GEOL 204 Dinosaurs, Early Humans, Ancestors & Evolution:
The Fossil Record of Vanished Worlds of the Prehistoric Past
Spring Semester 2026 "What is It?": Identifying Fossils and the Nature of Species
Many skulls of the dire wolf Canis dirus discovered at the La Brea Tar Pits, California
"Of what use are the great number of petrifactions, of different species, shape and form which are dug up by naturalists? Perhaps the collection of such specimens is sheer vanity and inquisitiveness. I do not presume to say; but we find in our mountains the rarest animals, shells, mussels, and corals embalmed in stone, as it were, living specimens of which are now being sought in vain throughout Europe. These stones alone whisper in the midst of general silence." -- Aphorism 132, Philosophia Botanica (1751), Carolus Linnaeus
"I am fully convinced that species are not immutable; but that those belonging to what are called the same genera are lineal descendants of some other and generally extinct species, in the same manner as the acknowledged varieties of any one species are the descendants of that species." -- Introduction, On the Origin of Species by Means of Natural Selection (1859), Charles Darwin
"The usual concept of species can be stated as follows (Mayr 1970): "Species are groups of interbreeding natural populations that are reproductively isolated from other such groups." This concept is grandly called "the biological species concept." But that is an arbitrary appropriation of a term with a more general and earlier meaning. I will instead use the term "reproductive species concept."" -- "Ecological species, multispecies, and oaks" (1991), Leigh Van Valen
BIG QUESTION:How do we identify fossils? What are species?
Parts is Parts: Homology, Analogy & Comparative Anatomy
In order to recognize how organisms are similar, or different, we need to compare its body parts. The important thing is to recognize the equivalent body parts: no sense in comparing a leg with a tail, or a jaw with a stomach. It had been noted by early anatomists that related organisms were built on the same "body design" (in German, Bauplan or "building plan"). In each of these, the underlying structure was repeated from organism to organism: these parts are considered to be homologous.
Homologous structures are the same body part, but might be shaped or modified differently. The wing of a bat, the front leg of a horse, the flipper of a whale, and the arm of a human are all homologous, and have the same basic parts: a single upper arm bone, a pair of bones below; some rounded wrist bones; some long bones in the palm of the hand; and series of long bones down each finger. But even though they are homologous, they have different functions.
In contrast, structures that have the same function but are derived from different body parts are analogous. The wings of bats are modified arms, but the wings of insects are modified gill flaps.
The science of comparative anatomy was developed to describe, compare, and contrast the homologous structures of different kinds of organisms. Given a language of comparative anatomy, we can show how two specimens are similar and how they are different. And we can characterize different types of organisms based on their distinctive combination of features.
Taxonomy: the Naming of Names Taxon (pl. taxa): a named group of organisms.
Naturalists have long noted that there exist units of natural diversity, species, in which the members share certain distinctive features with each other. Traditionally, each culture had its own name for the animals, plants, and other organisms in their region. But EACH culture had its own set of names, so the same type of animal might have many different names. During the 1600s and 1700s, methods were proposed for a formal scientific set of names. Following the work of Carl von Linne' (Linnaeus) in the 1700s (most specifically, the rules he set down in the Systema Naturae ("System of Nature") in 1758; later workers added and modified the system (primarily with the addition of new "ranks")), species were recognized as one unit within a nested hierarchy of larger clusters of organisms: taxa (singular, taxon; literally, "named thing").
Some of the Linnaean rules:
All names are in Latin or Greek, or are modified into Latin form;
Each name must be unique;
All names are fit into a nested hierarchy (species into genera, genera into families, and so forth);
In traditional Linnaean taxonomy, there is a set of official ranks (from smallest to largest, species, genus, family, order, class, phylum) (later workers added additional intermediate ranks, such as tribes, subfamilies, superfamilies, subphyla, etc.);
The primary unit is the species (pl. species)
Refers to a "specific" kind of organism
The next higher unit, the genus (pl. genera) is composed of one or more species
Refers to a more "generic" category than species
Definition of a "genus" is even more problematic than that for species, since it is composed of one or more "species";
Each genus has a type species: all other species are assigned to the genus based on their similarity to the type species;
Linnaean taxonomy has its own special set of grammatical rules:
Genera have one word names (e.g., Panthera, Homo, Ginkgo, Tyrannosaurus);
The genus name is always Capitalized and italicized (or underlined if you don't have access to italics);
Species have two word names, the first part of which is the same as the genus name (e.g., Panthera leo, Homo sapiens, Ginkgo biloba, Tyrannosaurus rex)
The genus name ("generic nomen") is ALWAYS capitalized, the second part ("trivial nomen") is ALWAYS in lower case, and the name is ALWAYS italicized or underlined;
Species names can be abbreviated by using only the first letter of the genus name, followed by a period (NEVER by a hyphen): H. sapiens and T. rex are correct;
H. Sapiens or T-Rex are WRONG!! (Subtle hint: do not use the incorrect form on your homework or tests);
Higher order taxa are groups that contain genera. All taxon names other than species have one word names, which are capitalized; all taxon names other than genera and species are in roman letters (i.e., they are never italicized/underlined): Dinosauria, Tyrannosauridae, Animalia; not Dinosauria, tyrannosauridae, or animalia.
Traditionally, higher order taxa all had specific ranks: Kingdom, Phylum, Class, Order, Family. And as more diversity to life was found, these got subdivided and subdivided or grouped and grouped again, so we got Subclasses and Infraorders and Superfamilies and Tribes (and Subtribes!). But in the later 20th Century it was recognized that these formal ranks were actually meaningless!! An "order" doesn't mean anything in particular, and two "classes" aren't actually equivalent in any special way. So (as we'll see next week) we've abandoned these higher order ranks, but we STILL use the names of the taxa. Those DO represent real things: groups of living things.
Taxon names of whatever "rank" have some etymology (derivation) (that's true of all words, really). Sometimes the name might be descriptive (e.g., "Smilodon fatalis", the "fatal saber-tooth") or it might honor a place or geological unit of discovery (e.g., "Kayentachelys", a fossil turtle found in the Kayenta Formation of Arizona) or some individual (e.g., "Eldredgeops", after famed trilobite paleontologist and evolutionary biologist Niles Eldredge). But the name can be inaccurate (e.g., "Basilosaurus"--Emperor Reptile--is a whale, not a reptile!) but if the name was formed obeying the rules of taxonomy, that inaccurate descriptor is fine.
Type Specimens and Type Species: Another aspect of Linnaean taxonomy is that each species must have a particular type specimen. This is a particular individual preserved specimen (extant animal) or fossil (extinct animal) that is the "name holder" for that species. A type specimen is specifically referred to in the original description and diagnosis of the species. It need not be the most complete specimen known at the time (although that helps, as the more complete it is, the better the chance a less-complete individual can be compared to it!). The type specimen plus all the additional (referred specimens) are collectively called the hypodigm. Ultimately, if a species is regarded as being "valid" (that is, representing a real species in Nature), the type specimen is the only individual that is absolutely certain to belong that that species.
Similarly, each genus has a particular type species. This is the particular species to which the genus name is linked. If a genus is valid, the type species is the only species that is absolutely guaranteed to be within that genus.
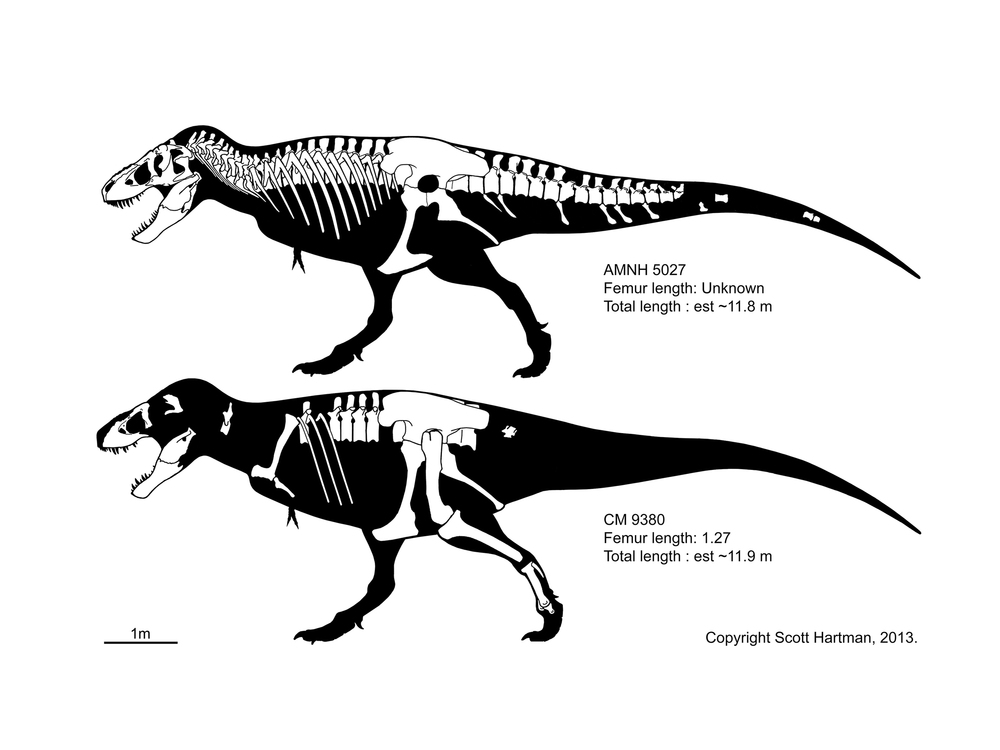
As an example, CM 9380 (in the collections of the Carnegie Museum of Natural History) is the type specimen of Tyrannosaurus rex, and Tyrannosaurus rex is the type species of the genus Tyrannosaurus.
Parataxonomies: There is a formal set of names for some types of fossils that is parallel to, but independent of, the biological nomenclature of actual species and genera and the like. These are parataxonomies. For instance, there is an "ootaxonomy" of "oospecies" and "oogenera" and "oofamilies" of fossil eggs, and a whole complex of ichnospecies for trace fossils. These are even given italicized Latinate names and use rules of priority and the like. But these are names of the eggs, burrows, footprints, etc., and NOT of the organisms that produce them.
Because there is disagreement about the features used to define a particular species or genus, different biologists and paleontologists will sometimes disagree about which specimens belong in a particular species, and which species belong in a particular genus (and so forth).
Taxonomists who consider a particular set of specimens to represent many taxa are called splitters; those who consider a particular set to represent few taxa are called lumpers;
If a taxonomist feels that some specimens of a genus belong to an as-yet unnamed species, they can split these specimens off as a new species (which a new type specimen);
On the other hand, if a taxonomist considers that two previously named species are not distinct enough from each other to truly be distinct species (that is, the taxonomist regards the two names as synonyms), they may lump them together:
In these cases, the Rule of Priority is used: whichever of the names was published first, even if only by days, is the name that must be used; the older of the two names is the senior synonym and the younger one(s) the junior synonym(s):
The same case applies to genera: if two genera are thought to represent the same genus, the first named genus name is the senior synonym, and thus has priority.
Here are some cases:
Splitting: the American Devonian species of trilobite Phacops rana (named by Green in 1832) was eventually regarded as being sufficiently different from the type species of Phacops (P. latifrons, named by Bronn in 1825) to belong to a new genus. No other genus name was already available for rana, so Struve proposed "Eldredgeops" in 1990. We now regard that American species as Eldredgeops rana (Green 1832) (the coiner's name is in parentheses to mark that this was not the original genus the species was in.)
Lumping: the Javan "apeman" was initially given the name Pithecanthropus erectus Dubois 1892. Subsequent work showed that it was not sufficiently different from our own species and related ones (like Neanderthals) to warrant its own genus name. Since we regard both the extinct Java species and our own as belonging to the same genus, and since Homo Linnaeus 1758 has priority over Pithecanthropus Dubois 1892, we now call the Javan form Homo erectus (Dubois 1892)
Sometimes, by accident, two taxa wind up with the same name. These are said to be homonyms. In this case, the senior (earlier proposed) of the two names occupies the name (i.e., it gets to keep it!). The junior homonym needs a new name: maybe there is another name already proposed that could be used, but if not it needs a new name. For instance, a dinosaur was given the name Syntarsus in 1969; unfortunately, a modern beetle was given that name back in 1869! So the beetle occupies Syntarsus, and the dinosaur wound up being renamed Megapnosaurus in 2001.
For those interested in a website concerning some unusual Linnaean species names, click here.
But, What ARE Species?
What is a species? Above we see the rules for these names, but it doesn't tell us about what it is being named.
Linnaeus' "species" were taxa like lions, tigers, black bears, etc. These were assemblages of individuals that share certain attributes:
Similar appearance
Similar habits and behaviors
Similar habitats
Darwin did not regard species as a distinct "kind" of biological entity. Instead, he considered them as essentially the same thing as geographic or stratigraphic variations (see these below), but ones in which extinction has removed the intermediate forms that otherwise would blend into the closest living relative group.
20th Century biologist Ernst Mayr (and most contemporary biologists) formalized their definition of a species as a "an array of populations which are actually or potentially interbreeding, and which are reproductively isolated from other such arrays under natural conditions". (Almost certainly you learned some version of this in high school and BSCI classes.) It works pretty well for the first pass: it emphasizes isolation, and thus species would represent pools of shared genes which do not get mixed with their closest relatives. Mayr and his followers refer to this as the "biological species concept" (or "BSC"), but as paleontologists Leigh Van Valen (see quote above) pointed out, this is an over-reach on the part of its proponents, and it is better termed a "reproductive species concept".
But there are some problems with this. For one: hybrids (crosses between two separate species) do occur naturally, and many of these are actually fertile! And for paleontologists: we can't test interfertility between populations because they are dead!
Other problems with the BSC exist, too:
Impossible to define for asexual species, unless each clone lineage is a separate species, despite being otherwise no different from each other than are two individuals are within sexually reproducing species
Extremely difficult to test in most wild populations, and (worst for paleontology) IMPOSSIBLE to test for fossils!
Existence of rare but real natural hybrids show that such isolation is not complete by any means
Existence of ring species shows that you can go through a continuum with no boundaries and yet reach conditions of genetic incompatibility between end members.
This is part of what is called the species problem: it is damned difficult to find a good, useful criterion that can be broadly applied to our ideas of what species are and how we recognize their boundaries. Biologists have tried to come up with hard-and-fast rules about how to recognize species, and although they have named many such "species concepts", these tend to cluster around two major different ideas:
Species are recognized by some shared aspect or attribute held in common by members of the species (similarity criterion)
Species are distinguished from other species by some attribute that makes them different from other (difference criterion)
Although related, they really aren't talking about the same things necessarily.
There are other species definitions and concepts that people have tried to apply, but none have been able to universally encapsulate the diversity out there.
There are thus problems with each of the distinct species definitions/concepts/criteria. None entirely encompass the range of biological possibilities, or are either too restrictive or too inclusive compared to the species as actually recognized and used by field naturalist and other working biologists. But just to add to the difficulty, here are some additional problematic issues:
Species originating within another species: This is a particular problem for the phylogenetic species concept. We recognize that some species (indeed, likely many!) originate from subpopulations of previously existing species; the REMAINING subpopulations are free to persist under the original morphology, range, habitat, gene exchange, etc. A notable example includes a number of extinct Hawaiian geese and the surviving Branta sandvicensis (nene), which represent a cluster of new species arising from isolated populations of Canada geese (Branta canadensis), the rest of which's populations remained in the original morphology and habitat. Similarly, polar bears (Ursus maritimus) arose as isolates within the cluster of populations of brown bear (Ursus arctos).
Introgression: This is a problem for the BSC. The rise of genomic analysis has revealed that not only is hybridization in the wild not a rare phenomenon, it does not (as some BSC advocates hoped and claimed) only produce rare individuals of reduced fitness, and thus do not contribute to the adaptive success of variants within the population. Instead, it has been discovered that introgression (the repeated backcross of hybrids with their parent species, resulting in significant gene transfer from one species to another) is actually disturbingly common. Indeed, it is appearing time and time again in many, many groups of animals (it had long been recognized to be common in plants.) In just the last several years it has been documented among many types of mammals, including various species of bears, of big cats, of mysticetes (baleen whales), of proboscideans (including the survival of genes from extinct elephantids in the surviving species), and in the genus Homo. Note that in all of these but whales the studies have included recent (Pleistocene) fossil DNA, showing this phenomenon is on-going. And in at least the human case, the Neanderthal genes introduced into some modern human populations code for traits with positive adaptive significance: that is to say, individuals that are descendants of these hybrids have the potential for increased selective success compared to other individuals.
Species originating by hybridization: This introgression pattern achieves greater expression in entire species which arise by hybridization between closely related taxa. Note: this is a distinct phenomenon from the hybridogens mentioned last time: those are species where every individual has to be produced by a hybrid breeding event. These cases were initially established by hybridization, but later contained populations of interbreeding individuals with distinct morphologies and genomes from either ancestral lineage. These cases remain controversial, but two examples that seem to be reasonably well-supported are: the red wolf (Canis rufus) of Southeastern North America, with ancestry in the grey wolf (C. lupus) and either (or BOTH) the eastern wolf (C. lycaeon) and the coyote (C. latrans); and the Jefferson mammoth (Mammuthus jeffersonii), a morphologically-distinct form genetically nested among the Columbian mammoth (M. columbi) but with a substantial amount of woolly mammoth (M. primigenius) genes.
So where does that leave us?
There really are clusters of biodiversity with shared history, interbreeding, ecology, etc.
In general (barring ring species, etc.), these variation within each cluster is less than the the distance to the next such cluster
At lower levels of analysis, reticulation (interbreeding) dominates inheritance over divergence (branching)
As you move to higher level branching tends to dominate, but (as we now know) reticulations do not cut off at some arbitrary point. (Even though they are essentially absent [so far as we know] above the old-style family level, reticulations are present by rare at very high levels in the form of various sorts of endosymbiosis, as in the origins of lichens, eukaryotes, plants, etc.)
So the word of advice is that at present no species concept/criterion is perfect, so hold off on accepting them as Holy Writ!
Keep in mind, the goal of Science is to describe Nature. Modify rules to fit Nature, not Nature to fit rules.
And specifically with regards to fossils: in the end--with the rare exception of fossil genomes--all fossil species are morphospecies, since we really can't see other aspects of them. And since there is the time factor that neontologists don't have to deal with, we see stratigraphic variation which segue into chronospecies. (We'll talk more about this issue and rates of change, later.)
In real life, species do seem to have "fuzzy boundaries", and the distinction between different closely related species on the one hand and clusters of variation within a species are nearly impossible to tell. In fact, biologists go through shifts of fashion towards increasing splitting (the former idea) and lumping (the latter) over time. Currently the fashion is towards splitting: consequently, whereas in much of the 20th Century we recognized only one species each of African elephant, gorilla, orangutan, Nile crocodile, and orca, early 21st Century taxonomists recognize two or more. (On the flip side, dinosaur paleontologists seem to be following the opposite trend, lumping once-separate species and genera into each other).
As with many things, we run into problem with typological thinking: the idea that there are ideal types of things, and that we judge a specimens membership in a group by how well it conforms from that type. Instead, we find that variation is the reality. So we need to use population-based thinking. (Next lecture we will add tree-based thinking.)
Ultimately, for paleontologists we are stuck looking only at shapes (and in fact, only the shapes of those hard parts that survive fossilization).
The question then becomes: how different do two individuals, or two populations, have to be for us to consider them different species? This is actually a terribly difficult question even with living organisms!! There are several sources of variation:
Sexual dimorphism: different sexes are different sizes and shapes and have different structures
Ontogenetic (growth): babies look different from juveniles look different from subadults look different from adults (can be even more
extreme in animals that undergo metamorphosis, like amphibians and many insects)
Geographic: populations in different regions might have slightly different sizes, color patterns, proportions, behaviors, etc. For example, some biologists consider the populations of orangutans, tigers, African elephants, etc. as distinct species; others simply regard them as regional variants
Stratigraphic: lineages (ancestor and descendant populations) may shift in some traits or characteristics over time
Individual: one of the great "discoveries" of Darwin and Wallace (we'll meet them next lecture), the recognition that no two individuals in a population are identical! (Before them, many people thought that there existed the perfect "type" of each kind of organism, and all variation is degeneration from that perfection. Darwin and Wallace showed that the variation is the reality)
Taphonomic: Not an issue that we have to deal with so much with modern species, but a great issue with fossil ones. We never have 100% of the organism preserved as fossils, and are lucky to get 100% of the hard tissues in complex organisms. So the difference that we see between two specimens may be due to differential quality or quantity of preservation rather than actual biological differences.
In fact, the recognition that species were NOT absolute kinds, but instead have "fuzzy" boundaries that blend into each other, is one of the main clues to the discovery of evolution.
Identification and Display Features
Many animals engage in various types of display:
Sexual (aka courtship) displays: attract potential mates
Territorial displays: defend territory (which might also include mates)
Defensive display: more generally, to warn off potential threats
Specific Recognition: features distinctive to each species. Especially common where multiple closely-related forms live in the same
environment
Some of these forms of display result in preservable "showy" features of the anatomy that might help us identify species more easily, or ironically confound us into thinking that two different sexes represent two different species!
Sexual strategies: male and female animals have different priorities in terms of reproduction. Males can in principle fertilize many many individuals, while females typically have fewer sex cells (eggs) available at any given time. With less cells to use, females often are "choosier" in terms of mates. So many species evolve displays in which males somehow "show off" (in terms of physical features, ritual motions, combat between
rivals, etc.) and females evaluate the display.
For example:
Sexual Dimorphism: when the two sexes (at least as adults) have distinctive forms. Difficulty in testing this in the fossil record:
Are you looking at fossils of two sexes in the same species, or two different species?
Many sexual display features (and sex organs!) are soft tissue, so they would only rarely fossilize
If you have only a few individuals, are you looking at true dimorphism, or just end members in a continuous spectrum?
Some things to look for in potential cases of sexual dimorphism:
One morph (often the male) is rarer
One morph (often the male) is showier
Distinction between alleged males and females is less pronounced prior to sexual maturity
Changes to dimorphic forms is rapid once sexual maturity is reached
Frequency of the two morphs remains relatively constant in strata where the species is found.
In very rare cases the eggs or embryos have been found inside a fossil,
which rather unambiguously shows them to be female. Otherwise, there can be circumstantial evidence. For instance, if the species has crests, horns, etc., and these are some rarer showier crests, these might more likely be male.
An alternative to sexual displays for showy structures, however, is specific recognition systems (SRS). In this cases, different species have unique characteristics within their ecosystem to recognize other members of the species from all other species they encounter. For cases of olfactory and aural SRS we are lost with regards to fossils. But we have potential with visual SRS.
Things to look for in potential SRS:
Likely to be most pronounced when related species are sympatric (since they live together, there is greater need to distinguish A from B)
Differences should be in obvious traits, not subtle internal ones
Need NOT be sexually dimorphic!
Identification and Ontogeny
Another potentially confounding issue is ontogeny. As organisms grow they might look profoundly different. Some undergo varying degrees of metamorphism: consider tadpoles vs. frogs, or caterpillars vs. butterflies. Such examples also occur in the fossil record: without sufficient information and samples of the various growth stages in the proper sedimentological context, we might mistake different growth stages as different species.
But it need not be so profound a change to be an issue. For some fossil species there remain debates over whether smaller individuals of a particular group from a formation are the juveniles of the larger species, or are smaller species that lived sympatrically (in the same time and place). Without a large enough sample size and sufficient numbers of intermediate stages, this might be very difficult to resolve.




{kind=link}
{kind=link}
{kind=link}