

Key Points
- Humans originate among Primates, nested within "monkeys" (Anthropoidea) and "apes" (Hominoidea)
- Initial discoveries of early humans were mostly in Eurasia, leading some to conclude this was where our lineage arose. Similarly, many paleoanthropologists considered large brains to have evolved prior to upright stance.
- Additionally, many considered the great apes ("Pongidae") to be a monophyletic group, the sister taxon to the human lineage.
- However, paleontological and genetic studies confirmed that many of Darwin's original speculations were correct: humans are nested phylogenetically among the African apes (Hominidae), and Africa rather than Eurasia was our ancestral homeland.
- Additionally, evidence showed that upright stance evolved well before the expansion of brain size, such that early stem-humans (members of Hominina) were essentially just bipedal plains apes.
- Early homininans were relatively diverse, with some adapted to life on the margins of forests, and others in the open grasslands. Their diets varied from more omnivorous to more specialized grain-and-tuber eaters.
Traditional cultures (and a fair number of modern people!) do not consider humans to be animals. But in fact, were are indeed part of Metazoa (animals). Once people began to accept that humans were a kind of animal, the question arose: what KIND of animal? Or, to put it another way, what are Humanity's closest relatives?
Some traits of Homo sapiens are widely shared with other species (i.e., warm-bloodedness, or skeletons, or multicellularity, or DNA). Others are shared only with our closest living kin. Still others are unique to us among living animals, but we inherited them from our now-extinct ancestors and kin. And finally, some traits ARE unique to Homo sapiens.
Primates are a highly diverse group, including such forms as lemurs, lorises, tarsiers, platyrrhines (New World monkeys), cercopithecoids (Old World monkeys), apes, and humans (among the living forms).
A common statement from people (even well-meaning people who support evolution!) is:
"Okay, so humans are related to monkeys and apes, but we are not descended from monkeys and apes, right? It's just that we share a common ancestor with monkeys and apes, right?"Introduction
"Light Will Be Thrown on the Origin of Man and His History"
Humanity Among the Animals
WRONG!!
In fact, "monkeys" and "apes" are paraphyletic series. Old World monkeys are more closely related to apes and humans than they are to New World monkeys; chimps and bonobos are the living sister group to humans, and more closely related to them than to gorillas and orangutans and gibbons; gorillas are more closely related to chimps + humans than to orangutans and gibbons; orangutans are more closely related to African apes and humans than they are to gibbons. Thus, some apes are more closely related to humans than to other apes. Hence, humans ARE a kind of ape and descended from other apes (the concestor of humans and chimps, and of humans and gorillas, and of humans and orangutans, and of humans and gibbons would be called an "ape" if we were to see it. Similarly, the concestor of New World monkeys and of humans and apes would be a monkey, and of Old World monkeys and of humans and apes would be a monkey. These would not be any LIVING species of ape or monkey, but would conform to our understanding of "ape" or "monkey" by any reasonable definition.)
(As we will see below, there was a time (particularly in the early and mid-20th Century) when some evolutionary biologists actually argued that apes and humans were separate clades from each other descended from a common non-ape, non-human primate.)
A History of Human Origins Studies
The word "Primates" (a Latin word with three syllables, pronounced "Prime-ate-ees") is not a traditional name for a group of animals. It was proposed by Linnaeus in his Systema Naturae, and means "the first ones". It isn't that Linnaeus thought these were the first animals who ever lived; it is named after a much more mundane reason: they are literally the first group of animals discussed in the book! Linnaeus' Primates included some animals (like bats) we don't include among primates anymore. (By the way, when in lowercase, the word is two syllables: pronounced "primates".)
The very first genus listed by Linnaeus is Homo (humans), and the first two species are Homo sapiens (us) and Homo troglodytes. The second animal was not based on a type specimen, and as described as a mishmash of chimpanzee and orangutan traits. Another species (listed among Linnaeus' "monkey" genus Simia) is Simia satyrus, which is also a mishmash of chimpanzee, orangutan, and Old World monkey traits. In later decades, other authors would give a genus name for the first of these ape species, to become Pan troglodytes (chimpanzee).
[Soap-box time! One of the STUPIDEST things to come out of the mouths of people who should know better--such as medical doctors and science reporters--is to say "humans and primates". Humans ARE primates! Humans have ALWAYS BEEN primates! There has NEVER been a time when the word "Primates" was used as a formal taxon that humans weren't in it. In fact, you would have to come up with a new name for the rest of the group (for example, see next); we are the truest primates of all!]
In the 19th Century, Primates were subdivided into (or in some cases simply replaced by) the Bimana (humans, the "two-handers") and the Quadrumana (all other primates, the "four-handers" [because of the grasping ability of both the hands and feet]). This usage continued even into the Darwinian era (in fact, Darwin himself used it): the Bimana were then interpreted as evolving from the Quadrumana.
Studies comparing humans to non-human primates reached their climax in the conflict between two giants of 19th Century paleontology and anatomy, both of whom we have already met: Sir Richard Owen and Thomas Henry Huxley. Owen claimed that there structures in the human brain (in particular, the hippocampus) which were not present in the brains of other primates. From this, he argued that humans could not evolve from other primates, and indeed this fit his rejection of descent with modification in general. In contrast, Huxley showed that the hippocampus was indeed present in other primates, and that a gradation extends from humans, apes such as gorillas, chimps, and orangutans, and then "lower primates" and other mammals.Darwin did not discuss human origins very much (just one short line: see the top of the page) in The Origin, but devoted his one of the two volumes of his 1871 work The Descent of Man and Selection in Relation to Sex to the topic. In there, Darwin observed:
"In each great region of the world the living mammals are closely related to the extinct species of the same region. It is, therefore, probable that Africa was formerly inhabited by extinct apes closely allied to the gorilla and chimpanzee; and as these two species are now man's nearest allies, it is somewhat more probable that our early progenitors lived on the African continent than elsewhere." -- Chap. 6
Thus, there was now a set of testable hypotheses for human origins beyond the observation that humans were a type of primate:
- Humans were more closely related to the African great apes (the chimp genus Pan and the gorilla genus Gorilla) than to all other living animals
- Stem-humans originated in Africa, and spread to the rest of the world from there
But what of fossil remains of stem-humans?
The late 19th Century saw discoveries of early members of the human lineage found in Eurasia, but not yet Africa:
- Essentially modern human fossils from the late Pleistocene, assignable to our own species (Homo sapiens) in Cro-Magnon, France (found in 1868), and other localities
- Somewhat older fossils of a robust, thick-browed form found in Belgium (1829), Gibraltar (1848), and most famously the Neander Valley (Neanderthal), Germany: Homo neanderthalensis
- Even older Pleistocene fossils, smaller brained but fully upright, in Trinil, Java, Indonesia: Pithecanthropus erectus (later transferred to our genus as Homo erects)
These were all relatively derived species (even "Pithecanthropus" [literally, "ape-man"]). Many speculated that the ancestors of these forms would likely have been a big-brained animal with a sloping back. The idea was that brains are what make humans humans, so that this "essential trait" evolved first. Upright stance would have evolved later, freeing the hands to allow tool use. However, this model was build without physical evidence. (And indeed the Piltdown hoax was designed to play off of just this particular model.)
Many early 20th Century paleontologists (e.g., H.F. Osborn) considered the stem-humans to be independently derived from an ancestral "higher primate" than the "Pongidae" (great apes: chimps, gorillas, orangutans) and the gibbons as far back as the Miocene or Oligocene (and thus were separate lineage for 10s of millions of years). (This is the origin of the idea that humans and apes "derived from a common ancestor that wasn't an ape or a human"). Still others argued that humans, great apes, gibbons, Old World monkeys, and New World monkeys, all derived independently derived from some tarsier-like ancestor in the early Cenozoic. And many of these argued that the concestor of these groups lived in Eurasia.
In the 20th Century, however, evidence from sub-Saharan Africa began to come to light that radically changed the mainstream of paleoanthropology (but which, as it turned out, confirmed Darwin's two primary hypotheses). In 1924 a small gracile form Australopithecus africanus (African southern ape) was found in Pliocene-aged sediments of South Africa, followed in 1938 by more heavily-built Paranthropus robustus (robust near-man). These forms were fully-upright, smaller-brained, and African.
African paleoanthropology exploded in the late 1950s onward as Richard and Mary Leakey (and their descendants), Donald Johanson, and their various colleagues and rivals, found numerous species of stem-humans ranging from essentially-modern forms of the late Pleistocene to all sorts of Pliocene and latest Miocene "ape-men". Some of their dig sites have become the most famous paleoanthropological sites around: Olduvai Gorge; Lake Turkana; Hadar, in the Afar Triangle; Laetoli; and more. This secured the African origins of humanity, and the sequence of upright-first, big brained-second model.
{kind=link}
In the 1970s, several new lines of data came to play which overturned other aspects of human origins. Since the mid-20th Century the poorly-known primate "Ramapithecus" was interpreted as a stem-human that lived in the mid-Miocene: this would have required a divergence time between the human-line and other African primate line before 13 Ma or so. Also, at this time it was thought that gorillas and chimps (including Pan paniscus, the bonobo) were more closely related to each other than to humans. Reanalysis of the "Ramapithecus" fossils showed that they were simply remains of the already-known Sivapithecus, a stem-orangutan. Furthermore, the new genetic evidence demonstrated that Pan and Homo were more closely related to each other than either were to to Gorilla, and that the stem-Pan/stem-Homo divergence was VERY recent (around 5-7 Ma).
The living members of Primates show the following major phylogenetic divisions:
- Strepsirrhini, including the Lemuriformes (lemurs) and Lorisiformes (lorises, pottos, and galagos)
- Haplorrhini, further subdivided into:
- Tarsiriiformes (tarsiers)
- Anthropoidea (or sometimes "Simiiformes"): monkeys and apes, further subdivided into:
- Plathyrrhini (New World Monkeys)
- Catarrhini (Old World primates), further subdivided into:
- Cercopithecoidea (Old World Monkeys)
- Hominoidea (apes, including humans)
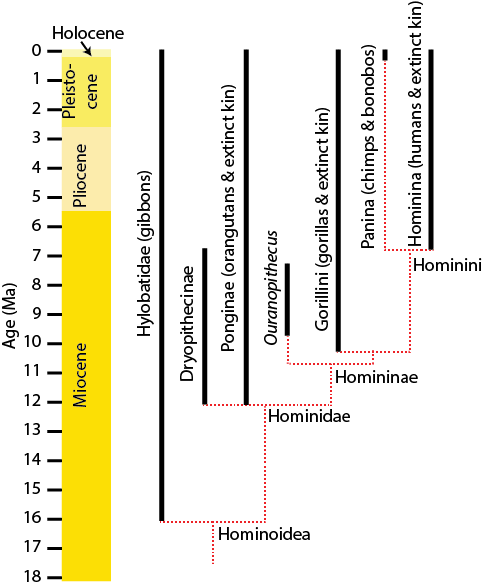
A close-up of the living members of Hominoidea:
- Hylobatidae: gibbons, or "lesser apes"; 17 or so species in four genera; limited to Southeast Asia and Indonesia; extremely acrobatic swinging animals
- Hominidae: "great apes", divided into:
- Ponginae: represented today solely by the two species of Pongo (orangutans)
- Homininae: the African great apes, divided into:
- Gorillini, represented today solely by the two species of Gorilla
- Hominini (hominids), subdivided into:
- Panina (paninans): chimps (Pan troglodytes), bonobos (Pan paniscus), and all (now extinct) taxa closer to them than to Homo sapiens
- Hominina (homininans), Homo sapiens and all (now-extinct) taxa closer to us than to Pan
{kind=link}
Below is a simplified phylogeny for the hominoids, including some fossil clades:
Human Traits?
What features make humans humans? That is, what traits are truly unique to Homo sapiens? And of those traits which are unique to humans among living animals, which ones are shared with extinct stem-humans?
Let's look at traits that have been suggested:
- Morphological traits:
- Fully upright stance
- Greatly enlarged brain
- Facial retraction (that is, essentially no snout)
- Reduced body hair
- A chin
- Precision grip between thumb and all other fingers
- Physiological
- Menopause (most other mammals do not have this; instead, female fertility declines relatively gradually with age, much like male fertility)
- Long adolescence (most other mammals have a very short period between childhood and adulthood)
- Longevity (most mammals of our mass live far shorter lives)
- Behavioral
- Complex symbolic language
- Collective learning (discoveries of one person or one generation get added to the next, and so on)
- Mating pair bonds
- Long distance travel
- Cooperative hunting
- Technological/Cultural
- Stone tools
- Fire use
- Boats
- Art
- Domestication of other animals and plants
{kind=link}
Field and laboratory observations show that many of these are actually present in other animals conversantly, and yet more are shared with us and various other primates (and thus are almost certainly synapomorphies of various clades of Primates). For example:
- Tool use is present in all sorts of animals: woodpecker finches, sea otters, etc.
- Domestication of fungi is present in leafcutter ants
- Art of a sort is present in bower birds
Among traits discovered to be present in various non-human primates which were once thought to be strictly human:
- Chimpanzees engage in cooperative hunting
- Chimpanzees have also been observed to use multiple tools together (i.e., a tool kit)
- Use of spears to stab has been observed in chimpanzees and orangutans
- Stone tool use is established in chimps, Old World monkeys, and New World monkeys
- Symbolic language of a sort has been established in some Old World monkeys, who have specific calls associated with specific potential threats. (It is unknown if wild apes have such, but they do have fairly complex call systems)
- In a laboratory setting, chimps, bonobos, gorillas, and orangutans all show an ability to grasp complex language including syntax: however, although they have this ability we do not know if they use it in the wild
- Similarly, laboratory experiments with apes show that they can solve various complex problem sets
The stem-human trace fossil record (which is given its own name: the archaeological record) shows the presence of tool kits far more complex than any living non-human animal, but still far less complex than Homo sapiens technologies.
On the Road to Phelamanga
In the last several decades, our knowledge of the diversity of stem-human species has greatly increased. In fact, many of these were only found in the last decade. These discoveries have filled in much information about the anatomical changes in the history of our lineage.
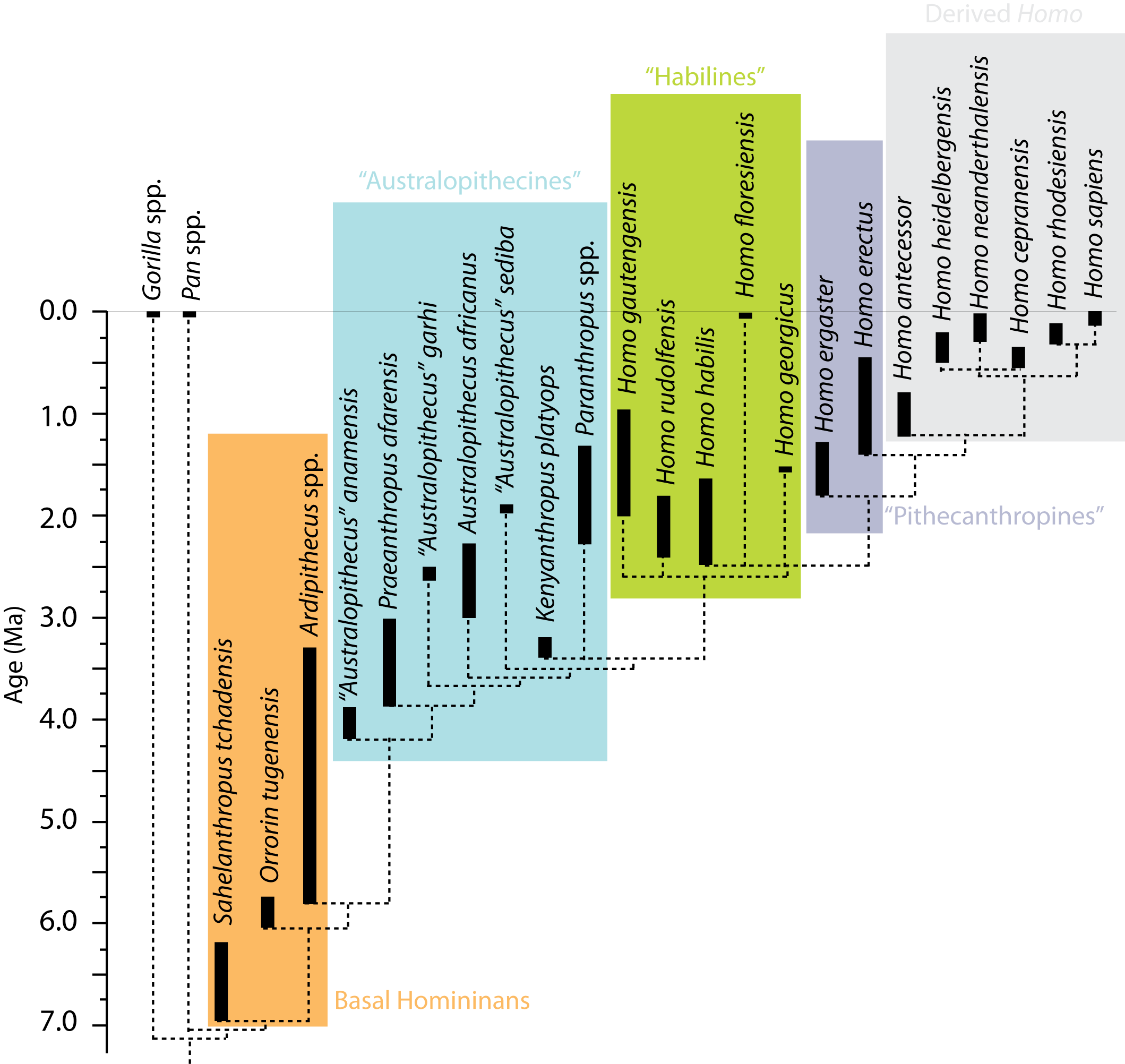
No one analysis has yet included all the fossil homininans. For this class, I have used the most extensive studies so far as the framework of the tree, and then show (by dotted lines) the likely placements of some species (or possible species). (If you want to see the cladogram used in earlier incarnations of this course, it can be found here. You can compare and contrast it with the more recent one.) As with all cladograms, this phylogenetic hypothesis can (and almost certainly WILL) be refuted with new information.
{kind=link}
On top of this, while some homininan species are based on fairly complete fossils (including multiple individuals to give a sense of the range of anatomical variation), many others are based on limited material. Different paleoanthropologists might lump some of these taxa together; others may split some of these species into multiple ones.
Furthermore, paleoanthropologists are somewhat behind the times with regard to the adoption of cladistic approaches to taxonomy. In particular, they tend to use paraphyletic taxa (such as "Australopithecus" for pretty much every species between Ardipithecus and Homo. Below is a phylogeny of some of the more completely-known stem-human taxa, using the genus-level taxonomy of typical paleoanthropologists:
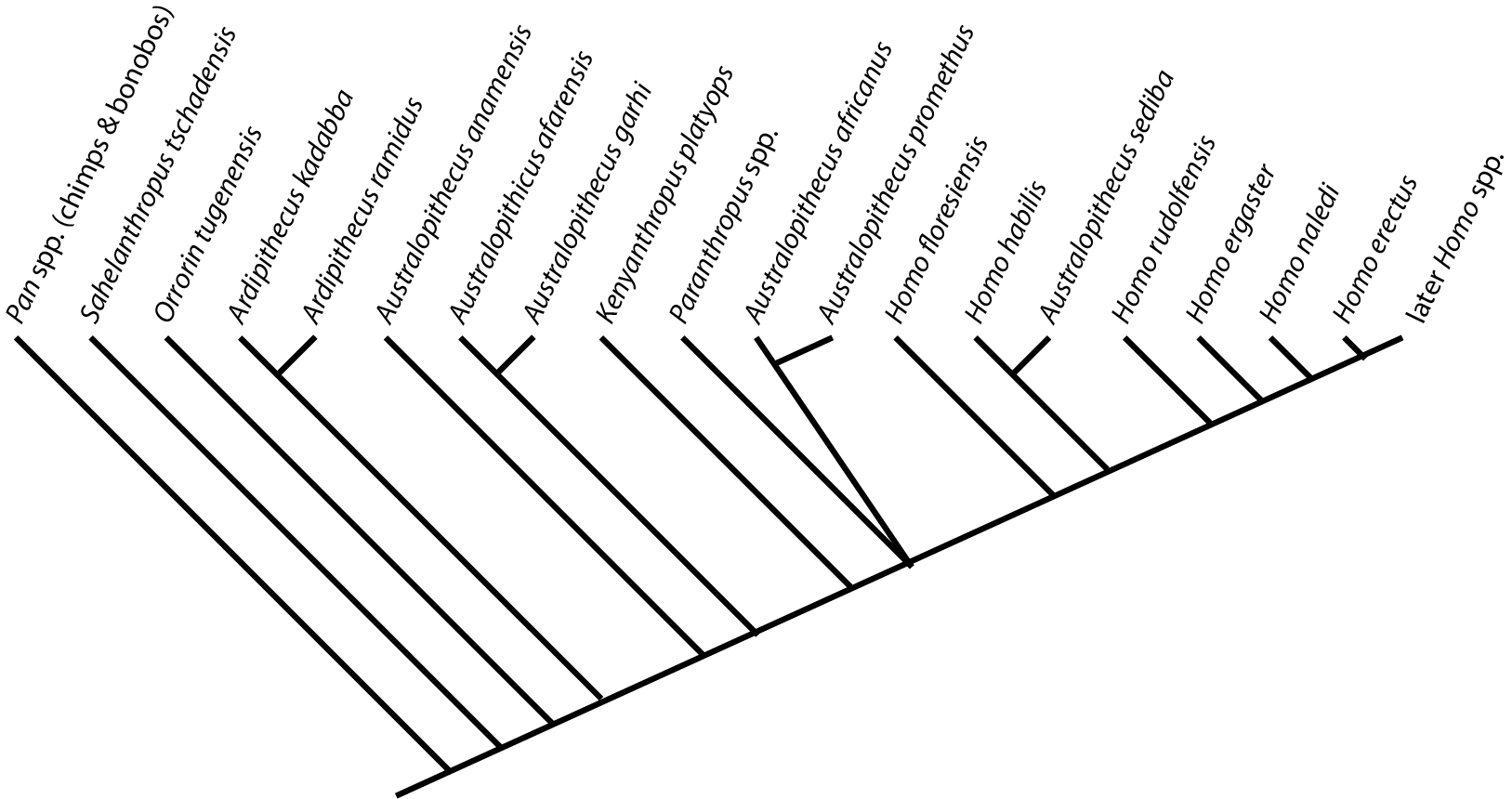
Note how "Homo" is paraphyletic in this, and "Australopithecus" winds up being polyphyletic!!:
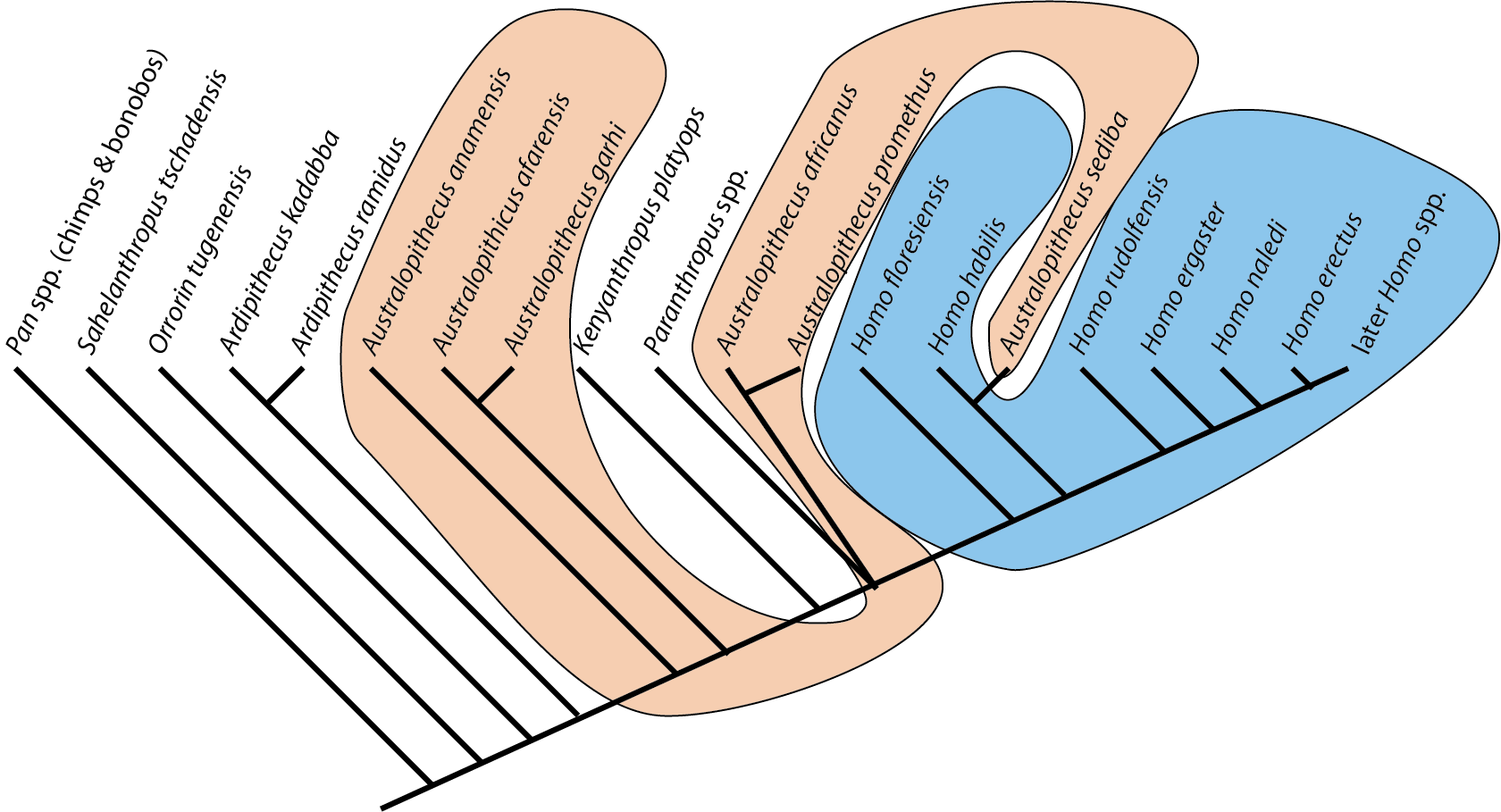
For this course I will use only monophyletic genera. This will result in some names which may be unfamiliar to those familiar with stem-humans: e.g., "Australopithecus" afarensis is called Praeanthropus afarensis:
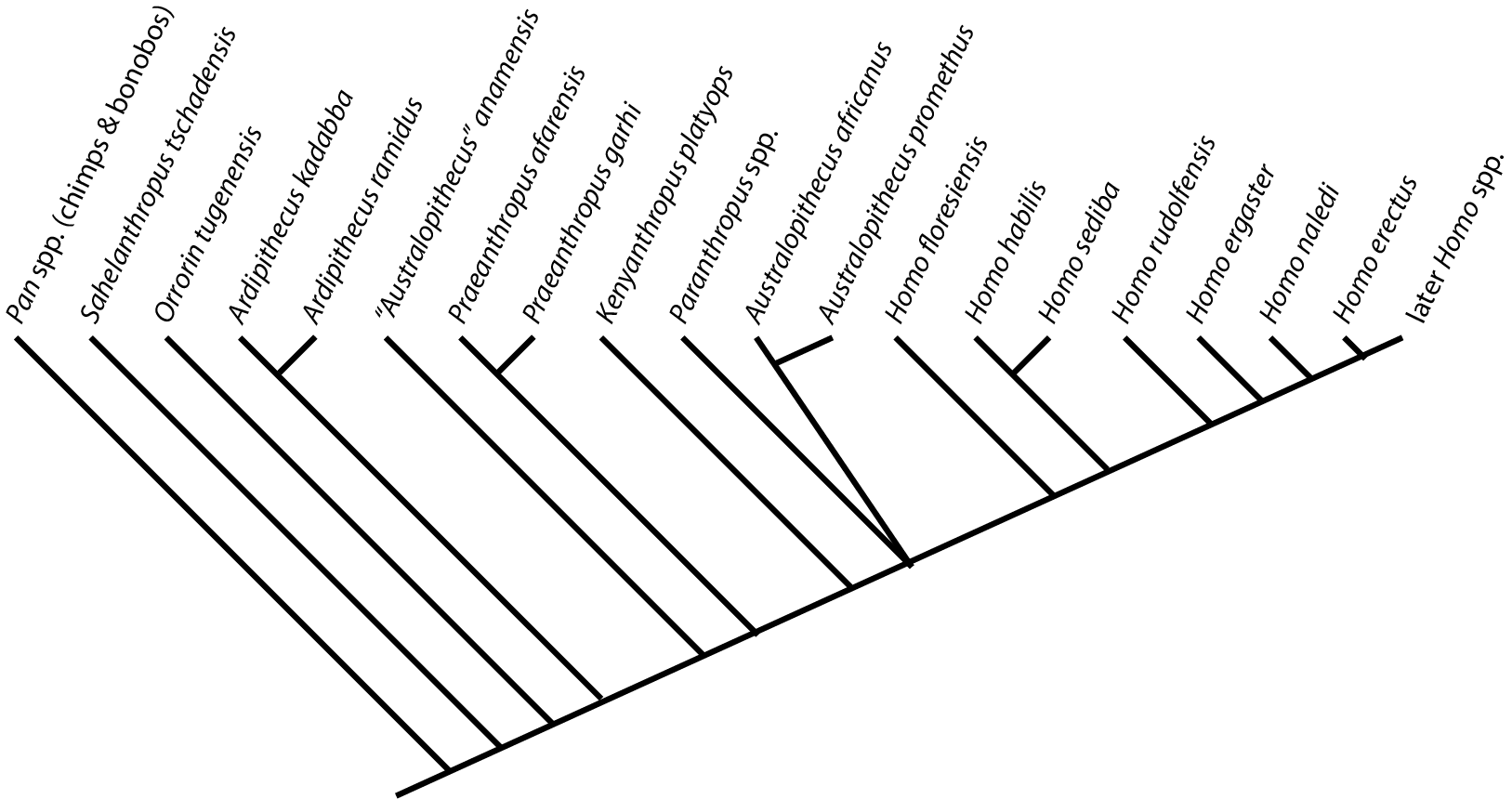
That said, there remain some major points of unresolved relationships and taxonomy. In particularly (as discussed next lecture), the taxonomic names and positions of the populations close to the ancestry of modern humans and Neanderthals is particularly muddied.
Phylogeny of the Homininae
Below is a (possible) phylogeny for Hominina (Homo and all taxa closer to Homo than to Pan):
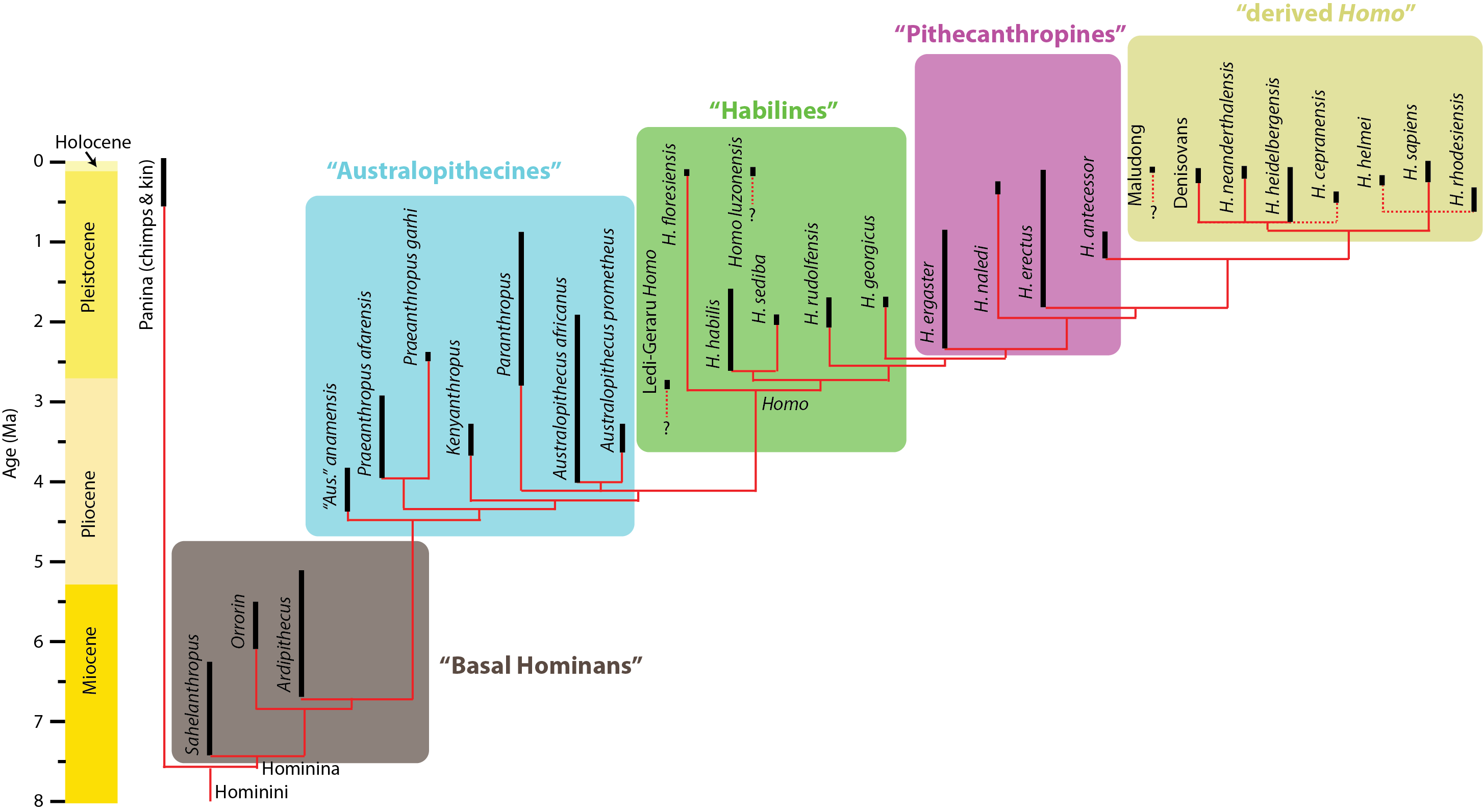
(The solid lines represent relationships discovered in the 2015 study by Dembo et al., with a few additions from genetic analyses and an update including Homo naledi; dotted lines represent inferred positions for taxa who have not yet been subjected to a formal analysis. The age ranges mostly follow the recent summary by Wood and Boyle, with a few more recent updates.)
For the purposes of this course, we won't be overly concerned about the fine details of homininan phylogenetic relationships. Instead, what will be more important is the general trends within the stem-human lineage. For that purpose, I've blocked out a series of paraphyletic grades (and a single clade) that we'll look at as representing the phases of transition on the way from the human-chimp concestor to the modern humans.
"Basal Homininans": The oldest known fossil homininans are approximately contemporaneous with the molecular dates for the split between Panina and Hominina, in the 5-7 Ma range. In particular, the oldest known representative is Sahelanthropus tchadensis (7.43-6.38 Ma) from Chad. Until 2022 it was known only from a single partial skull, which shows reduced canine size and a few other homininan attributes. Newly discovered hindlimb bones show it was most likely bipedal, but forelimb bones show it was still an adept climber. The phylogenetic position of this fossil remains in some dispute: a few researchers consider it to be a stem-gorilla (basal member of Gorillini). Age dates for this fossil are tricky, as it was collected on the surface. The rocks of that vicinity (and the other fossils with which it was found) suggest an age around 7.0 Ma.
{kind=link}
More clearly a stem-human is Kenyan Orrorin tugenensis, dated somewhere between 6.14-5.2 Ma. Fragmentary remains of multiple individuals, including skull elements, teeth, and a number of limb bones, have been found. The limb bones suggest an upright stance, but with grasping hands and feet. The associated sediments and fossils indicate that it lived in a dry evergreen forest.
{kind=link}
By far the best understood basal homininan taxon is Ardipithecus, known from the older A. kadabba from approximately 6.7-5.11 Ma and A. ramidus from 4.6-4.262 Ma: both are from eastern Africa. Known from sufficient skeletal and skull material to reconstruction. It retains basal traits (a chimp-sized brain; a deep rather than wide pelvis; a grasping digit I ["big toe"] of the foot), but shows some homininan features as well (reduced canines; hindlimb bones more upright).
{kind=link}
{kind=link}
{kind=link}
Over all, it appears that the basal homininan grade represents apes adapted to the margins of forests and/or dry rather than rain forests. They were still at least partially arboreal. There is no sign of any higher level of intelligence or technology in these species than in chimps or gorillas or orangutans: they were simply a drier, slightly more upright, variety of chimp.
"Australopithicines": The next phase of homininan evolution is a paraphyletic series that have all been classified as species of "Australopithecus" at some point or another. (A monophyletic Australopithecus would have to be the clade of Australopithecus africanus and everything closer to it than to us.) More colloquially, these species are sometimes called "apemen", since they are generally upright like humans but still had fairly ape-like brains and snouts.
The oldest "australopithecines" (the East African [Kenya and Ethiopia] "Australopithecus" anamensis) are known from about 4.37-3.82 Ma; the youngest (the robust Paranthropus robustus) up to 0.87 Ma. All known "australopithecines" are from Africa.
The best known "australopithecines" represent two different major morphotypes:
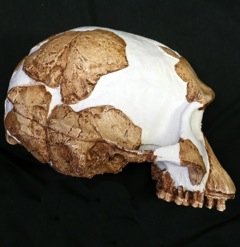
- "Gracile australopithecines". This grade includes: "Australopithecus" anamnesis; 3.65-3.35 Kenyanthropus platypus of Kenya; 3.89-2.9 Ma Praeanthropus afarensis (once called "Australopithecus" afarensis, and famous for the excellent skeleton "Lucy" and "Selam"; also including specimens once called "Australopithecus bahrelghazali" from Chad and "Australopithecus deyiremeda" from Ethiopia); and the related 2.5-2.45 Praeanthropus garhi of Ethiopia. Additionally, well-known 4.02-1.9 Ma Australopithecus africanus itself belongs to this group. These basal forms show the rise of a fully-upright stance (evidenced by body fossils and excellent tracksites); a shift of relative brain size from essentially chimp-like (in Praeanthropus) to larger (in Australopithecus); and a habitat shift from the marginal forests (in Praeanthropus) to the savannas (in Australopithecus).
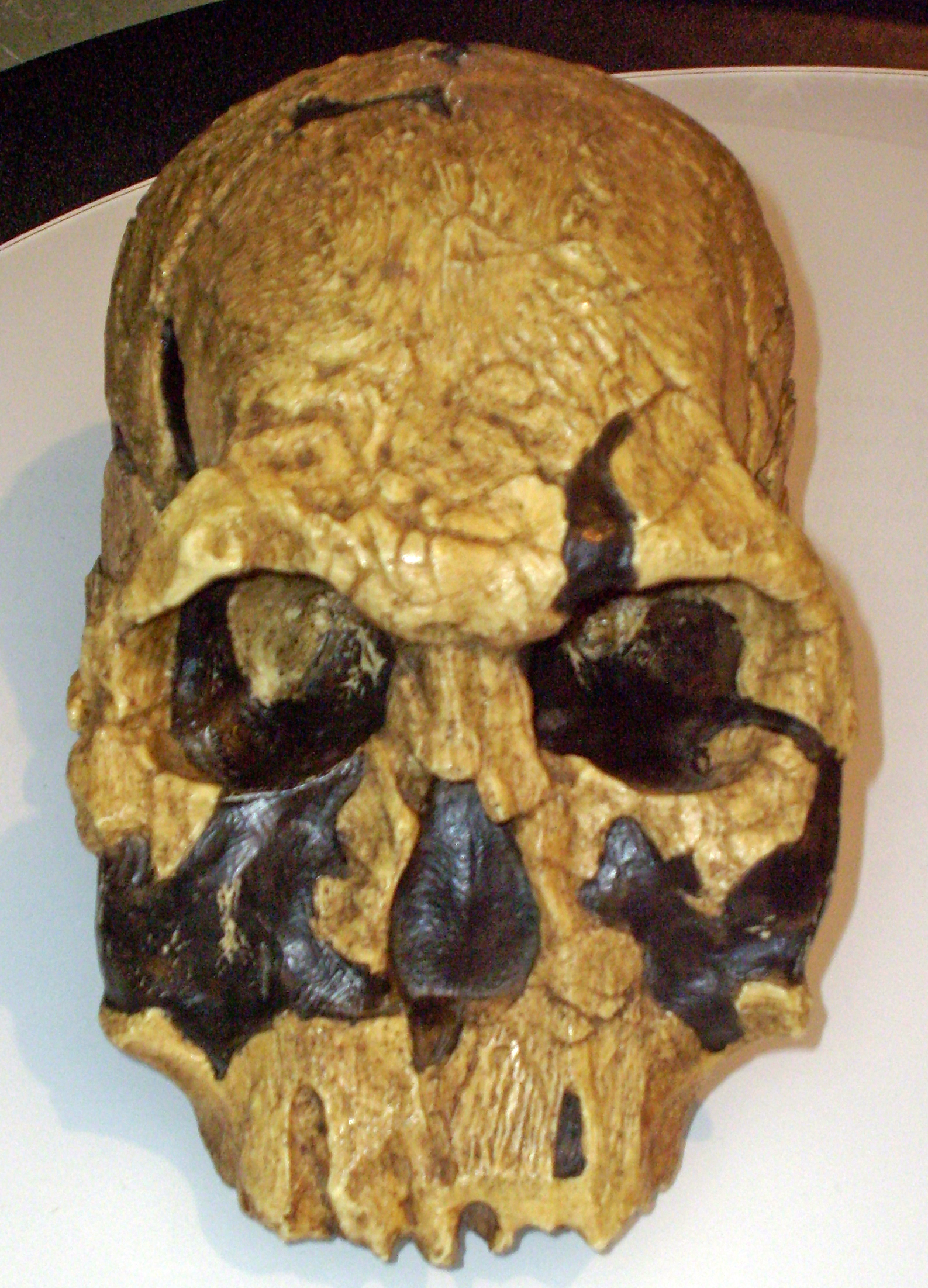
- "Robust australopithecines", which seem to form a single genus Paranthropus. These have greatly enlarged molars and enormous jaw muscles: isotopic and tooth-wear analysis shows that they had a substantial fraction of their diet in the form of grasses, making them essentially primate horses. Paranthropus has a larger brain size than chimps or Praeanthropus, more comparable to Australopithecus proper. Three species are known: 2.73-2.33 Ma P. aethiopicus of Ethiopia; 2.5-1.15 Ma P. boisei of East Africa, and 2.27-0.87 Ma P. robustus of Southern Africa. Paranthropus represents a clade nested within the gracile australopithecine grade.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
"Australopithecines" are associated with the Oldowan toolkit: a simple set of stone-tools shaped and flaked into choppers, cutters, hammers, etc. These are marginally better than non-human living primates can make. This toolkit is known in the archaeological record from about 2.6 to 1.7 Ma, although additional work will likely extend the range.
{kind=link}
The "Australopithecines" appear to have been the "plains apes": hominids adapted to life outside the forest. The savannas themselves were expanding during their tenure: the rise of the modern (Quaternary) glaciation cycles at 2.588 Ma meant that the world began to dry, and forests to retreat. There is a comparable rise in the diversity of other plains-adapted species of other African lineages contemporaneous with the "australopithecine" phase.
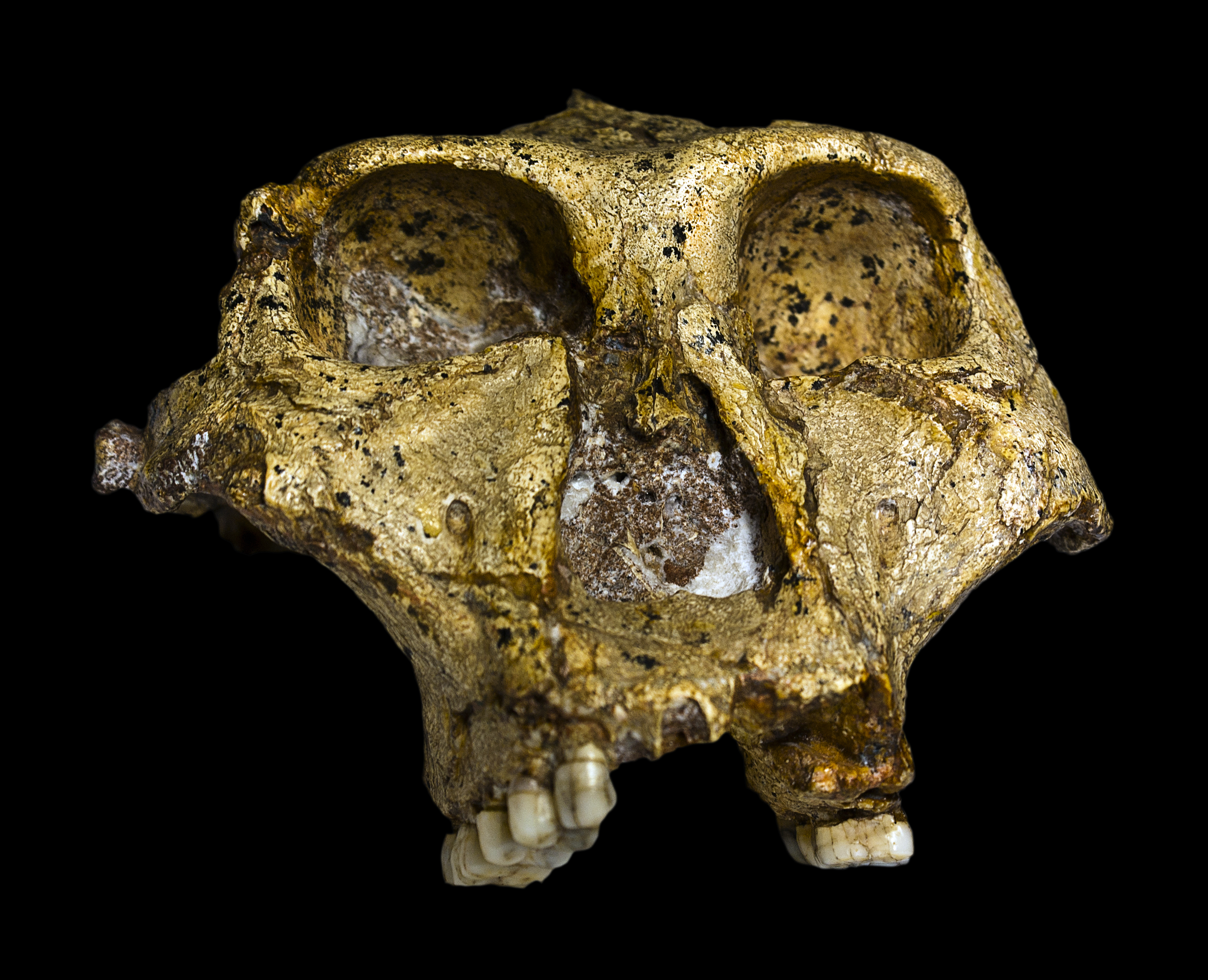
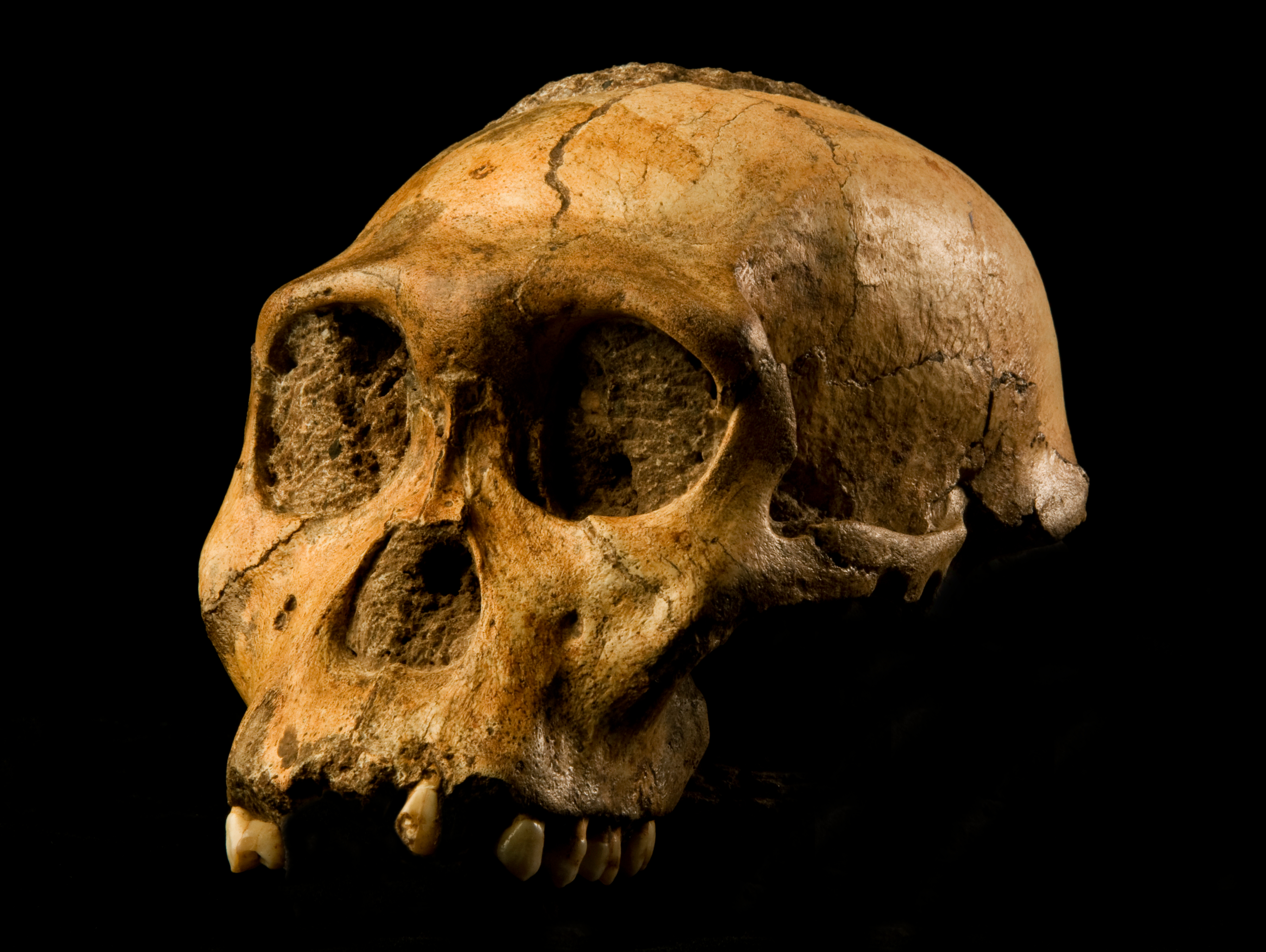
"Habilines": The basal members of the genus Homo show a marked increase in brain size over their "australopithecine" relatives. This phase might be called "habilines", after the first discovered member of this grade: Homo habilis of Eastern and Southern Africa. The oldest well-dated of these (Homo habilis itself, including specimens once placed in its own species "H. gautengensis") ranged from 2.6-1.65 Ma. Its sister taxon, 2.05-1.91 Ma Homo sediba (previously considered a species of Australopithecus) is from South Africa. 2.09-1.78 Ma Homo rudolfensis was very similar to H. habilis in general appearance: indeed, some paleoanthropologists consider all of these the same species.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Some limited fossils probably belonging to an as-yet unnamed species of Homo date to 2.8-2.75 Ma of Ledi-Geraru, Afar, Ethiopia.
All these African habiline species were relatively gracile, but taller than gracile australopithecines (though still only about 1.3-1.5 m tall). In the case of Homo habilis and Homo rudolfensis there is evidence of an increase of meat-eating over more basal lineages. (Some of this was likely from hunting small game, but much of it could have been from scavenging).
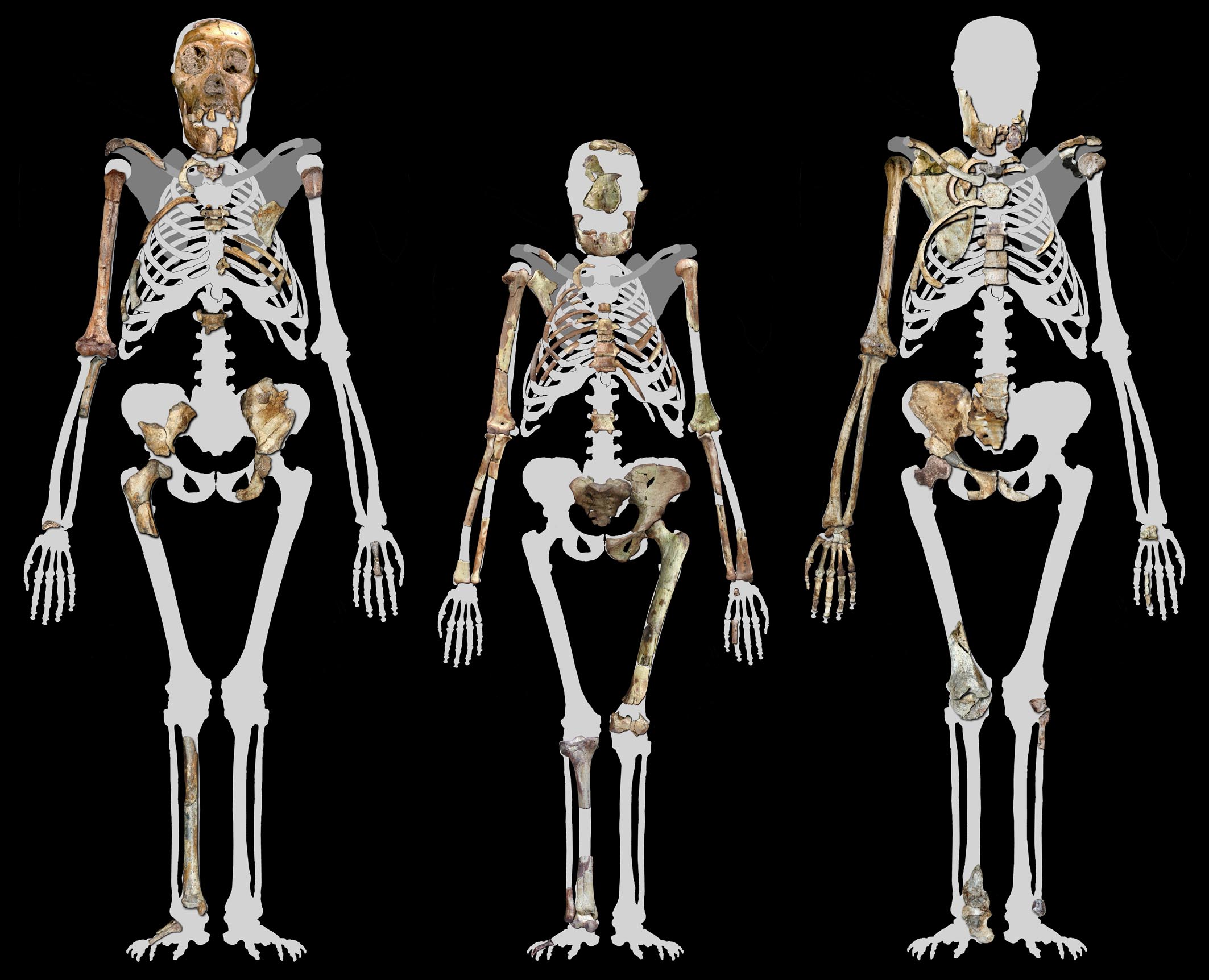{kind=link}
The two species remaining in the habiline grade in the cladogram above (the problematic taxa H. floresiensis and H. georgicus) are the basalmost homininans outside of Africa; their story will be featured in the next lecture.
One pattern is that is beginning to merge is that it appears that "australopithicines" retained the ancestral pattern of marked sexual size dimorphism typical of other hominids, but that this size-discrepancy becomes reduced in the genus Homo.
{kind=link}
Diets of the Proto-Humans
Recent work on the paleoecology of proto-humans (basal homininans through "habilines") have helped reveal a diversity of diets among these species, as one might expect with such a diversity of forms. Lines of evidence include:
- Gross overall morphology of the bones and teeth
- Wear patterns on the teeth
- Isotopic study of tooth and bones
- Analysis of the environments in which they lived
- Archaeological (trace fossil and tooth) analysis at sites
Some of the main discoveries:
- Many homininans have relatively simple tooth wear (limited pitting or scratching or so forth, and often in the same direction). Two exceptions are Paranthropus robustus, and the advanced members of Homo we will see next lecture. This suggests a greater range of diets in these forms than in most of the others.
- Isotopic analysis shows that Homo sediba had a diet strongly skewed to C3 plants, that P. boisei had one skewed very much to C4 plants, and that the others all showed intermediate (mixed) values.
- In the case of Homo sediba it indicates it was primarily a forest-dwelling animal eating forest foods. In fact, combining the type of wear on its teeth and the isotopic values, it may well have eaten a substantial amount of tree and branch bark, as do modern gorillas.
- In the case of Paranthropus boisei, it is possible that it ate a lot of grasses. However, lacking horse-type grazing teeth, it may have eaten a lot of rushes and sedges, including their tubers.
{kind=link}
In summary, we see in the early proto-humans animals which are typically within the size range of chimps and bonobos, and whose brain sizes start as comparable to Pan but which, especially in the case of the "habilines", are proportionately larger.
This changes, however, when we get to 2.6-2.0 Ma, when we have traces of tool marks on large animal carcasses, showing marrow- and meat-eating becomes common. And it is (probably not coincidentally) around the same time that brain size greatly expands.
As we will see, it is among the "habilines" that homininans leave Africa.
Some Relevant Videos