that can be acted on by natural selection.
Evo-devo also reveals deep homologies: while some organs may be the product of convergent evolution in different lineages (classic
example is the eye) and are thus analogous structures, the tissues from which the eyes are made, and the developmental genes that control this,
are often homologous at a much more ancient level.
Many of the phenomena discussed above have been studied in fossil as well as living organisms. In fact, mass extinctions in particular are a paleontological field! We will move on to look out what insights the fossil record provides for understanding evolution.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}