

"Naturalists try to arrange the species, genera, and families in each class, on what is called the Natural System. But what is meant by this system? Some authors look at it merely as a scheme for arranging together those living objects which are most alike, and for separating those which are most unlike; or as an artificial means for enunciating, as briefly as possible, general propositions, that is, by one sentence to give the characters common, for instance, to all mammals, by another those common to all carnivora, by another those common to the dog-genus, and then by adding a single sentence, a full description is given of each kind of dog. The ingenuity and utility of this system are indisputable. But many naturalists think that something more is meant by the Natural System; they believe that it reveals the plan of the Creator; but unless it be specified whether order in time or space, or what else is meant by the plan of the Creator, it seems to me that nothing is thus added to our knowledge. Such expressions as that famous one of Linnaeus, and which we often meet with in a more or less concealed form, that the characters do not make the genus, but that the genus gives the characters, seem to imply that something more is included in our classification, than mere resemblance. I believe that something more is included; and that propinquity of descent, the only known cause of the similarity of organic beings, is the bond, hidden as it is by various degrees of modification, which is partially revealed to us by our classifications." -- Chapter 13, On the Origin of Species by Means of Natural Selection (1859), Charles Darwin
"The affinities of all the beings of the same class have sometimes been represented by a great tree. I believe this simile largely speaks the truth. The green and budding twigs may represent existing species; and those produced during each former year may represent the long succession of extinct species. At each period of growth all the growing twigs have tried to branch out on all sides, and to overtop and kill the surrounding twigs and branches, in the same manner as species and groups of species have tried to overmaster other species in the great battle for life. The limbs divided into great branches, and these into lesser and lesser branches, were themselves once, when the tree was small, budding twigs; and this connexion of the former and present buds by ramifying branches may well represent the classification of all extinct and living species in groups subordinate to groups. Of the many twigs which flourished when the tree was a mere bush, only two or three, now grown into great branches, yet survive and bear all the other branches; so with the species which lived during long-past geological periods, very few now have living and modified descendants. From the first growth of the tree, many a limb and branch has decayed and dropped off; and these lost branches of various sizes may represent those whole orders, families, and genera which have now no living representatives, and which are known to us only from having been found in a fossil state. As we here and there see a thin straggling branch springing from a fork low down in a tree, and which by some chance has been favoured and is still alive on its summit, so we occasionally see an animal like the Ornithorhynchus or Lepidosiren, which in some small degree connects by its affinities two large branches of life, and which has apparently been saved from fatal competition by having inhabited a protected station. As buds give rise by growth to fresh buds, and these, if vigorous, branch out and overtop on all sides many a feebler branch, so by generation I believe it has been with the great Tree of Life, which fills with its dead and broken branches the crust of the earth, and covers the surface with its ever branching and beautiful ramifications." -- Chapter 4, On the Origin of Species by Means of Natural Selection (1859), Charles Darwin
BIG QUESTION: How do we reconstruct how species are related to one another?
---box on typoloogical thinking & essentialism The Scala Naturae
How are organisms related to each other? That is, what is our plan of systematics (the rules for organizing taxa)? Even before the recognition of evolution (descent with modification), people ranked organisms on a Scala Naturae (scale or ladder of nature, sometimes called the "Great Chain of Being"). Humans were ranked above the animals (under the old idea that humans were not animals…), but below supernatural entities like angels and God. And within the animals, some animals were considered more "noble" or "worthy" than others.
Even after Linnaeus gave a specific way to catalog the diversity of life, people recognized "higher" and "lower" versions of animals and plants: mammals were "higher" than reptiles, which were "higher" than fish, which were "higher than invertebrates; trees were "higher" than mosses; etc.). But Linnaeus's system gave a more important aspect to compare groups: the taxonomic level at which any two species were united. So lions (Panthera leo) and jaguars (Panthera once) were united at the low level of the genus Panthera; lions and cheetahs (Acionyx jubatus) were united at the higher level of the family Feliday; lions and bears (Ursidae) at the level of the order Carnivora; and so forth.
But what did it really mean by "more closely related"? Darwin (see first quote at top) identified an answer: our classifications were pointing at the degree of recency of common ancestors ("propinquity of descent", in Victorian English). So species within a genus had only recently diverged from each other; species united at the level of a family had more distant ancestors; and so on.
Note: something that Darwin also noted, even before he wrote The Origin, is that the idea of "higher" and "lower" was an outdated concept. As all living creatures have equally ancient ancestry, nothing alive is really any more or less "high".
Consequently, THERE IS NO EVOLUTIONARY SCALE! Nothing is "higher" or "lower" on an "evolutionary scale". There IS "primitive" (closer to the ancestral condition) and "derived" (more transformed or specialized from the ancestral condition), but these are relative and specific statements. And they might apply differently to different organ systems in the same pairs of organisms. For instance, humans have vastly more derived (transformed) brains relative to star-nosed moles, but the mole's nose is vasty more advanced compared our primitive one.
Darwin observed (second quote at top) that the primary pattern to related the various species past and present is a Tree of Life:
(Darwin wrote in a notebook that a better metaphor would be the "coral of life", since anything below the tips is dead material made of rock: i.e., fossils.)
The Tree of Life would represent the basis for the higher level classifications: the farther down the tree any two branches joined up, the farther back in time was the divergence between those branches and the more distantly related they were.
This allowed a framework for a new style of systematics:
Ernst Haeckel coined the word phylogeny for a "family tree" of Life (or some subset thereof). This is a representation of an hypothesis of the actual historical pattern of ancestors and descendants. Since all living things are related (at least at some level), there should be one true historical branching Tree of Life. The task, then, is to reconstruct it.
Ideally, one could combine information from living creatures with their ancestors to complete this tree. But there are difficulties:
But how could we reconstruct the shape of the Tree of Life? Evolutionary biologists from the mid-19th Century until the later 20th Century lacked a good repeatable methodology to establish this. The most common method was simply arranging groups of living and extinct species which seemed to be closely related based on their anatomical features on a stratigraphic scale, and connecting the earlier forms with later ones on the assumption that they were the direct ancestors.
The problem, as we noted last lecture, is that being an ancestor is a very, very specific thing. Being an early relative is not enough: an ancestor is specifically a great great great… grandparent, not a cousin a few dozen times removed.
How to deal with these issues? A solution was found by East German dipterist (fly specialist) Willi Hennig in the 1950s (but not know in English until the 1960s). The technique was called phylogenetic systematics, or more commonly cladistics (from the Greek "klados" ["branch"] for clade, meaning a branch of the Tree of Life). Hennig recognized that finding direct ancestors in the fossil record would be hard, and demonstrating that the fossil was a direct ancestor rather than a close relative of that ancestor would be even harder. So instead, his method focused on estimating the pattern of shared common ancestry. In order to make that estimation, cladistics looks at the distribution of derived characters [evolutionary specializations]. Here is how it works. Below is a grouping of some modern animals:
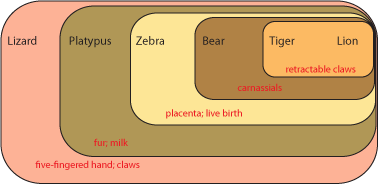
Each set contains multiple taxa. Each set is characterized by derived characters not seen in the taxa outside that set. For example, the lizard shares with all the others the presence of a five-fingered hand and claws. The platypus and all the others except the lizard shares fur and milk. Since lizards (and alligators and frogs and fish and many other animals not shown here) do not have fur and milk, it is most likely explained by having evolved in the common ancestor of platypus, zebra, bear, tiger, and lion AFTER that ancestor had diverged from the common ancestor of lizard, platypus, zebra, bear, tiger, and lion. In other words, platypus, zebra, bear, tiger, and lion most likely had a more recent common ancestor with each other than any did with lizards.
It is generally easier to show this pattern using a simplified stick figure called a cladogram than the set diagram. Below is the cladogram showing the same information as the sets above:
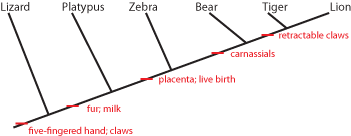
The little red "tick marks" show that the derived character state appears at that point of the cladogram, and is passed on upwards. To show what that means, see the following:

So the common ancestor of zebra, bear, tiger, and lion evolved a placenta and live birth, and passed it on to its descendants. And the common ancestor of bear, tiger, and lion (but NOT zebra) had its molars evolve into carnassials (shearing teeth), and passed that trait on.
Not all character states are useful in figuring out evolutionary relationships. Primitive (or ancestral) states do not help. For example, both lizard and platypus lay shelled eggs rather than have a placenta and live birth. Does that mean that these two animals shared a more recent common ancestor with each other than either did with the other animals here? NO! In fact, having a shelled egg is the ancestral state for a larger (more inclusive) group of animals, including birds, crocodilians, turtles, snakes, echidnas, etc. Having a shelled egg is simply the condition (state) of all advanced terrestrial vertebrates. The technical term for a shared primitive character is a symplesiomorphy.
However, fur and milk are derived states not found in terrestrial vertebrates other than mammals, and indicate a common ancestor to these animals not found in lizards, birds, crocodilians, turtles, etc. The technical term for a shared derived character is a synapomorphy
Similarly, sharing a five-fingered hand doesn't unite the lizard, platypus, and bear (for example) exclusive of the zebra. A five-fingered hand is a symplesiomorphy, and will be passed on that way unless it evolves into something else.
(NOTE: a character can be "primitive" at one level, but "derived" in a different context. In this cladogram, retractable claws is a derived feature that shows us that lions and tigers share a more recent common ancestor with each other than with anything else on this cladogram. However, if we were trying to figure out the relationship between lions, tigers, jaguars, and leopards, than retractable claws would be a primitive charcter shared by all of them, and so useless.)
Zebras have only a single finger on each hand. This is an autapomorphy or unique derived character. Although autapomorphies might be useful in understanding how that animal lived, they don't help us figure out where they fit in the cladogram, since those traits evolved after the ancestor of (in this case) zebras diverged from any shared ancestor of any of the other animals on the list. (However, if we added horses or donkeys to this cladogram, a single finger on each hand would be a good shared DERIVED character, and help us figure out that horses and donkeys share a common ancestor not shared with bears, lions, tigers, etc.).
Convergent characters could actually mislead us. For example, tigers and zebras both have stripes, whereas the other animals on this list don't. Does that mean that tigers and zebras share a common ancestor not shared by the other animals? We might think so at first, but the shared presence of carnassials in bears, lions, and tigers (but not zebras) and of retractable claws in lions and tigers (but not zebras) indicate a different pattern of ancestry. Convergent characters can be very annoying, because they might cause us to make the wrong estimation of the cladogram.
An additional type of character (not seen here) are reversals: when an ancestor had a derived condition, but the trait evolved "back" to the primitive condition. For example, snakes are descendants of typical legged lizards. But snakes have evolved the loss of limbs, so that they resemble the limbless ancestor of vertebrates. For another, while the common ancestor of living birds could fly, but ostriches, rheas, cassowaries, etc. have reverted to the primitive flightless state. Reversals that are unique characters do not help in estimating evolutionary relationships, but ones which are shared might. In general, though, reversals (like convergent characters) are annoying and are a major source of error for our analyses. Collectively, we call convergent characters and reversals homoplasies.
So, in review:
How do we tell which states (conditions) of a character are ancestral, and which are derived? One common method is to look at outgroups: more distant relatives of the taxa in question. Looking at fish, amphibians, turtles, lizards and snakes, crocodilians, and birds show that no placenta is the primitive condition among vertebrates, and that the placenta of some mammals is a derived condition. (It is best to look at multiple outgroups.)
We have a set of taxa for whom we want to understand the evolutionary relationships: the ingroup. They will vary from each other on different features. The question is: which features represent symplesiomorphies (shared primitive characters inherited from their distant ancestors) and which are synapomorphies (shared derived characters). In order to do this, we need some data to figure out what the ancestral condition is.
Since finding actual ancestors is very difficult, we need a stand in for the ancestor. It should be a close relative of the ingroup (based on some previous studies), but outside of the ingroup. This is called the outgroup. (In more complete analyses, we would use multiple outgroups and additional evidence (developmental sequences, for instance) to estimate what the ancestral condition was.
We then can map out the distribution of traits in each of the possible cladogramss. For each, we count the minimum number of state changes from primitive to derived (or from one derived to another derived, or even from derived to primitive) condition to explain the observed observations for that tree. The total number of these state changes for each possible tree are compared to all the others; the one (or, more often, ones) with the fewest number of required changes is/are called the most parsimonious tree(s) or preferred tree(s).
Of course, this may not be the true tree at all! But it IS the best solution for the observations concerned. Additional observations and reanalysis with new characters, new taxa, or both could result in a different tree shape (tree topology) being preferred.
Relationships, Not Similarity: Darwin, Hennig, and their followers advocate the use of common ancestry, rather than overall similarity, as the basis for classification. The reason? Overall similarity might represent common ancestry if the organisms are similar because of shared derived characters. But overall similarity might also simply represent the shared inheritance of primitive traits.
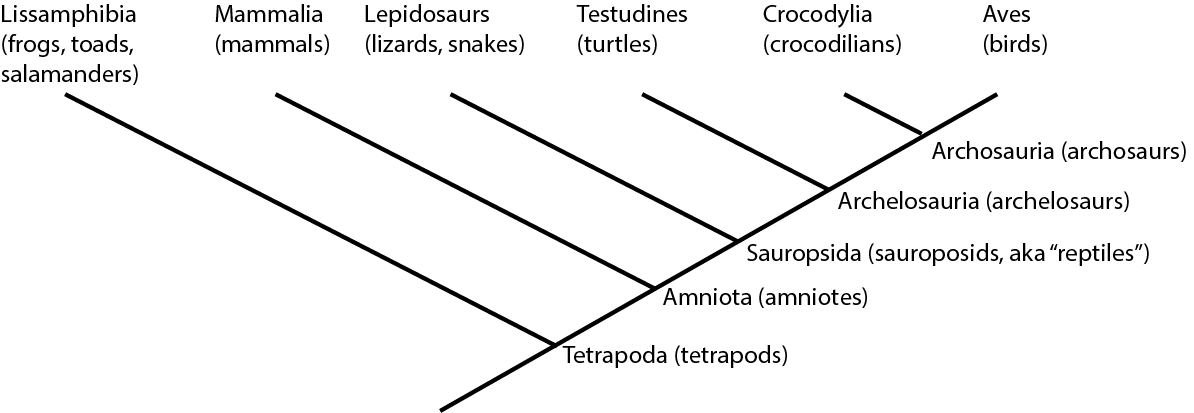
For example, below is the cladogram for the living members of Tetrapoda, the clade (group) of terrestrial vertebrates:
On a cladogram, a name refers to the entire clade: an ancestor plus ALL of its descendants. (Another name for
"clade" is "monophyletic group" [monophyly = one branch].)
We could also write out this information like an outline: Tetrapoda
So:
Traditionally, crocodilians would be thought to be "more closely related" to lepidosaurs and turtles than to birds, because of the overall similarity. But this overall similarity was simply shared primitive features. Instead, as we will see, crocodilians and birds actually share far more derived features with each other than either does with lepidosaurs or turtles. Thus, birds and crocodilians are more closely related to each other than either are to lepidosaurs or turtles. (To use the technical term, Aves and Crocodylia are each other's sister group.):

In fact, lepidosaurs are no more closely related to crocodylians than they are to birds! They simply resemble crocodilians more because the bird lineage has gone through many more spectacular evolutionary transformations!
Note that the traditional version of "Reptilia" (= Lepidosauria, Testudines, Crocodylia, but not Aves) is NOT a monophyletic group. Instead, it is what is called a paraphyletic group: an ancestor, but NOT all of its descendants. You can see the distinction here:
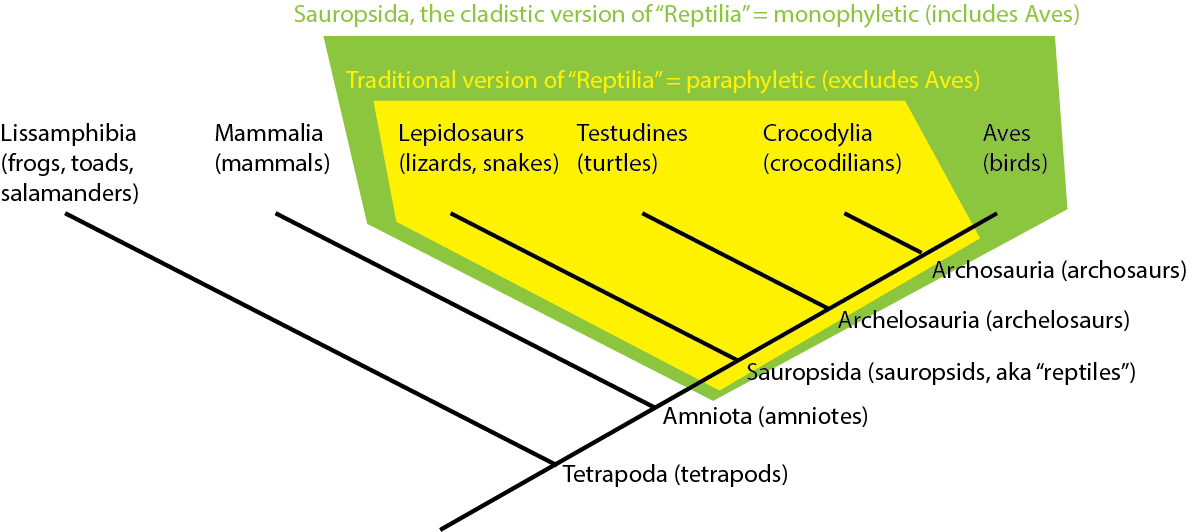
Many traditional Linnean groups have turned out to be paraphyletic groupings (Pisces; Invertebrata; Fissipedia; etc.). However, because cladistic taxonomy uses only clades (monophyletic groups), we no longer use these paraphyletic taxa.
Sister Groups, Not Ancestors: One important aspect of cladistics is that our analyses are designed to find the most likely sister groups for given taxa, not their actual ancestors. So in the case above, Mammalia is not the ancestor to Sauropsida or vice versa. They are simply each other's closest relatives. However, as we shall see, we can use cladistic analyses to estimate what the ancestral condition most likely was.
Names on the Tree: Higher-Level Taxonomy: Paleontologists and other biologists have started to use phylogenetic definitions for taxa: that is, they specify a particular relationship and any taxon that qualifies under that relationship is member of that clade. Biologists have coined the word concestor for most recent common ancestor. While the chances of finding the concestor of any two (or more) clades is slim, we can recognize that there did exist such a species somewhere in time. This recognition gives us a way of defining taxa.
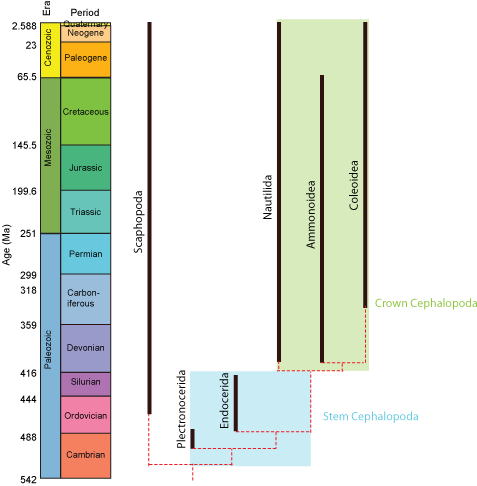
Paleontologists sometimes find it useful to refer to the crown-group: that is, the clade formed by all descendants of concestor of the living members of a clade. The total-group is the clade comprised of the crown-group plus all taxa sharing a more recent common ancestor with it than with the next most closely related crown-group. Finally, the stem-group is the paraphyletic part of the total group excluding the crown group. In the example below, the total-group Cephalopoda includes the modern Nautilida and Coleoidea and all taxa more closely related to them than to Scaphopoda. Nautilida and Coleoidea define the crown-group, which also contains the extinct clade Ammonoidea (which although extinct is nevertheless a descendant of the concestor of nautilids and coleoids.) Plectronocerida and Endocerida, however, belong to the stem-group of cephalopods.
However, we can go beyond simply talking about crown- and stem-groups. For example, Amniota is defined as "the concestor of Mammalia and Reptilia, and all of its descendants." And Dinosauria is "the concestor of Iguanodon and Megalosaurus, and all of its descendants.". So any taxon that comes from the node (joining point) that links Megalosaurus and Iguanodon is a member of Dinosauria (i.e., is a dinosaur); and anything that lies outside that node is NOT a dinosaur. So in the cladogram below:
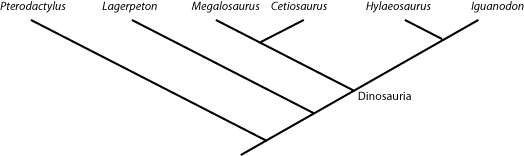
Cetiosaurus, Hylaeosaurus (and, by definition, Megalosaurus and Iguanodon) are all dinosaurs, but Lagerpeton and Pterodactylus are not.
Names coined in this fashion are said to have node-based taxonomic definitions. These have the form "the concestor of A and B and all of its descendants" or more generally "the least inclusive clade containing A and B". (A and B are said to be the specifiers or anchor taxa.)
Names can also be be defined another way: by sister group relationship rather than by concestry. For example, the two major branches of Dinosauria are Saurischia and Ornithischia. The former is defined as "Megalosaurus and all taxa sharing a more recent common ancestor with Megalosaurus than with Iguanodon", and the latter is defined as "Iguanodon and all taxa sharing a more recent common ancestor with Iguanodon than with Megalosaurus". So Cetiosaurus and Megalosaurus are both saurischians, and Hylaeosaurus and Iguanodon are both ornithischians. These names have branch-based taxonomic definitions. These have the form "A and all taxa sharing a more recent common ancestry with A than with B" or more generally "the most inclusive clade containing A but not B".
Similarly, the clade Dinosauromorpha is defined as Dinosauria and all taxa sharing a more recent common ancestor with
Dinosauria than with Pterodactylus. Showing those relationships (by labeling the name, and by color blobs) onto the
cladogram, we see:

In principle, one can also coin apomorphy-based definitions. These have the form "The clade stemming from the first organism or species to possess apomorphy M as inherited by A" or "the most inclusive clade exhibiting character state M that is synapomorphic with that in A". For example, in the diagram above, Hylaeosaurus is unique in possessing osteoderms (armored bony plates in its skin). One could define a clade "Hylaeosauroidea" with the form "The clade stemming from the first species possessing osteoderms as inherited by Hylaeosaurus". However, most practitioners of phylogenetic systems of nomenclature avoid this, as apomorphies almost always exhibit transitional stages, which makes the dividing point operationally difficult to define.
Inferring Missing Information:
No fossil is complete, so we can't always directly tell what character state was present in a given species. But we can sometimes us the phylogenetic position of a fossil to help us estimate what that state was. Below is a cladogram of tyrannosauroids (tyrant dinosaurs), with the number of fingers on the hand listed:
:
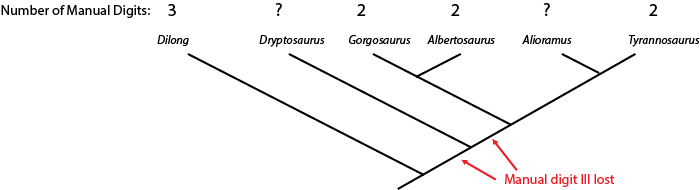
The hand is not yet known for Alioramus or Alectrosaurus. However, we know that Dilong has the ancestral condition of a three fingered manus (shared with many other carnivorous dinosaur groups), and that Gorgosaurus, Albertosaurus, and Tyrannosaurus all share the evolutionary specialization "loss of manual digit III" (meaning that they had a two-fingered hand). The simplest explanation is that the concestor of these three dinosaurs had already lost manual digit III, and passed on this derived trait to the different descendant lineages.
Because Alioramus (based on other evidence) is ALSO a descendant of that concestor, we predict that it too had a two fingered hand. To assume it had a three fingered hand would mean assuming an evolutionary change (in this case, a reversal) for which we had no direct evidence. The simplest explanation for our current observations is that Alioramus was a two-fingered dinosaur.
But things are unclear for Alectrosaurus. It might be that the loss of manual digit III occured after the ancestor of Alectrosaurus diverged from the concestor of Alberosaurus through Tyrannosaurus: if that were the case, Alectrosaurus would have a three fingered hand. OR it might be that the loss of manual digit III happened before the Alectrosaurus lineage and the Albertosaurus + Tyrannosaurus lineage had diverged: in that case, Alectrosaurus was a two fingered dinosaur. So at present, things remain ambiguous for this taxon.
For any fossil form, we can find the two closest outgroups which are still extant. These represent the extant phylogenetic bracket (or EPB) of the fossil taxon. If we hypothesize that the extinct form has a particular soft tissue structure, behavior, or other character that cannot be directly observed from the fossil record, this inference falls in three categories of decreasing confidence:
Below are examples of each of these:

In the first case, the basal horse Eohippus is bracketed by Ceratomorpha (rhinos plus tapirs) and Equus (the living horse genus). Both of the EPB possess fur, so the inference that Eohippus was furry is a secure Type I inference.
In the second case, the therapsid "protomammal" Thrinaxodon is bracketed by living mammals (which possess fur) and living reptiles (which do not). We might infer that Thrinaxodon was furry, but this is a less-secure Type II inference. Of course, preservation in a Konservat-Lagerstatt might someday show that Thrinaxodon had fur (direct fossil evidence) OR there might be a similar discovery of fur in a taxon more distantly related to mammals than Thrinaxodon (which would make the furry Thrinaxodon hypothesis a Type I inference, with an extinct but well-preseved taxon sitting in as part of the bracket).
In the final case, some paleontologists have hypothesized that the bird-hipped herbivorous dinosaurs in Ornithischia had mammal-like cheeks. While this is argued on circumstantial evidence, it represents the least-secure Type III inference using the EPB method, as neither members of the EPB (crocodylians and birds) have cheeks.
Inferring Ancestral Status: Looking at the above situations, we used the observed pattern of character states to infer the most likely status of that given character at a particular node (for instance, in the tyrannosaurid case above, the status of a two-fingered hand at the concestor of Tyrannosaurus and Albertosaurus, but of a three-fingered hand at the concestor of Tyrannosaurus and Dilong.) By this method, we can predict the set of character states for the various characters present at any given node of the tree. (Of course, because of missing data or homoplasy, some of those statements will be uncertain.)
The resulting set of character statements is a prediction of the condition of that concestor. Like any hypothesis these will be modified with new information; nevertheless, it represents our best way at present to estimate the status of any specified ancestor.
Of a particular note, we'll find that the concestor of the crown-group will have the basal (ancestral) condition for the living members of that clade, but that the various more distantly related members of the stem will have progressively fewer and fewer number of the synapomorphies of the crown. In fact, the very first taxon of the stem-group would have almost none (or even possibly no) synapomorphy with the crown group that is not also present at the node uniting the crown group and its closest living crown group. After all, when it lived, there was no way of knowing that the brand-new subdivision of the population (which after time gave rise to our crown group of interest) was in any way distinct from all other subdivisions of the ancestral species. It is only the accumulation of new traits over long time that make the new lineage distinctive.
Divergence Times
On a cladogram we don't normally see numerical time. Because the nodes on a cladogram represent the presence
of shared common ancestors, they are guide to the relative sequence of divergences. So in the cladogram of
feathered dinosaurs to the left below:
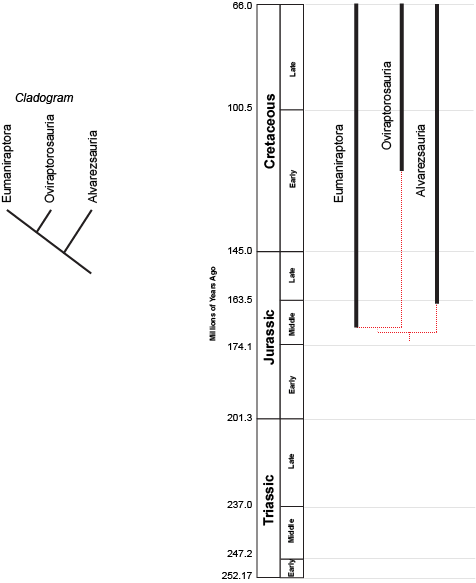
the concestor of Eumaniraptora + Oviraptorosauria must have lived more recently (closer to us in time) than the concestor of (Eumaniraptora + Oviraptorosauria) + Alvarezsauria. (After all, the latter concestor is the ANCESTOR of the (Eumaniraptora + Oviraptorosauria) concestor!) But we can't tell numerical time from a cladogram.
However, we can plot the known stratigraphic ranges (that is, the range between the oldest and youngest specimen) of each taxon. Those are the black lines on the phylogeny to the right above. The dashed red lines represent the cladistic relationships between the taxa. We see that the divergence between Eumaniraptora + Oviraptorosauria had to have happened sometime before the oldest member of these sister taxa (in this case, late Middle Jurassic eumaniraptorans). The divergence between Alvarezsauria and (Eumaniraptora + Oviraptorosauria) had to happen earlier than that. We don't know the exact divergence date unless we actually find the concestors; but we can find the minimum divergence date (in both these cases, the Middle Jurassic).
The phylogeny also shows us other information. For instance we see that Oviraptorosauria is first known from the later part of the Early Cretaceous.
We can also see that there are still plenty of dinosaurs to find! Assuming our cladogram is correct, there should be Middle Jurassic, Late Jurassic, and early Early Cretaceous representatives of the Oviraptorosauria lineage (these wouldn't be any of the currently known members of Oviraptorosauria, but dinosaurs sharing a more recent common ancestor with them than with eumaniraptorans).
Also, The Tree Room has a great set of explanatory webpages about understanding cladograms.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}