

Key Points
- The early history of Hominina, and even of the genus Homo, is restricted to Africa. But around 2 Ma species of Homo spread out into the Near East, and from there to Europe and to central, eastern, and southeastern Asia.
- Early branches of Homo were relatively small, but soon species approximately as large as us evolve.
- Around 2.6 Ma or so, we begin to see humans eating larger animals: initially as scavengers, but eventually as hunters. This includes deferred consumption of large carcasses. Related to this, by around 2-1.5 Ma, we have evidence of fire-use among humans, for warmth, cooking, and so on.
- Also around the the "pithecanthropine"-grade, humans evolve some our distinctive features, such as naked skin (initially becoming dark to protect from UV), increased sweat gland density, elongate legs, and running stamina. These attributes are associated with a traditional mode of persistence hunting: relentless pursuit at a faster prey item until it weakens from exhaustion.
- Derived Homo contains three major clades: the robust European and west Asian Homo neanderthalensis lineage; the also-robust East Asian Homo longi ("Denisovan") lineage; and the more slender African Homo sapiens lineage.
- After the rise of Homo sapiens, there is eventually a shift of behavior that includes finer-made tools, more use of sea food, more symbolic art; and other features that may point to more complex symbolic language.
- During the last 200 kyr, the number of human species decreases from about 8 worldwide to (by 11.7 ka) just us.
Below is a (possible) phylogeny for Hominina (Homo and all taxa closer to Homo than to Pan), as seen in last lecture:
Eurasian "habilines": "Habiline"-grade species include some of the first to move out of Africa. The 1.85-1.77 Ma Dmanisi, Georgia, site has abundant remains of a small-bodied homininan (or multiple homininan species?). Originally considered early specimens of Homo erectus (and still considered such by some paleoanthropologists of an extremely lumpy variety), others recognize it as a new more basal species:
H. georgicus. At this time the rifting between the Arabian and African plates was incomplete, and there was still land connection between the Red Sea and the Gulf of Aden. Additionally, the world was wetter, so that instead of a Sahara and Arabian desert, this region was a relatively-continuous "savannahstan" connecting the African savannas and the steppes of Eurasia. Not just homininans but other African mammals moved into Eurasia at during this interval.
It has to be said, however, that there is considerable variation in the skulls of the Dmanisi homininans, and some studies suggest that multiple species are actually present here. Phylogenetic studies have not resulted in a single consistent position for this material: some place it very basal, at about the same level as Homo floresiensis (which it resembles in size and overall development); other features put it closer to the H. ergaster/erectus-"pithecanthropine" level, and some even suggest it is derived from H. erectus. These alternative positions are reflected in the multiple uncertain placements on the cladogram above.
There is another habiline-grade small-bodied species similar in many ways to H. georgicus but FAR more recent. This is
Homo floresiensis, the "hobbits" of
Flores Island, Indonesia. These are 1.1 m tall homininans from Indonesia known from fossils that are incredibly young: merely 100-60 ka based on the bony material, and 190-50 ka for their tools. (Until 2016 it was thought they survived MUCH more recently than that: a mere 74 to 12 ka (and tools going back to 95 ka): in other words, they almost made it to the Holocene.) Even with the revised stratigraphy and dates, H. floresiensis is among the youngest non-sapiens stem-humans known: contemporary with almost the last Neanderthals.
The "hobbits" were an island isolated population, and thought initially to be an insular dwarfed form of H. erectus, but details of the skeletal anatomy show that they are fairly basal within Homo. (There is even a school of thought that they are simply abnormally-developed H. sapiens, but this is at odds with both skull and skeletal observations.) Only recently discovered, these specimens hint at a greater diversity of human-line peoples even at the recent past. Phylogenetic analysis places them as nested among "habiline" species such as H. habilis, H. sediba, and H. rudolfensis.
This just in for 2019! Yet another small Asian homininan has been reported: Homo luzonensis of the Philippines. Dated to about 67 ka, this appears to be similar in size to H. floresiensis; what little is known of its body (mostly a foot bone...) is different from either the "hobbits" or H. sapiens.
The presence of one definite (and one or more possible additional) habilines within Eurasia in the Pleistocene--including one at the far end of how far you could walk by land during a glacial maximum--suggests that there are other similar forms yet-to-be discovered throughout Eurasia.
Introduction
Out of Africa, Version 1.0
Phylogeny of the Homininae, Continued

{kind=link}
{kind=link}
{kind=link}
"Pithecanthropines", Carnivory & Persistence Hunting
Some have grouped the species of this grade simply into the one species Homo erectus, while in the late 19th and early 20th Century different specimens were placed in various genera (Pithecanthropus, Sinanthropus, Javanthropus, Telanthropus, Tschadanthropus). (Yet another hominid specimen once lumped into H. erectus is Meganthropus palaeojavanicus. A 2019 study shows it isn't a hominine, but rather as pongine--that is, a stem-orangutan.) We will follow the generally partition of recent paleoanthropological work: the named species (2.27-0.87 Ma African Homo ergaster, 1.85-0.070 Ma truly Asian Homo erectus), and 1.2-0.936 Ma European/North African H. antecessor. However, we will use the traditional grade-name "pithecanthropine" for this phase of human diversity.
{kind=link}
{kind=link}
{kind=link}
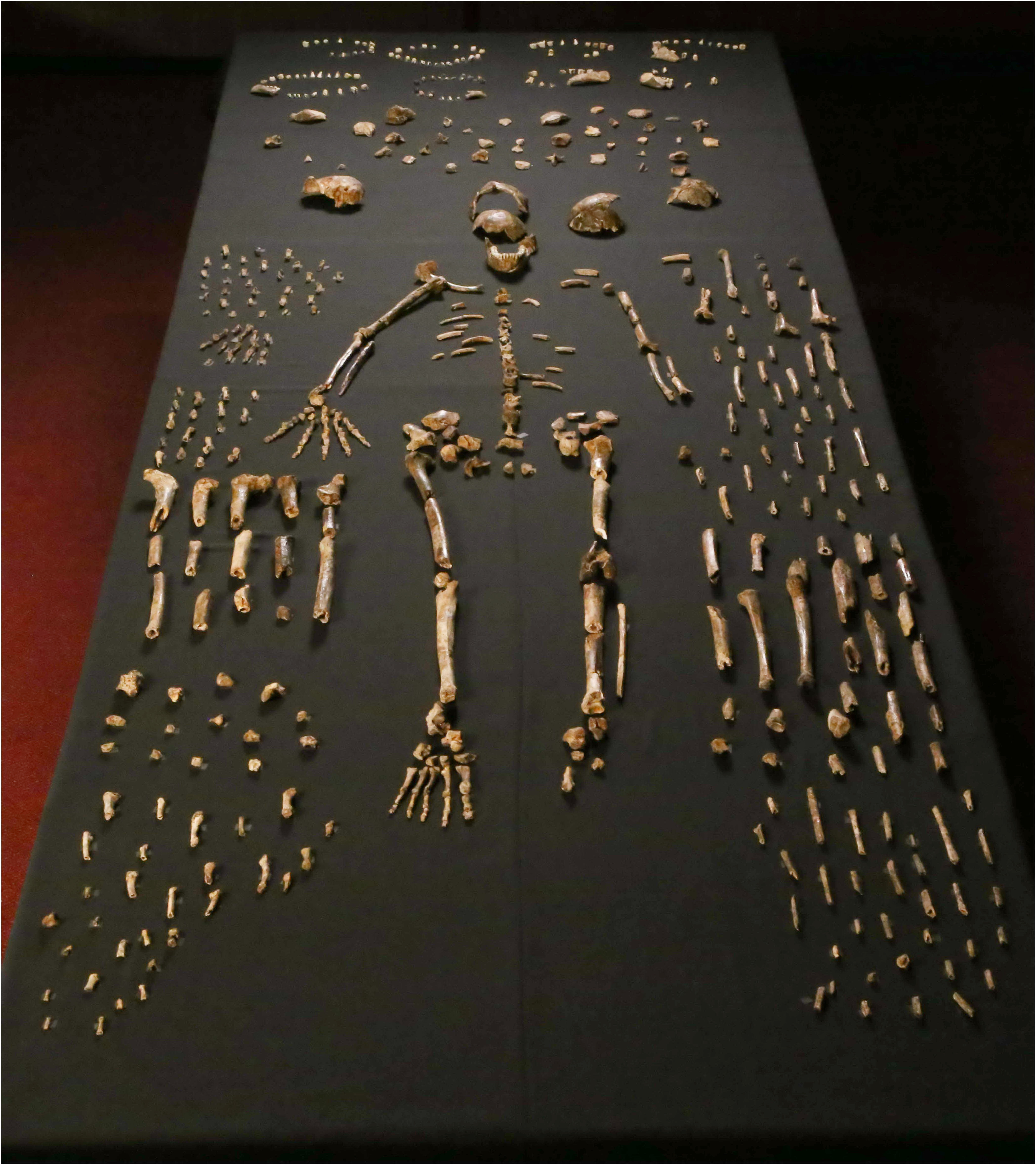
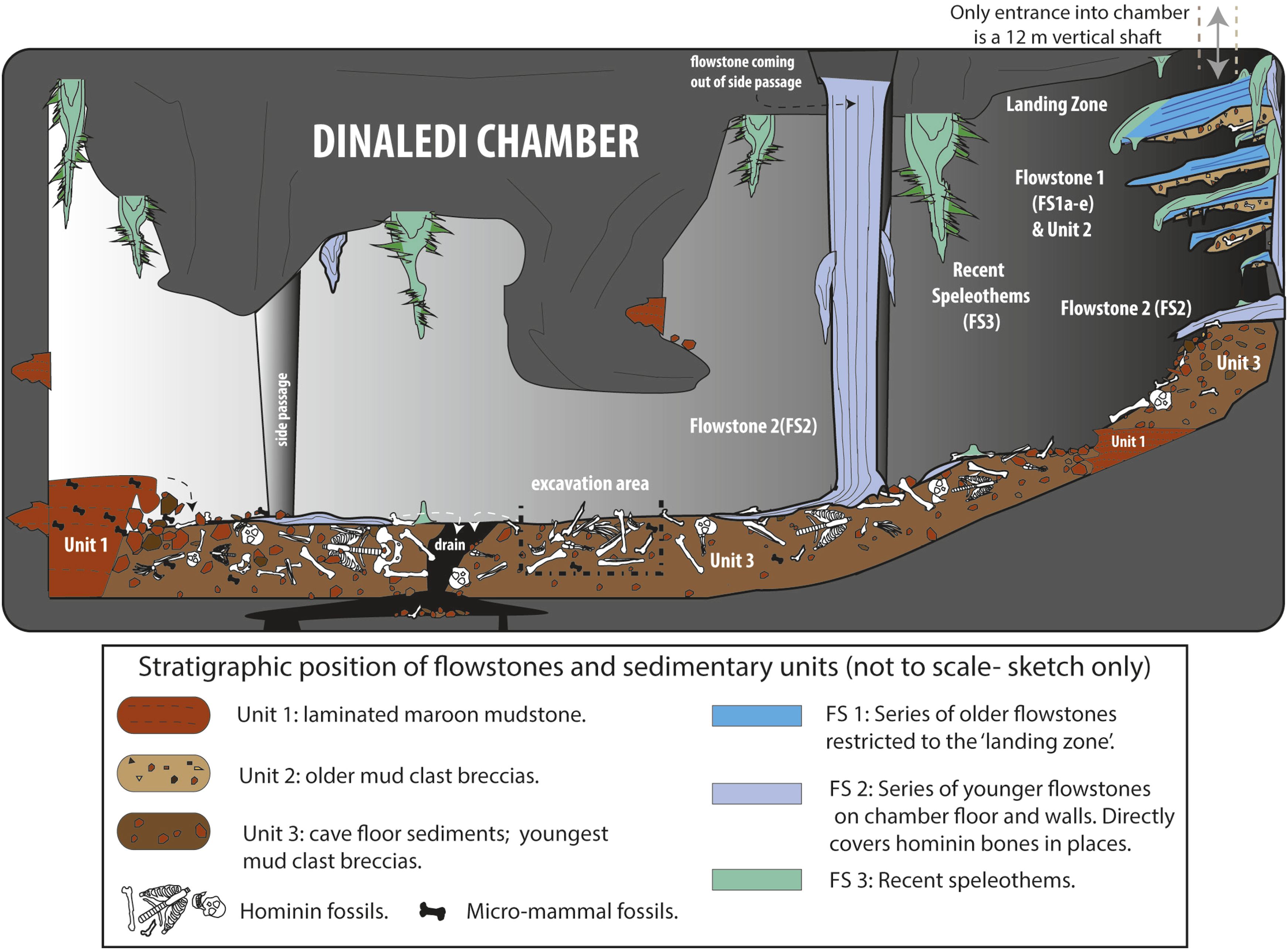
A relatively new discovery is Homo naledi of South Africa. Found deep in a cave, this species is still under study. Initially thought to belong in the "habiline" part of the tree, new studies suggest it is higher up among the "pithecanthropines"; however, some paleoanthropologists consider there to be multiple species represented among these fossils. Originally, reliable age dates were not yet available for it: it was first thought to be somewhere between 3 and 2 Ma, then sometime between 2 Ma and 900 ka, possibly younger. But in May 2017 a higher resolution stratigraphy finds a much younger age: 335-236 ka.
{kind=link}
{kind=link}
{kind=link}
There is a considerable increase of body size (heights up to 1.85 m are known) and of relative brain size with the rise of pithecanthropine-grade Homo. There is circumstantial (and not always convincing) evidence for the use of fire by these peoples going back to about 1 Ma. The diet includes a considerable fraction of meat of large animals, including mastodonts and mammoths. (This trait does go back to more basal forms: even the habiline H. floresiensis seems to have hunted the dwarf species of Stegodon on Flores, and trace fossils show feeding on large mammals by 2.6 Ma.) Since even a band of homininins couldn't eat an entire elephant in one sitting, this may indicate the rise of deferred consumption: caching food to eat later and share with others. The meat-rich diet involving giant animals suggests we have reached the point where the human feeding ecology is a technology-based omnivory (as it remains today.)
This meat-based diet is very important for the human lineage for a physiological reason: our gigantic brains. Brains are metabolically expensive: they need a lot of fuel. In the wild most high-energy food stuffs (honey; certain fruits; etc.) are only seasonally available; furthermore they lack the protein needed to help build and maintain brains. In contrast, meat (including fat and marrow) fits the bill very well. So it might not be coincidence that big brains and consistent meat diets show up around the same time in homininan history.
The build of typical "pithecanthropines" is far more modern than earlier phases: although not exactly of the same proportions as H. sapiens, their limbs indicate that like us they could engage in long-distance endurance running. (It has been shown that although most African large mammals are faster than humans, we can cover the same amount of distance per day as they can because we are marathoners rather than sprinters.) There are some very good trackways from "pithecanthropines" (even H. antecessor tracks from England) that show their locomotion was essentially modern. So from the hips down, at least, we are dealing with species that are "modern" (unlike the earlier grades of homininans).
{kind=link}
But the bodies of "pithecanthropines" and later Homo has a lot of attributes that seem nonsensical for animals feeding on large-bodied mammals: the teeth are smaller; the bite forces weaker; and so on. However, technology continues to take over what was once bone and muscle: tools can serve as "external" claws, teeth, etc. Meat which is mashed up is easier to chew and digest than that straight from the carcass; meat which is cooked is even more digestible! Tracing the oldest use of fire can be quite difficult (especially as fire can be produced by non-human agents like lightening and volcanoes). Possible traces of hearths (fire pits) have been described as far back as 1.5 Ma or so, but are fairly rare. Charred bones are identified back to about 1 Ma. Definite hearths, though, with clear signs of excavation, boundary rocks, etc., are only definitely present for the last 500 ka or so in Africa, and younger in other parts of the world. Given the difficulties in preserving and recognizing fire use in the fossil record, the actual origins of this behavior is possibly much earlier than the oldest known example.
Another set of anatomical and physiological changes that seem to have occurred in this part of the tree seems to be naked, dark, and (under exertion, at least) sweaty skin. We lack skin impressions for stem-humans, but the study of human integument has had advances from a number of lines of evidence. Our living sister group has thick dark fur and pale skins, with sweat glands mostly concentrated on the naked palms and soles. In contrast, the ancestral human skin condition is dark, and all humans have greatly reduced body hair and have lots of sweat glands all over the body. These are all interrelated transformations. Genetics shows that the rise of dark skin had definitely occurred in the human lineage by at least 1.2 Ma, and possibly earlier. Dark skin is defense of functionally-naked skin against UV radiation; thick dark fur protects chimps' and bonobo's pale skin, so dark human skin indicates species exposed to sunlight. But why lose protective fur? This is related to thermoregulation. Primates do not pant, nor do we have large snouts equipped with extensive nasal turbinates (the two main methods other active plains-dwelling mammals use to dump heat). So homininans evolved a larger exposed surface and expanded presence of sweat glands as a means of keeping the body cool in tropical settings. These changes seem to have all occurred in the 1.6-1.2 Ma range. (However, it is possible they happened earlier in our lineage.)
(For what it is worth, here is the later history of human pigmentation: while dark skin is useful in tropical exposed regions, it winds up being detrimental in temperate-to-polar regions, because it diminishes the body's ability to generate vitamin D and folate. So populations which moved into colder, darker regions [more about this next lecture] evolved de-pigmentation of the skin. The genetic evidence shows that Europeans evolved their de-pigmentation convergently from Central/East Asian populations. These transformations seem to be geologically-recent (just about 7500-5000 years ago), and also coincide with dietary changes associated with the rise of agriculture and (at least in Europe) lactose tolerance.)
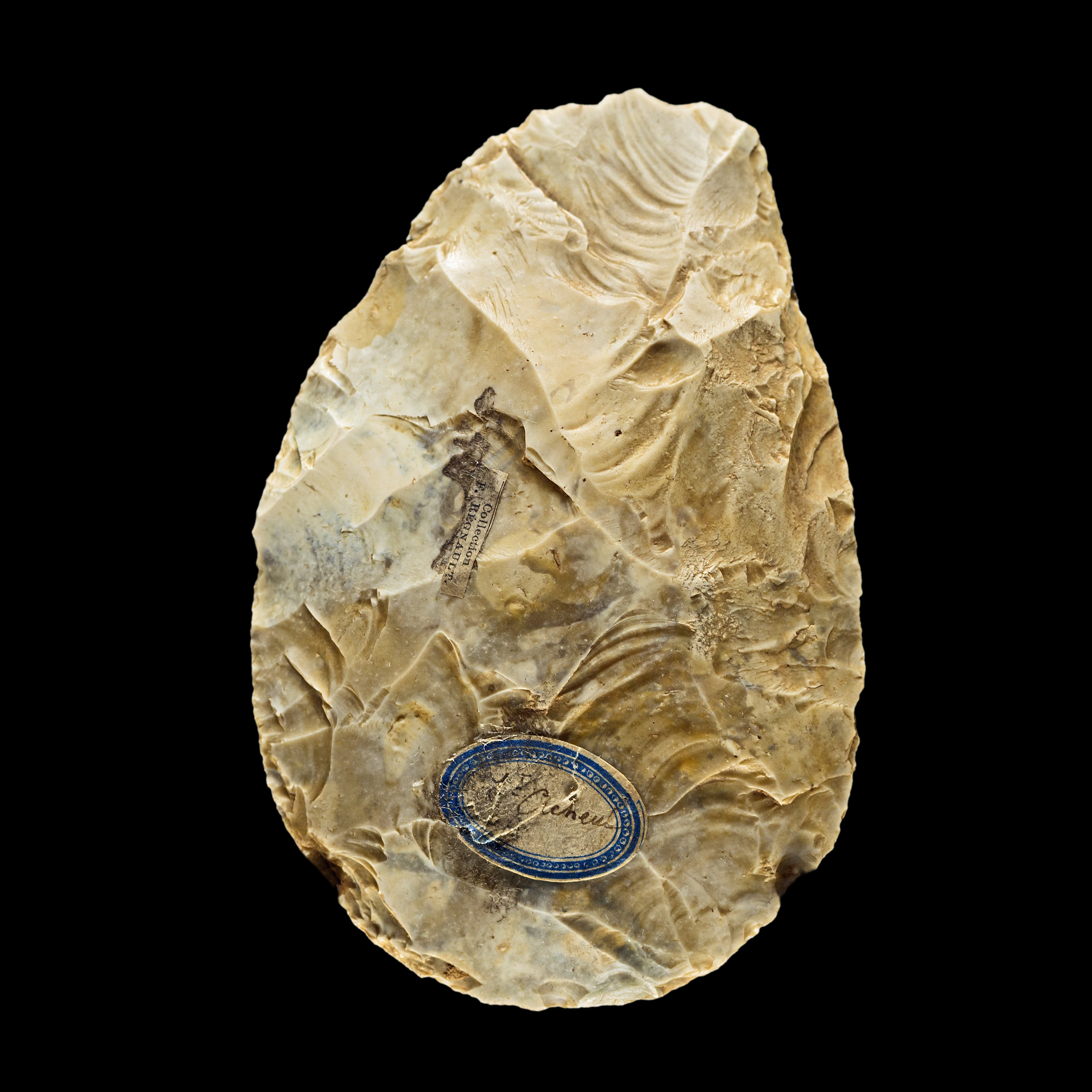
Pithecanthropine-grade Homo (and some later forms) are associated with the Acheulean toolkit with its characteristic "hand axe". Acheulean tools are known from 1760 to about 200 ka.
{kind=link}
{kind=link}
H. ergaster represents the African species of this grade. It is the oldest "pithecanthropine" and might be the likely ancestor of the Eurasian forms. (At one point H. georgicus was considered to be just Eurasian-representatives of H. ergaster.) H. erectus-proper is known from mainland Asia (including the famous "Peking Man" specimens from near Beijing) as well as in Indonesia (the "Java Man" and "Solo Man" fossils). H. erectus is known until about 230 ka on the mainland, and the Java population may have survived to a mere 70 ka! H. antecessor is known from Spain and France, and possibly from some northern African sites.
Derived Homo
In the west (Europe and Africa) appeared very large brained hominins. Sometimes termed the "archaic Homo sapiens", others divide these up into various species, such as H. rhodesiensis of 700-100 ka Africa (from the north to the south), the 500 ka Homo bodoensis of Ethiopia (which is almost certainly just a specimen of H. rhodesiensis), and 260 ka H. helmei of South Africa. Some paleoanthropologists consider this a single diverse widespread species (under the name "H. heidelbergensis"). These same scientists think that H. neanderthalensis and H. sapiens are thus both derived from this single widespread species.) This complex of species seems to split into two clades: a Eurasian and northern African clade (H. cepranensis, heidelbergensis, neanderthalensis, and the Denisovans), and the other (initially) strictly African (H. helmei, rhodesiensis, and sapiens.) However, there are expressions of this phase in Asia as well.
There are two general morphologies (species??) of fossils from China that might represent non-sapiens members of this phase of Homo history. In Dali and Jinniushan (northern China) there are fossils of a robust and large-brained fossils, dating from 260-130 ka. The Jinniushan fossils include a fairly good specimen of a robustly-built female about 169 cm tall and 79.6 kg, with an astonishing cranial capacity 1330 cm2! (This is larger than the average of H. sapiens!) The Dali/Jinniushan species (if they are indeed one distinct species: if so, the proper name would be "Homo daliensis") have been considered by some as an H. erectus-derived form strongly convergent on H. neanderthalensis. More recently, some workers have considered this simply an extremely eastern population of "H. heidelbergensis", or even early members of the H. sapiens lineage. A partial mandible dredged up from near Taiwan (but very poorly dated) might also be from this population. A truly exciting new possibility is that these might actually represent the elusive Denisovan people (about whom we will say more next lecture.)
{kind=link}
{kind=link}
More fossils of these possible Denisovans have shown up in China. In 2017 in central China a series of fossils (including one with an astounding 1800 cm2 cranial capacity: the largest in any individual fossil hominin!) were described from Xuchang. In 2021 a second set of extremely similar fossils were described from 146 ka deposits of northeastern China and given the name "Homo longi" ("dragon man"). At least some of the authors, though, actually do think these are more complete fossils of Homo daliensis rather than a new species. Like the earlier discoveries, these are robust forms with huge cranial capacities (up to 1420 cm2!) Morphologically the robust Pleistocene Asian Homo (whether it is H. daliensis or H. longi or "Denisovans") have some traits of both the sapiens and neanderthalensis lineages; in the cladogram here were are referring to them as "Denisovans" and following the genetic evidence that they are the sister taxon to H. neanderthalensis.
The European line of derived Homo is represented by H. heidelbergensis from 700 to 100-200 ka, and its likely descendant species H. neanderthalensis from 197 to about 30 ka (and even younger, <28.5-25 ka, at Gibraltar). (H. cepranensis of 430-385 ka Italy is considered by some a distinct species, and by others as simply a well-preserved specimen of H. heidelbergensis.) Neanderthals has the largest average brain size of all hominins (yes, bigger than ours: average of 1640 cm2 for a male compared to our 1270, and with one individual with 1736!!) and were very powerfully built (body strength of a 19th Century weightlifter). Their nasal regions were extremely well-developed, and recent studies show would have been very effective at retaining body heat in cold conditions. They show many unique derived characters lacking in all modern humans, and are almost certainly not our ancestors in general (however, they DID interbreed with humans on occasion: more about this later.) Neanderthals lived in Europe and the Mideast, but even as far east as central Siberia, in cold (but not glacial) conditions. Breaks in their bones match those found in rodeo performers, suggesting that they manhandled large animals.
{kind=link}
{kind=link}
{kind=link}
Neanderthals had a VERY meat-rich diet. In fact, they seemed to have preferred the meat of large animals (horses, woolly rhinos, etc.). Even when they ate food from the sea, it seems to be in the form of beached whales and seals. However, a population of Iberian Neanderthals seem to have been vegan, based on their isotopes. Also there is evidence from tooth wear, bits of food found within teeth, isotopes, and fossil association of Neanderthals eating palm dates, legumes, pistachios, water lilies, and (possibly cooked) grains (barley, in particular). A curious possibility has been suggested for some of this plant food in the diet: it may have been the stomach contents of herbivores that were eaten, rather than plants foraged directly by the Neanderthals.
Overlap between modern humans and Neanderthals was very brief (less than 6000 years). Some have suggested human-Neanderthal mating, but the evidence is highly problematic and is better explained as common juvenile traits rather than hybrid traits. At the end of their history Neanderthals improved their technology, producing Châtelperronian technology: finer than Mousterian, but still lacking in obvious art except for some probably beads for necklaces.
Neanderthals seem to have held out last of all in southern Iberia (including the rock of Gibraltar).
{kind=link}
The sister taxon to Neanderthals are a population known from very few bones but for which we have a lot of genetic material: the Denisovans. The very limited bony material for this group is from 51-28.9 ka. We'll discuss them more in the next lecture.
The technology of the derived Homo species begins with versions of the Acheulean toolkit. There is evidence of spear point use (based on the distinctive impact fractures on bones) around 500 ka, and preserved wooden spears from 400 ka. (Given the low likelihood of preservation, wooden spears might go back much earlier.) (NOTE: in the usage here, a spear is a pointed pole weapon used in the hands; it need not be thrown. A spear specialized for throwing is a javelin.)
The Neanderthal tool kit, the Mousterian, was far more sophisticated than the Acheulean. However, it lacked apparent art, nor did it show much in the way of fine tools, nor did it change much over more than 150,000 years, nor did it vary much from region to region.
Homo sapiens and Elimination Rounds
The "Thinking Humans"
Likely derived from Homo rhodesiensis, the oldest specimens of true Homo sapiens date to about 300-315 ka for specimens discovered in Morocco) (prior to 2017 only definitely by 160 ka.) Anatomically modern humans are present in Africa from this point onward, and there are some in Greece by 210 ka and in Israel from 180 ka. These latter may represent a population inferred by genetic evidence to have left Africa before 130 ka along the southern margin of Arabia, hugging the coastal regions of southern and southeastern Asia, and eventually colonizing Australia (by about 65 ka) under at least some older models of human migration. The oldest Asian fossils of Homo sapiens (from South China) are from about 80-113 ka, and may also represent this phase. A third "out of Africa" phase (at around 50 ka) is the main population to colonize Europe, most of Asia, and from there the rest of the world.
Anatomically, H. sapiens is distinct from H. neanderthalensis and H. rhodesiensis (and other relatives) by:
- A flatter face
- A face which is anatomically lower on the skull (as if it "slipped down"), creating our characteristically-big forehead
- A true chin (forward projection of the base of the lower jaw), as opposed to the sloping-backwards shape of the lower jaw of other relatives
Homo sapiens diets and food-procurement behavior changes relative to earlier homininan species. It is within H. sapiens that we have the oldest evidence of:
- Shellfish in the diet (definitely by 164 ka)
- Barbed harpoon points (and catfish bones in trash middens) by 77 ka in Zaire
- Bows and arrows by 64 ka
- Poison use (on arrow heads) by 60 ka
- Baskets (discovered by impressions of woven plant matter) by 26 ka
- Wild wheat and barley ground and baked at 23 ka in Israel
- Pottery by 20 ka in Japan
Elimination Rounds
Taking a look at the above information, we can track the number of species of Homo present for the last 200,000 years:
- Minimum number of human species at 300 ka (about the time of the oldest H. sapiens): 8
- The lineage leading to H. floresiensis in Indonesia
- Homo naledi, assuming the current dates are correct
- The H. erectus population in Java (but perhaps the mainland H. erectus were already extinct?)
- The Dali/Jinniushan people in northern China
- H. heidelbergensis in Eurasia and northern Africa
- H. neanderthalensis
- The lineage leading to the Denisovans (which might be the Dali/Jinniushan people)
- The first H. sapiens in sub-Saharan Afica
- Minimum number of human species at 100 ka: 6
- H. floresiensis in Indonesia
- The H. erectus population in Java
- The last H. heidelbergensis (if the youngest age dates are correct: if not, it is gone)
- H. neanderthalensis
- The lineage leading to the Denisovans
- H. sapiens, including some in the region around Israel and others along the southern coastal regions of Asia up to South China
- Minimum number of human species at 50 ka: 5
- The last of the H. floresiensis in Indonesia
- The last of the H. erectus population in Java (if they indeed lasted this long)
- H. neanderthalensis
- The Denisovans
- H. sapiens, now across Africa, Asia, much of Europe, and even to Australia
- Minimum number of human species at 11.7 ka (base of the Holocene): 1
- Just us, the "last man standing": H. sapiens, now across in all of Africa, Eurasia, Australasia, and both North and South America
The "Great Leap Forward"
In terms of the early anatomically modern H. sapiens we find a surprising lack of evidence for:
- Finely made tools
- Fishing or net-making
- Long-distance goods exchange
- Decoration (except there may be some Neanderthal beads, and ochre is known from Neanderthal and pre-sapiens African sites)
- Art (paintings, carvings)
- Marked regionalization of culture
- Rapid transformation of technologies
The appearance of these behavioral traits has been called the "Great Leap Forward" by Jared Diamond. Around 50 ka or so we see the rise of representational and symbolic art, finely-made tools, and the like.
What caused (or allowed) the Great Leap Forward? It was once thought be associated with the evolution of a mutant FOXP2 gene, which allows fine motor control of the mouth and tongue. The modern human version of this gene is missing in all other living primates, and seemed to have appeared no earlier than 200 ka (i.e., long after the split between the ancestors of H. sapiens and the H. heidelbergensis-H. neanderthalensis line. However, with our understanding of the Neanderthal genome, it appears these peoples had the same version of the FOXP2 gene we do.
ALL modern humans are descended from African populations; we are all the scatterlings of Africa. Some voyages (throughout Europe and continental Asia, into glacial lands, over the land bridge to the New World) were on foot. Unlike other homininans, we have a substantial amount of seafood in our diet: fish, shellfish, and the like. So many human cultures lived near the coastline. (Incidentally, since much of H. sapiens history is within a glacial rather than an interglacial, sea levels were much lower then, and most of the coastal sites of those time are now underwater.
Additionally, H. sapiens seems uniquely to be a boating species. Modern humans reached Australia by 65 ka (based on the maximum age for the Madjedbebe rock shelter locality in northern Australia; others argue no date older than 42 ka in Australia is secure), but there was no land connection at the time; in contrast, the Neanderthals never made the much shorter crossing from Gibraltar to Africa. And boats may have helped humans colonize the New World: controversially as early as 30 ka, unquestionably by 13 ka (but with new strong evidence for 21 ka!).
Arrival of modern humans (or at least modern humans equipped with certain levels of technology) seems to result in mass extinction of many local animals: around 46 ka in Australia; around 14 ka in glacial Eurasia; around 11-13 ka in the New World; and many, many times in islands around the planet.
From the point of divergence from the ancestors of chimps through all this times, humans seem to have lived in small bands. Comparisons with both non-agricultural human societies and with non-human hominids suggests these were almost all bands of about 30 or so at most, made up of closely related individuals: everyone would be a first or second cousin. There would be occasional exchange of genes between neighbors, but aggression between them would help maintain cultural and linguistic separateness. At the larger end would be tribes: units of a few hundreds, still closely related by birth and maintaining identity by means of unique customs, languages, and a near constant state of low-level "warfare" (although having far more in common with "drive-bys" than with later wars). The ability for any one band/tribe to conquer or absorb another would be relatively limited.
Food acquisition would be various combinations of hunting and gathering (collectively called foraging). Foragers can have a very good diet, but in order to expand a population it has to grow by "extensification": spreading out to new lands. (Later on, humans develop methods of "intensification": increasing the productivity of the land, ultimately becoming animal husbandry and gardening.) Because of a reliance on extensification, and our amazing tolerance to different environmental conditions, H. sapiens began to spread out to all corners of the world.
We can now answer many questions about human origins with great degree of certainty:
- Where did Humanity come from? Africa.
- What did Humanity come from? A lineage of apes adapted to more open-land environments.
- What were the first people like? Depending on how broadly you define "people", either very similar to modern chimpanzees (i.e., "people" = "Hominina"), large-brained long-legged animal possessing control of fire and a relatively sophisticated tool kit (i.e., "people" = "Homo"), or identical to modern people (i.e., "people" = "behaviorally modern Homo sapiens").
- How did the first people live? Regardless of the definition of "people" (see above), almost certainly in small bands practicing hunting and gathering.
So think about it: our physical bodies, our physiologies, and our behaviors evolved in conditions that nearly none of us now live!
Some Relevant Videos