


"It is clear, therefore, that we are now in an altogether exceptional period of the earth's history. We live in a zoologically impoverished world, from which all the hugest, and fiercest, and strangest forms have recently disappeared; and it is, no doubt, a much better world for us now they have gone. Yet it is surely a marvellous fact, and one that has hardly been sufficiently dwelt upon, this sudden dying out of so many large mammalia, not in one place only but over half the land surface of the globe." - The Geographical Distribution of Animals, Alfred Russel Wallace (1876)
and
"On one side are those who argue for a climatic cause, while on the other are those who hold human hunting to be responsible. I subscribe to the black hole theory of extinction, which suggests that at the end of the Pleistocene all of those huge mammoths, sloths, camels and mastodons disappeared into a black hole. Not any black hole, mind you, but that very discerning one lying between nose and chin on the Clovis physiognomy." - The Eternal Frontier, Tim Flannery (2001)
BIG QUESTION: What happened to the Pleistocene megafauna?
The Pleistocene Megafaunal Extinctions
Ice Age Mammals and the Antiquity of Humans
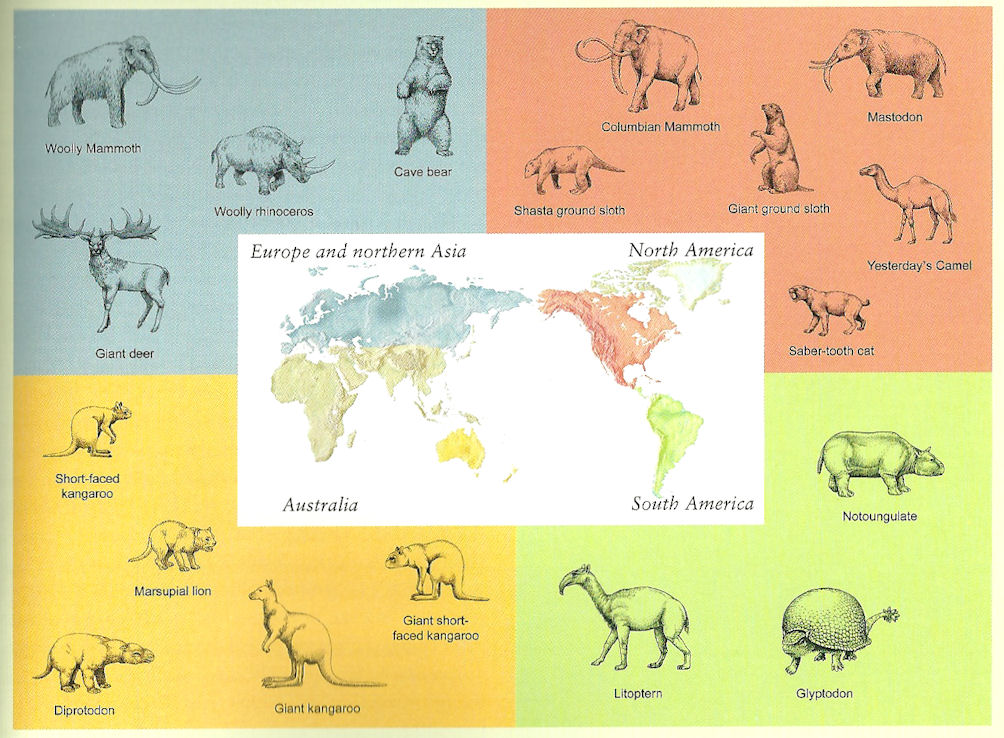
Some of the first fossils found and named by paleontologists were giant mammals and birds from the very youngest geologic past: Mammuthus (the various mammoth species); Mammut (the mastodon); Coelodonta (the woolly rhino); Megatherium and other giant ground sloths; Diprotodon, a giant wombat of Australia; Toxodon, a giant placental mammal of South America; and so on. Early naturalists such as Louis Agassiz and Alfred Russel Wallace thought that these species were wiped out by the beginning of the Ice Age.
But as the fossil record improved, it was discovered that rather than a single "Ice Age" there were multiple advances and retreats of the ice, and these giant animals lived throughout this interval. Furthermore, Charles Lyell and others established that humans co-existed (and even hunted) these animals. Indeed, we have paintings from humans who witnessed these animals in life.
So if the Ice Age itself didn't wipe out these large animals (megafauna), than what did?
Roll Call of Doom
Africa: Essentially modern composition of the animal life in the Pleistocene (indeed, nearly modern in the Pliocene). So very little evidence for a Quaternary Period extinction among carnivores and herbivores from the continent of humanity's birth.
Indian Subcontinent: In work first published in 2020, the pattern for this region is somewhat similar to Africa. Many of the species present in the Pleistocene are still with us today. The extinction includes two proboscideans: the giant straight-tusked elephant Palaeoloxodon namadicus (possibly the largest land mammal of all time) and smaller Stegodon namadicus, an extinct species of the pygmy hippopotamus genus Hexaprotodon, and the local horse Equus namadicus (as well as regional extinction of the ostrich Struthio camelus and the pseudoextinction of the Indian aurochs Bos namadicus, ancestor of Indian zebu cattle).
East Asia, Southeast Asia, and Indonesia: Less well studied than many other regions, there were local mammoths and other proboscideans (such as species of Stegodon), rhinos, and deer which died out, as well as the giant orangutan relative Gigantopithecus and the tiny hobbit Homo floresiensis. (Indeed, on Flores Island there were giant storks, giant rats, giant lizards, and dwarf Stegodon: an odd community). Also, the "mystery" Denisovans and Maludong peoples disappear as well.
Boreal Eurasia: In addition to Mammuthus primigenius (the woolly mammoth: also in North America) and Coelodonta antiquitatus (the woolly rhino), there was the giant Irish elk (Megaloceros giganteus), the cave lion (Panthera spelea), the cave bear (Ursus speleus), and Homo neanderthalensis. (Review the Eurasian Ice Age lecture.)
Beringia: Many fauna shared in common between Boreal Eurasia and Continental North America.
Continental North America: Prior to the megafaunal extinctions, continental North America south of the Ice was one of the most diverse assemblages of large bodied mammals the world has ever seen; rivaling or surpassing the modern Africa savannas. Among the notable victims were the giant short-faced bear Arctodus simus, the giant American lion (once thought to be a jaguar-like cat, but now found to be the sister taxon to the cave lion and close to the extant lion) Panthera leo atrox, the dire wolf Canis dirus, the North American sabretooth cat Smilodon fatalis, and the giant condor-like Teratornis merriami among carnivores; and the extremely tall Columbian plains mammoth Mammuthus columbi, the more woodland American mastodon Mammut americanum, the giant black bear-sized beaver Castoroides ohoiensis, Yesterday's camel Camelops hesternus, the stagmoose Cervalces scotti, the huge broadheaded bison Bison latifrons, several species of native horses (including local populations of Equus ferus, the same species that in Eurasia would be domesticated to become the domestic horse), various species of ground sloth such as elephant-sized Eremotherium laurillardi, rhino-sized Megalonyx jeffersoni, and bear-sized Nothrotheriops shastensis, and the giant super-armadillo glyptodont Glyptotherium arizonae among the herbivores. The sloths and glyptodonts were recent immigrants from South America.
South America: For much of the Cenozoic, South America was an island continent with a unique fauna of marsupials, endemic placentals, giant birds, and so forth, but during the Neogene an interchange of faunas with North America over the young Isthmus of Panama mixed the assemblages. Some South American animals did well in the North: opossums, armadillos, porcupines, ground sloths, glyptodonts. But far more North American animals did well in the South: proboscideans, horses, camels, dogs, cats, rodents, raccoon-relatives, peccaries, tapirs, deer, squirrels, rabbits, and so on. There was a major extinction of the native South American animals at this time. But some of the native forms and new immigrants thrived and diversified up to the Pleistocene, including the largest gylptodonts such as Doedicurus clavicaudatus, the largest ground sloths such as Megatherium americanum and nearly as large Eremotherium laurillardi (which ranged into North America as well), as well as a diversity of smaller sloths, camel-like Macrauchenia patachonica and rhino-like Toxodon platensis (last survivors of once-diverse radiations of South American hoofed mammals), the sabrecat Smilodon populator, various elephants such as Cuvieronius hyodon, Stegomastodon platensis and S. waringi (both genera were also present in North America earlier), a local radiation of horses, as well as other diverse forms.
Sahul: In addition to the giant wombat Diprotodon opatum, other notable marsupials which died out in the late Pleistocene were the marsupial "lion" (probably more like a jaguar ecologically) Thylacoleo carnifex and the giant kangaroos Procoptodon goliah and Simosthenurus occidentalis. But in the Sahul some giant sauropsids were present as part of the megafauna: the possibly-carnivorous giant duck relative Genyornis newtoni; a giant constricting snake Wonambi naracoortensis (not a boa or python, but last survivor of a Cretaceous lineage of big snakes); the giant club-tailed horned turtle Meiolania platyceps; and enormous Varanus priscus (once called "Megalania"), a huge predatory lizard closely related to the Komodo dragon (Varanus komodoensis).
What sort of patterns do we see in the extinction event? In particular, are there common ecological factors? Common factors of intensity? Common factors of timing?
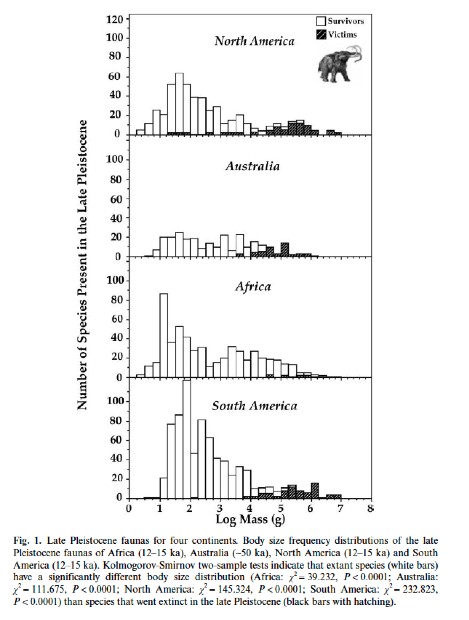
Size Pattern:Overall, there is an important thread to all these extinctions: they strongly bias towards size. That is to say, large animals tend to die out, small ones tend to survive. In contrast, the general ecology of the victims (carnivores vs. herbivores; browsers vs. grazers; woodlands vs. grasslands; uplands vs. lowlands; etc.) seem to be far less important. Hence, this has been called the Pleistocene megafaunal extinction as it really only affects the big land animals.
NOTE: Properly speaking, this is NOT a "mass extinction" as that term is used by paleontologists who work in the rest of the geologic record. We see no impact on the marine invertebrate fauna (which are the key aspects of ALL true mass extinctions), nor plants, nor smaller terrestrial fauna. It is limited to larger-bodied land mammals and reptiles (including birds). That is not so say there are no extinctions among smaller land animals; however, these do not show elevated rates above background (as opposed to the massive die off of large land beasts).
Regional Patterns: Why No African Megafaunal Extinction?: If we look at the differences in regional intensity of extinction, we notice a pattern among land mammals greater than 40 kg (88 lbs) adult body mass:
Why this pattern? Africa essentially doesn't participate in the extinction (it is essentially background rates), India very low rates, Europe has moderate rates, the Americas very high rates, and the Sahul catastrophic ones.
It is not a difference in environment: much of Africa is similar to either the Sahul or South America in terms of environment, while Europe and North America have largely similar environments.
The pattern does match a particular aspect, though: regions in which no homininan (or indeed, no hominoid!) had ever lived prior to the arrival of H. sapiens saw truly tremendous extinctions; Europe (and Asia, not shown)--where earlier species of Homo had lived by and hunted the local megafauna for hundreds of thousands of years--shows intermediate levels; and Africa--in which Hominina, and indeed Homo sapiens, first evolved and spent most of its/our evolutionary history--shows essentially none. The megafauna of Africa co-evolved with us, and so had much longer time to adapt to our escalating ability to hunt; the Eurasian forms had at least some experience with human hunters of some species; but the Brave New Worlds first encounters were with fully modern humans.
Timing?: This remains a contentious issue. As we will see there are a handful of late survivors of some species in isolated environments, so what do we mean when we refer to the timing of the extinction? Furthermore, not all the taxa die simultaneously on the scale of centuries.
That said, the signal is very clear on a broad scale: Australian forms undergo the strongest intensity of extinction in the 45-40 ka range; Boreal Eurasian forms tend to die out in the 30-25 ka range (with some late survivors); American ones in the 13-11.5 ka range. Intriguingly, these match pretty closely the arrival time of H. sapiens (or at least continued presence of them) to these respective regions.
An interesting test of the megafaunal extinction is the regional loss or decline of the fungus Sporormiella in the fossil record. Sporormiella flourishes in the dung of megafauna, so the decline and loss of its spores is a good sign that the biomass of big animals has crashed. It disappears all across North America at 12.6 ka, for instance, but returns around 10 ka when the modern bison species begins to flourish.
So what are the potential triggers for the megafaunal extinction? Four main contenders exist:
Let's take a look at these.
Although climate might be a trigger, it is unlikely to have been the actual killing mechanism. Nevertheless, climate change can greatly change the distribution of plants, their growing seasons, habitat ranges, and so forth. From modern experiences we recognize that habitat disruption is one of the primary factors in contributing to the endangerment and extinction of animals: with their home ranges broken up, each reduced subpopulation becomes much more vulnerable to extirpation (local extinction) from other factors.
Computer modeling does indeed show that during the Boreal Eurasian extinctions the northward migration of the cooler zones would greatly reduce the habitat of the "woollies" (mammoths, rhinos) and their predators. On the other hand, southern Eurasian megafauna (many of which also died out) would see their ranges increase. It is not certain to what degree these changes would have impacted on the fauna of the Americas and Australia, however.
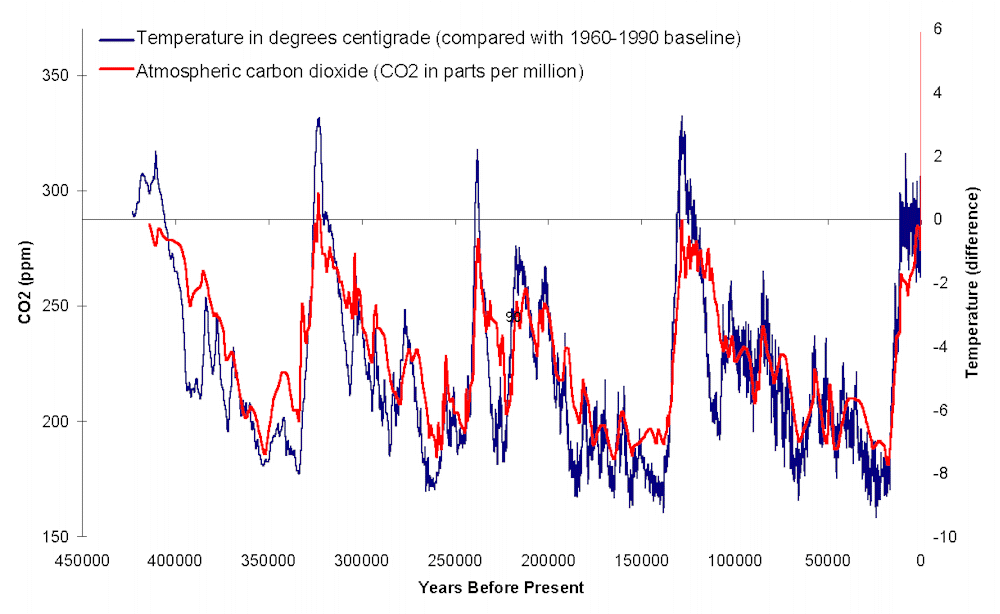
The Turnover Pulse model of evolutionary change would suggest we would see extinctions rather simultaneously around the world, since most climate shifts are global. But that is not the pattern that we see. If we look a the temperature curve for the Pleistocene and Holocene, we find that Australian and Eurasian extinctions are during the middle of a cooler phase, while the American ones do indeed fall during a dramatic glacial-interglacial transition.
But it is critical to note that nearly all these species survived climate transitions as strong or stronger earlier in their history. Most of these species were present beyond the scale of this graph, yet made it through every earlier warming or cooling phase. The most recent deglaciation is NOT the most extreme of the late Quaternary (or at least, not until the 20th and 21st Centuries, long after these extinctions!). So why didn't they die out in these earlier transitions?
Historically there have been cases where mass death of animals occurred. (For example, in the 1890s wild African antelope species suffered from the Eurasian rinderpest disease, brought in via infected cattle; or the 2015 Siberian plague of Pastuerella multocida which killed 70% of the saiga antelopes) No one has seen a widespread species wiped out by such a disease, but in principle such an event might be possible.
So might these extinctions be cases of hyperdisease? It is very unlikely that any one single disease could cause this scale extinction, but conceivably migrations of multiple species from one region to another might bring multiple diseases.
However, for the Boreal Eurasian and Australian events there does not seem to be immigration of fauna (other than humans) at the time of the extinctions. It is true that in the Americas there are "fellow travelers" (as we shall see) that might possibly have been vectors. But overall the phylogenetic and environmental diversity of the victims makes it unlikely that disease organisms were the primary culprits.
In 2007 R.B. Firestone and colleagues proposed that a cometary impact over North America around 12.9 ka was responsible for the North American megafaunal extinction, the end of the Clovis culture, and the trigger for the short-term Younger Dryas cooling event. This was claimed on the basis of supposed extraterrestrial material of various sorts (microdiamonds, iridium; possible tektites; fullerenes, etc.), as well as soot and charcoal, in unusual black carbon-rich layers at that time. While capturing the public imagination, many of the results of Firestone's lab have not been duplicated by other researchers. (And indeed, some have proposed that the initial discoveries included at least some contamination within the labs.)
But even if true, this cometary impact does not coincide at all with the Eurasian or the Australian extinction event.
So what about humans? The timing and the intensity of the extinction events seems to coincide extraordinarily well with arrival of modern humans into the respective regions. Additionally, the fact that the extinctions are limited to larger-bodied animals (either the likely targets of direct hunting, potential rivals, or animals whose resources were robbed as they became extinct) very much matches the expectations of encounters with foraging human tribes (who would be unlikely to effect the extinction of common small-bodied animals or marine invertebrates). But could humans have caused this scale extinction? That is, could we be "Homo destructor"?
Animals in environments absent of humans (such as the Galápagos Islands off of Ecuador) are naive; they do not represent humans as threats. So it is possible to approach them more closely than wild animals which have evolved around the presence of humans. So the Brave New Worlds fauna in particular might be more vulnerable than Eurasian and African megafauna.
Might there really have been what Paul Martin calls "Pleistocene Overkill" or "Blitzkrieg" in North America (and equivalent in Australia): that is, the 1000-year or so long killing spree? As it turns out, computer models show that at reasonable rates of hunting success, human population growth, and reproduction rates of prey, you could in fact wipe out continents' worth of animals in a millennium or two! And given that in the New World the Clovis culture range is basically of just this length, it appears that this might have been the case.
Some animals (mammoths, mastodons, horses, etc.) show signs of direct hunting. But would humans have hunted the predators to death? While it is not improbable that humans hunted them directly on occassion, it is more likely that sabrecats, dire wolves, and the like underwent predator crash: humans overhunted the animals the predators were better adapted to eating.
Some survivors were able to undergo a trophic shift: for instance, the California condor (Gymnogyps californianus) initially fed on large land mammals all over the American West as well as beached whales and seals on the coastline during the Pleistocene, but with the megafaunal extinction survived only on the coast. In fact, when first recorded by Spanish and English-speaking colonists, it was a shoreline bird. But the rise of the American ranchlands and the regional extirpation of whales and seals due to over-harvesting in the mid-late 19th Century drove the condors inland. (In the 20th Century poaching, poisoning, and habitat destruction drove the species to extinction in the wild, but it was saved in captivity and is being reintroduced to the West.)
Other predators survived by being generalists. While Smilodon and Panthera atrox were big-game specialists, the smaller P. onca (jaguar) and Puma concolor (puma, cougar, or mountain lion: they are all the same cat!) are generalists. Jaguars hunt everything from land animals to fish and turtles, and pumas have a similarly broad diet and an amazing habitat tolerance (from the high mountains to woodlands to plains to forests to rain forests). A tooth-wear analysis published in April 2014 showed that the pumas had a far broader diet than the sabrecats and American lions in the same localities: the ability to feed on a greater variety of foods meant the smaller cats could thrive when the menu got reduced.
Fellow Travelers: If you look at the list of the largest native North American mammals today, it turns out four of the most common (the moose Alces alces, the elk Cervus elaphus, the brown bear Ursus arctos, and the wolf Canis lupus) are all very recent immigrants. Indeed, they may have arrived in North America with the Paleo-indians! These species also live in Eurasia, and so are much better adapted to living alongside humans and our effects.
The largest contemporary North American mammal, the plains bison Bison bison, is actually YOUNGER than the Paleo-indian arrival. Descended from the older native B. antiquus by way of an intermediate species B. occidentalis, it originated around 10 ka. Even the oldest plains bison show signs of hunting (Folsom points). Bison in general have faster-than-expected rates of reproduction for big animals (only 217 days, compared to 335 for horses, 380 for camels, 450 for rhinos, and >600 for elephants) and thus were able to adapt to change faster than other large animals. Indeed, we see that the plains bison evolves their characteristic enormous herds soon afterwards, possibly in response to pack hunting by the newly arrived Paleo-indians and wolves.
Mammoth Steppes: Lost Biomes of the Pleistocene: In many of the places in world where tundra grows today, there were very different conditions during the Pleistocene. This was a very northern grassland/meadow environment called the mammoth steppe. In the mammoth steppes, there were far more plant species (including grasses) than in the tundra; there were very deep soils; permafrost was quite far below the surface, so the soils were better drained; and they supported much bigger biomass. Mammoths were not merely inhabitants of these steppes; they were likely the architects of them. Mammoths ripping up big chunks of grass, exposing soils and allowing other plants to recolonize. Furthermore, mammoth dung helped keep nutrients cycling through the environment (rather than locked up as in tundras). Loss of the mammoths would reduce the plant turnover and nutrient cycling, and the biome was replaced with tundra.
In summary, human impacts on the megafauna does not seem to have been direct overkilling of all the victims by any means. Instead, environmental effects cascaded from the loss of certain taxa, making other species vulnerable to extinction from other causes.
Last Holdouts: The last Megaloceros survived until a mere 6 ka (that is, 4000 BCE); and mammoths and some Caribbean ground sloths as recent as 4 ka (a mere 2000 BCE)! However they only survived on islands or other environments isolated from human presence.
Trophic Shift for Humanity: Previous to the megafaunal extinctions, humans could continue to expand their food base by extensification. But when the continental regions were fully occupied, and much of the easy meals (megafauna) gone, we begin to see a shift in how humans obtain their food. This is intensification: increasing the yield (productivity) of a region by modifying the landscape. The extreme version of this is agriculture, but historically there are other methods as well.
For example, some Native American foragers would clear cut patches in the forest, which tended to attract animals which prefer the forest margin habitat. An extreme example is the "fire stick farming" method in Australia (and probably elsewhere), where local foragers would light grasslands on fire to drive forth prey and cook in place tubers, burrowing animals, and so on.
But the most important form of intensification is indeed farming: domesticating wild species of plants and animals to become the primary food source. First in the Fertile Crescent (Mesopotamia and the Near East) around 10 ka, and later spreading to (or independently developed in) other parts of the world, this allowed far greater local populations. The ability to generate a food surplus (especially one that could be stored) allowed for the development of specialist disciplines such as chiefs, soldiers, metallurgists, scribes, and priests. Because of the development of writing we can actually read the thoughts and ideas of these peoples, and their own ideas about the world and where it came from.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}