Key Points:
•Whereas Chelicerata has long been known, Mandibulata has only been recognized within the last two decades thanks to the combined application of molecular and morphological phylogenetic methods.
•The mandibulate body has a head and a trunk whose segments may be divided into a thorax and abdomen or not.
•The mandibulate head appendages form one or two pairs of antennae, mandibles, and two pairs of maxillae.
•Mandibulata contains Myriopoda (centipedes and millipedes) and Pancrustacea (traditional crustaceans and Hexapoda.)
•Stem mandibulates include the Burgess Shale taxon Marella and other small and poorly known taxa.
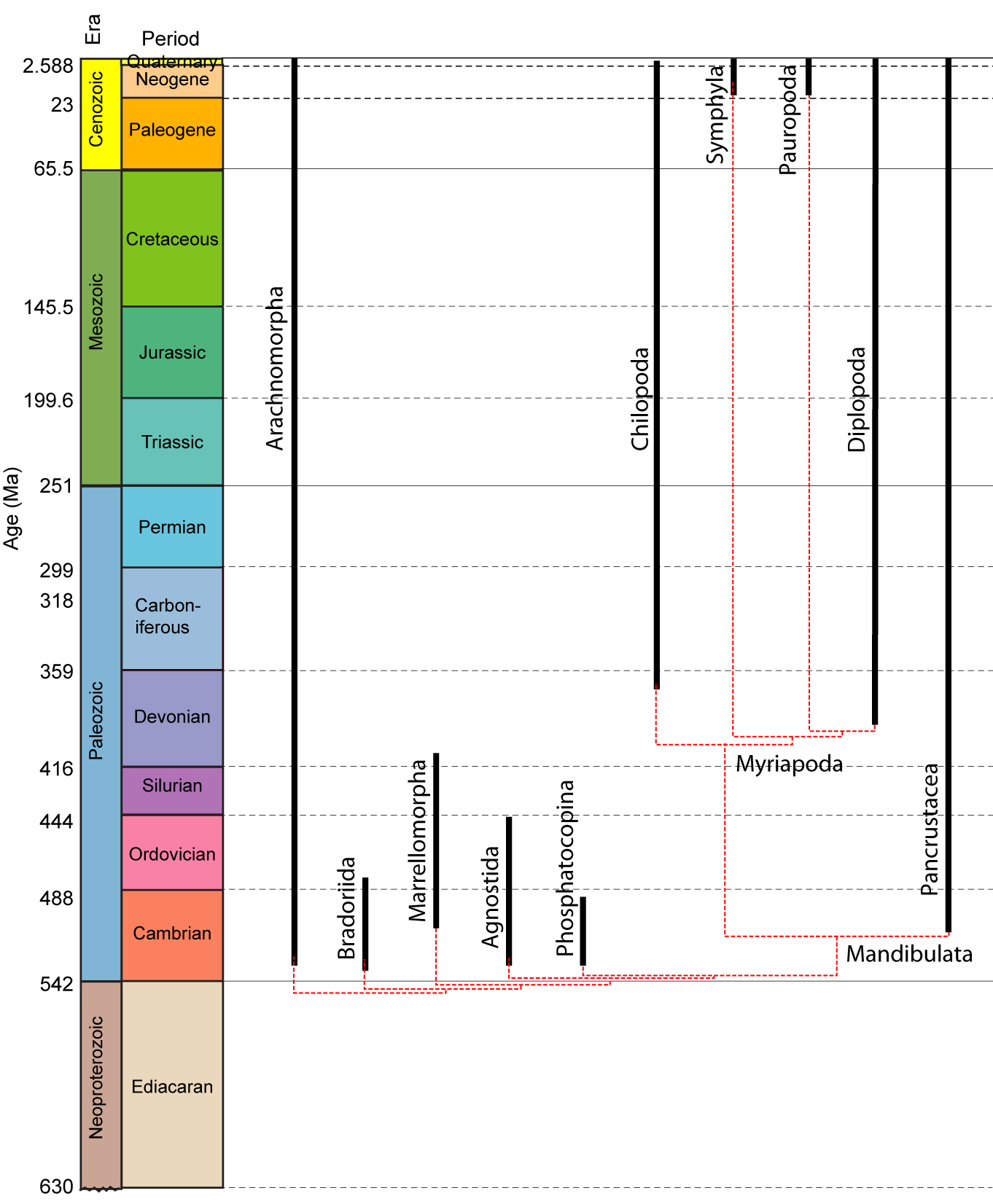
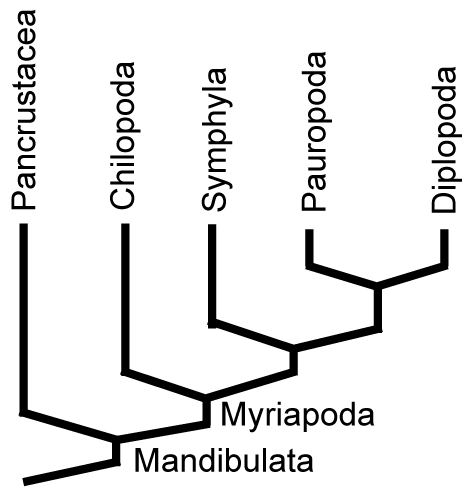
•Myriapoda contains Diplopoda (millipedes), Chilopoda (centipedes), Symphyla, and Pauropoda, each with unique patterns of tagmosis. Their synapomorphies include the exclusive use of a tracheal system for breathing and the presence of repugnatorial glands for the secretion of noxious chemicals.
•Chilopoda is present on land by the end of the Silurian, making them among the earliest land animals. In the form of Arthropleurida, diplopodans achieved their ecological zenith during the Carboniferous.
•A full inventory of Pancrustacea is impossible here. Diverse marine taxa with invasions of the land by isopods and Hexapoda.
•Pancrustacean phylogeny is unsettled, but by any account, Hexapoda is nested in its crown.
•Ancestral hexapodos are soil-dwellers. More derived forms are, in sequence, larger, possess wings, can fold their wings, and ultimately are holometabolous.
•The Late Devonian and Early Carboniferous comprise the Hexapod Gap, and interval when hexapods fossils are vanishingly rare. This corresponds to a similar gap for vertebrates - Romer's Gap.
•The Arthropod Head Problem, the struggle to understand the homologies of arthropod head appendages, seems to have been solved by the discovery of fossil stem arthropods and radiodonts from Chengjiang and the Burgess Shale that preserve patterns of innervation of head appendages.
"Working out the evolutionary stages by which all these combinations could have appeared is so difficult that has long been known as 'the arthropod head problem.' In 1960 R. E. Snodgrass even hoped it would not be solved, as trying to work out solutions was so much fun"
(Imran Ali Sani and Shahjahan Shabbir Ahmed, 2015.)
Mandibulata
The remaining arthropod diversity falls into Mandibulata, whose living members include:
Myriapoda (centipedes, millipedes, and kin)
Pancrustacea (traditional "crustaceans" plus insects and their kin)
Evolution of the concept:
The phylogenetic hypothesis of Mandibulata reflects phylogenetic work of the last decade by both molecular (E.G. Regier et al. 2010.) and morphological (E.G. Legg et al. 2013.) workers. It differs from the traditional taxonomy that still appears in many texts, where Hexapoda and Myriapoda form Uniramia - the arthropods without biramous appendages. Traditionally, there was no conviction that Uniramia and Crustacea were particularly closely related. Key aspects of the emerging consensus:
Myriapods, hexapods, and "crustaceans" are found to form a monophyletic Mandibulata (whose monophyly is well supported by molecular, developmental, and morphological analyses. (E.G. Mouthparts are derived from three sets of limbs whose homology are demonstrated by morphological similarity and gene expression.))
Hexapoda (but not Myriapoda) is found to be nested within Crustacea, rendering traditional Crustacea paraphyletic and sinking Uniramia completely.
A head with several pairs of specialized mouthpart appendages.
A trunk that may display homonomous segmentation (Myriopoda) or be divided into:
Thorax with walking appendages
Abdomen with swimming appendages or lacking appendages
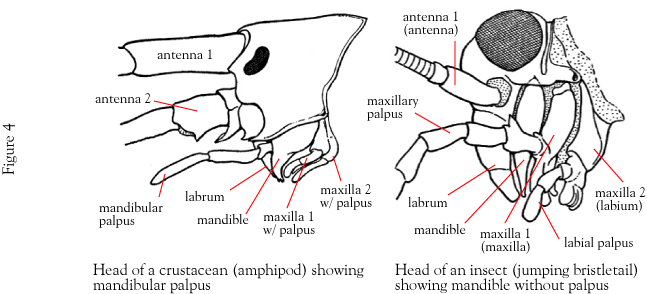
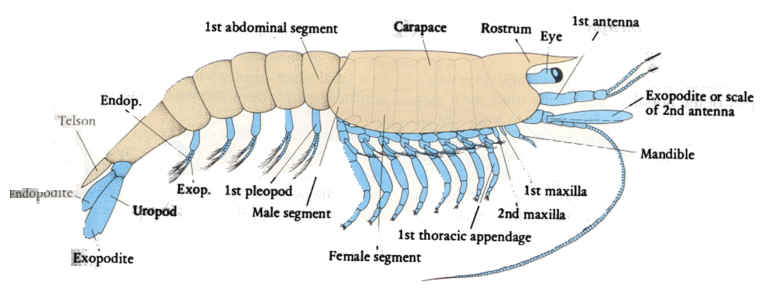
About mandibulatan head appendages: Whereas the equivalence of head appendages in trilobites and chelicerates is straightforward, those of Mandibulate are complex. We see, from front to back:
Second antennae: In crustaceans only, a second pair of antennae
Labrum (sing. labra - right): A midline structure that functions as an "upper-lip."
Mandibles: For shearing, piercing, and processing food.
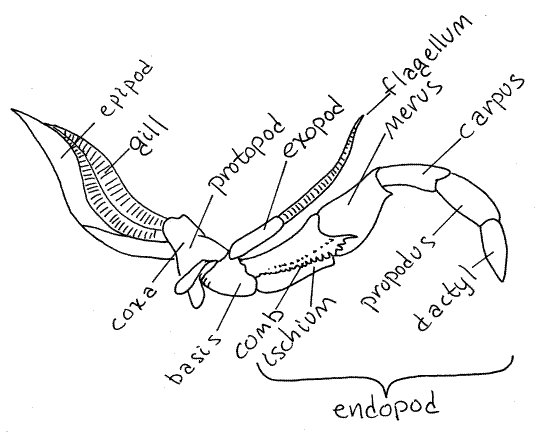
Maxillae: Ancestral biramous appendages for manipulating and tasting food. The slender exopodite is called a maxillary palp (in aquatic crustaceans, the mandible also bears a palp.) Ancestrally, first and second maxillae were present.
Maxillae can be fused or modified into derived structures. In the example of the insect (right) the second maxillae are fused to form a labium that functions like lower lip.
This katydid posed with its mouthparts for me.
All head appendages are readily visible in the katydid at right.
Recall that Mandibulata encompasses the living arthropods characterized by mandibles and two pairs of maxillae. This group encompasses over 75% of living animal diversity. With that much diversity, it is a wonder that with the application of phylogenetic techniques and the illumination offered by fossils, we understand its evolutionary history as well as we do. In this presentation we follow the results of Legg et al. 2013.
Living Mandibulate Diversity:
Falls into two major groups:
Scolopendra galapagoensis
Myriapoda: (Sil. - Rec.) Including
Diplopoda - millipedes (right)
Chilopoda - centipedes
Sally lightfoot - Grapsus grapsus
Pancrustacea: (C. - Rec.) Including
Hexapoda - Insects and their kin
A bewildering variety of traditional "crustaceans" (paraphyletic)
Stem Mandibulata: Mandibulata seems to have originated during the Cambrian, with organisms on the mandibulate stem occurring in the earliest Cambrian. Stem mandibulates include:
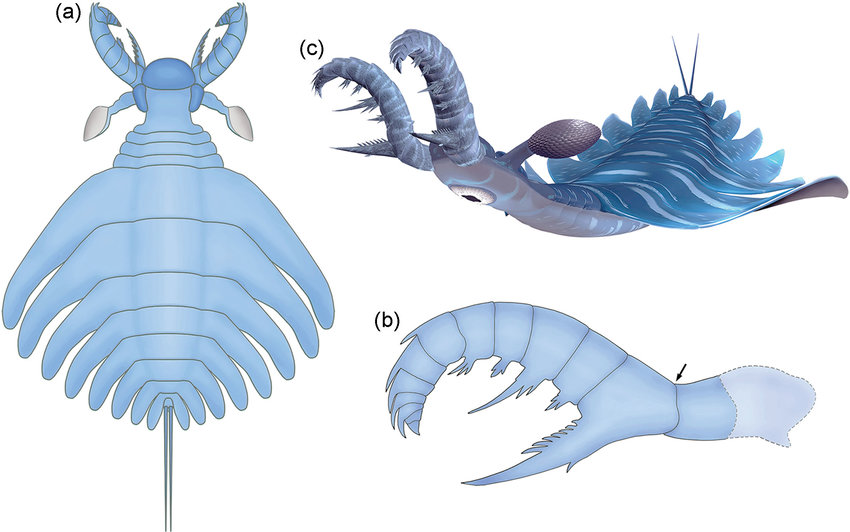
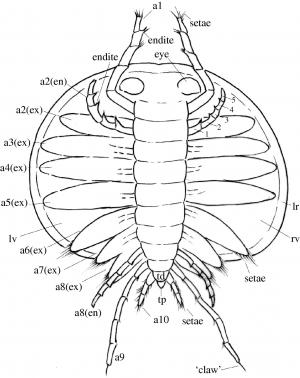
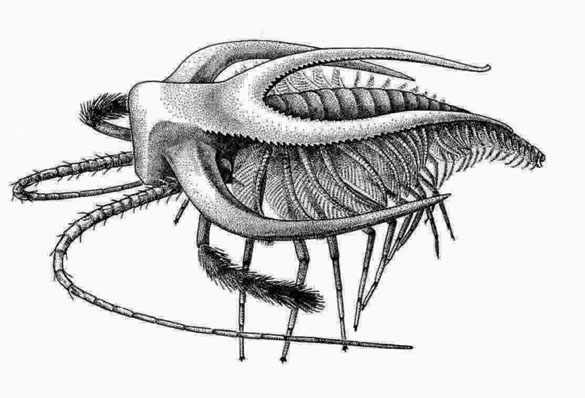
Bradoriida: (Cam. - Early Ord.) Tiny planktonic bivalve arthropods. (Do not confuse with bivalve stem-arthropods or with ostrocods.) Anterior two appendages may be homologs of antennae 1 and 2.
Marellamorpha: (Cam. - Early Dev.) The classic Burgess Shale fossil Marrella splendens is representative of a group that persists into the early Devonian. It's trunk appendages are plesiomorphic biramous, and unspecialized, but the first two may be homologs of antennae 1 and 2.
"Orsten taxa:" (Cam.) The analysis of Legg et al. 2013. recovered a paraphyletic group of taxa from the Orsten Konzervatlagerstätte. One interesting surprise is the presence in this group of the tiny bivalve "trilobite" Agnostus. This adds fuel to the discussion of whether agnostids are, after all, trilobites.
Phosphatocopina (Cam.) Yet another group of planktonic bivalve arthropods. Long considered to be ostracods, they are distinguised by details of limb morphology.
Note: All of the tiny bivalve taxa discussed above, along with living analogs, seem to develop from larvae possessing head appendages only, then to add segments and appendages posteriorly with each molt. This gives us confidence that their anterior head appendages are antennae. For our purposes, the presence of these appendages is a synapomorphy of total-group Mandibulata.
Myriapoda: (Sil. - Rec.)
Mandibulate arthropods with homonomous trunks (i.e. with repeating unspecialized modules). Whether they represent the first fully terrestrial animals depends on the resolution of claims that some early scorpions were actually aquatic. They were major components of the Silurian and Devonian land faunas, achieving large size as predators and as detritivores and grazers.
Potential synapomorphies:
Each segment bears paired repugnatorial glands that secrete noxious chemicals.
Breathing is with a tracheal system
The second antennae are lost (convergent with Hexapoda.)
Diversity Four monophyletic groups are known:
Symphyla: (Neog. - Rec.) Tiny eyeless inhabitants of soil and leaf-litter
Pauropoda: (Neog. - Rec.) Tiny inhabitants of soil and leaf-litter. Each trunk tergite covers two body segments.
Most recent phylogenetic analyses (E.G.: Shear and Edgecomb, 2010) reveal the phylogeny at right. Symphyla and Pauropoda have no fossil record except in Neogene amber, so we focus on the other two groups.
First maxillae united to form a "lower lip" (the gnathochilarium.)
The following body segment which would bear a second maxilla in most arthropods has no appendages.
Body segments: In millipedes, adjacent segments are fused developmentally so that each segment has two pairs of limbs, spiracles, etc. (An extreme development of the pattern in Pauropoda in which a single tergite covers pairs of segments. - Synapomorphy!)
Fossil diplopodans:
Trace fossils that could be made by myriapods extend to the Ordovician.
Arthropleuridea: (Sil. - Perm.) Long known as giant myriapods, arthropleurids have recently been shown to belong within Diplopoda. Their heyday was the Carboniferous - the source of their best fossils and trackways. Their fortunes waned during the early Permian along with the coal swamp environment they favored. Characteristics:
Big - up to 2 m. length
With specialized lateral lobes on their tergites, and limb specializations.
Oddly, although known from good fossils, there are no traces of a tracheal system. Could they have been semi-aquatic?
Chilopoda: (centipedes; primarily predators. Sil. - Rec.) Despite having a homonomous trunk, centipedes achieve a degree of biomechanical diversity, with some specialized for fast running and compact bodies (Scutigera right) and others being more serpentine.
Tagmosis: Centipedes have a single pairs of limbs, spiracles, etc. per body segment (plesiomorphic). Head appendages are plesiomorphic for Myriapoda:
Antennae
mandibles
first maxillae
second maxillae
Chilopod synapomorphy: The first trunk limbs are modified as poison-injecting forcipules and co-opted into the feeding apparatus.
Chilopod fossil record:
Body fossils of centipedes known from the latest Silurian. (See Edgecomb and Giribet, 2007 for a review of centipede evolution.)
Myriapod mystery: Given that Pancrustacea and Arachnomorpha were ancestrally marine, we would expect the earliest myriapods to be marine also. Although definite marine myriapods elude us, we are closing in:
Budd et al., 2001 report on a possible Upper Cambrian form, Xanthomyria spinosa , from Siberia. Inconclusive, but consistent with what we might expect.
Edgecombe et al., 2020 report that Heterocrania rhyniensis (right), a Devonian member of the obscure group Euthycarcinoidea (Cambrian - Triassic) displays probably synapomorphies with Myriapoda, suggesting a divergence age in the Cambrian.
Pancrustacea:
Pancrustacea (equivalent to "Tetraconata" of some authors) contains all traditional crustaceans and hexapods. Work of the last decade has produced the growing consensus that "crustacea" is paraphyetic with respect to Hexapoda, but there is no clear consensus of exactly where, within Pancrustacea, hexapods go. For now, we follow the morphology-based analysis of Legg et al. 2013., but interested students should also review the molecular result of Regier et al., 2010.
Synapomorphies of Pancrustacea, involved soft tissue characters of the brain and the shape of ommatidia, and exceed the technical limits of this course. Indeed, the whole topic is so complex that we can only scratch the surface. Beyond possessing the synapomorphies of Mandibulata, Pancrustacea displays:
Development from nauplius larva in which the anterior head appendages form first.
Small: Typically around 2 mm. Largest living species up to 30 mm. Largest fossil on record was 80 mm.
Lives within an expanded bivalve carapace. This is homologous to the carapace of other crustaceans, but with a mid-line hinge, allowing to to be closed over the rest of the animal.
The posterior body is greatly reduced.
Seven pairs of appendages are present, of which the first five are the antennae and mouthparts. These do double-duty as swimming organs.
Because they evolve rapidly, are widespread, and their valves are robust fossils, ostracods are classic index fossils.
Until recently, Ostracoda and Thecostraca (the barnacles) would have been united in the clade Maxillopoda however recent analyses do not find this to be so.
Tetraclita milleporosa
Thecostraca (Barnacles (Cirripedia) and their kin.) (Cam. - Rec.)
Characteristics:
Begins life normally enough, as yet another small crustacean with bivalve carapace. Soon attaches by the head to hard substrate, sheds carapace and secretes a set of distinct calcareous plates that are non-homologous to those of other arthropods.
Thoracic limbs modified to form suspension-feeding cirri.
Two major groups, each with its own pattern of adult plates:
Lepadomoprha: Goose-neck barnacles. The adult plates are non-homologous with those of balanomorphs (?) and the creature is anchored to the substrate by means of a fleshy peduncle.
Because they can form, in essence, barnacle "reefs" they have a very good record. The earliest putative barnacle: Collins and Rudkin, 1981 report on Priscansermarinus barnetti - a possible lepadomorph from the Cambrian Burgess Shale.
Branchiopoda ("sea-monkeys," water fleas, and clam shrimp) (Poor record - Cam. Rec.)
Characteristics:
Small, <10 mm.
Variable number of thoracic limbs
Basal section of thoracic limbs developed as paddles that also serve as gills. Hence the name "gill-footed." (Even though the typical exopodite ramus is suppressed.)
Fossil record: Waloszek, 1993 describes the earliest known branchiopod, Rehbachiella from the Late Cambrian of Orsten. Although originally marine, branchiopods today mostly occur in fresh-water. Indeed, brine shrimp (sea monkeys) have developed a life cycle in which eggs are able to withstand dessication for long intervals. Thus, new generations can repeatedly colonize ephemeral pools of water.
Cephalocarida (No fossil record): Horseshoe shrimp. A minor group with only twelve species. Strictly shallow-marine inhabitants of intertidal zone. Morphologically simplified, with no pleopods, and second antenna shifted posterior to mouth in adults.
Pandalus platyceros
Malacostraca (Cam. - Rec): The more familiar crustaceans.
Remipedia: (Carb. - Rec.), Discovered in 1981, these morphologically, are the most primitive looking crustacean group, with homonomous bodies, but for the characteristic crustacean tagmosis of cephalic appendages. Most live in submerged caves and places of permanent shade in brackish-saline aquifers. Thus their eyes are (presumably) secondarily reduced. Characteristics:
All post-cephalic appendages derived for swimming.
Remipedes have morphologically adapted for swimming on their backs.
Remipede mouthparts are raptorial and seem to be modified for transmitting venom, but we don't really know how they feed.
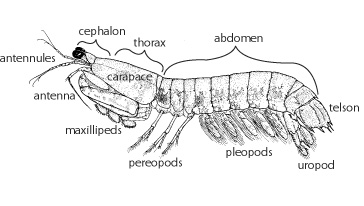
Like (other) crustaceans, the insect body is tagmatized into a cephalon, thorax, and abdomen
See below for more information.
But there are significant differences. Alas, insect cuticle is not calcified. Thus, their fossil record is poor compared to that of crustaceans (though not as bad as you might think). What there is comes largely from Konzervat-Lagerstätten
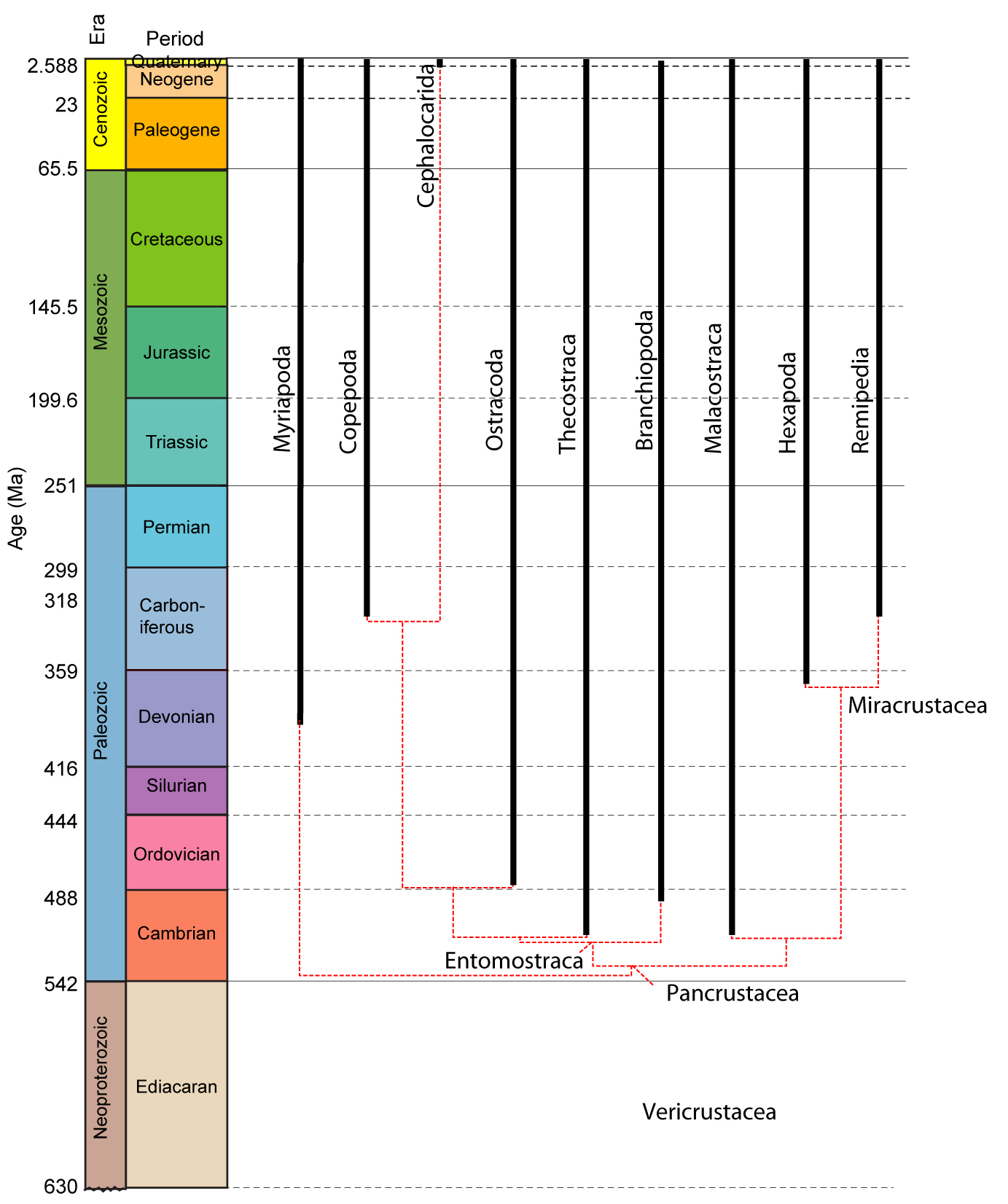
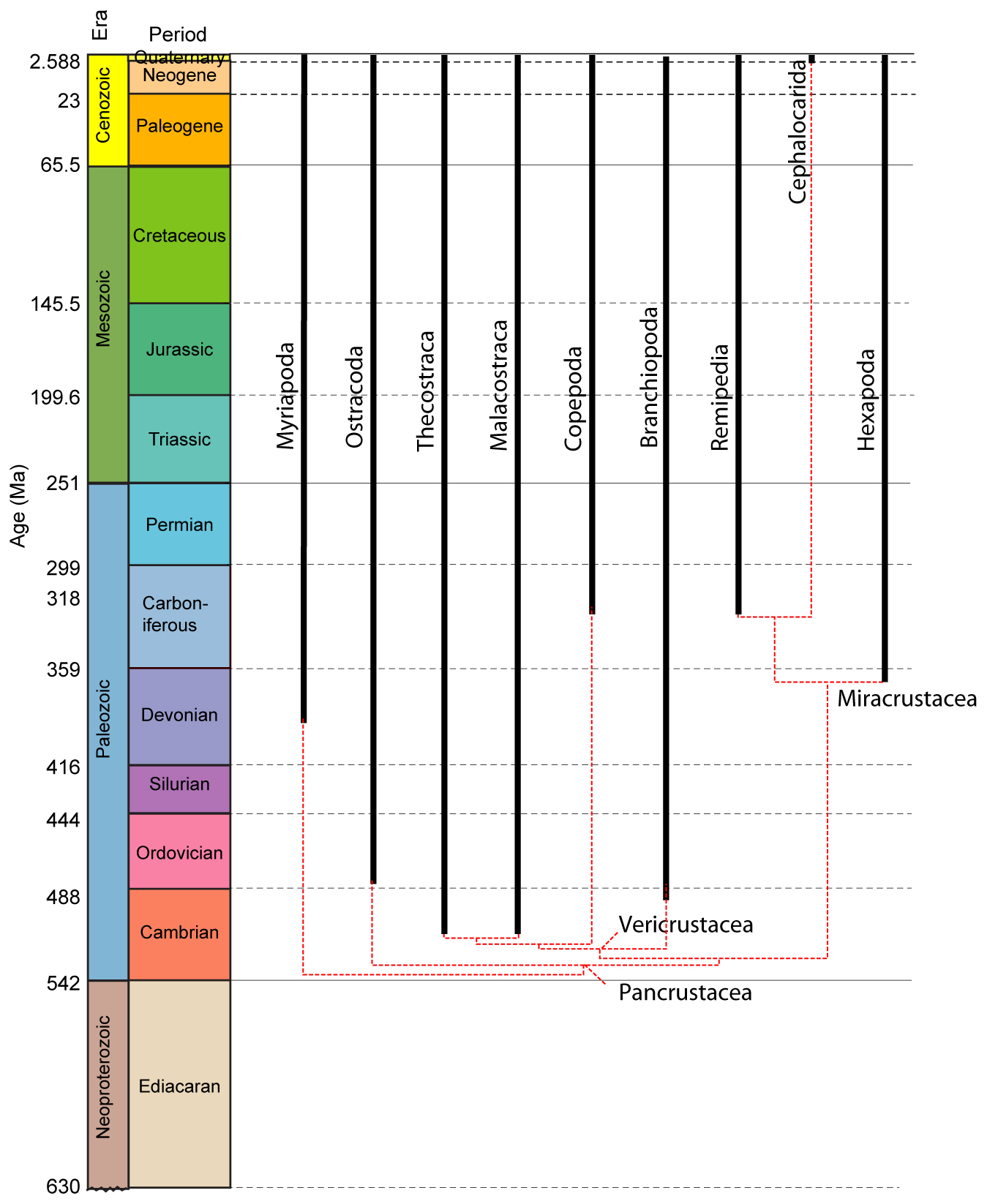
Pancrustacean Phylogeny:
The current state of the art is unsettled, with large areas of disagreement between molecular and morphological results:
Morphological: Legg et al. 2013.. Note, the synopses of their result above omits many fossil taxa. Interested students should consult the publications.
Points of Agreement:
Miracrustacea: Remipedia and Hexapoda are closely related. Seems odd, but each group has fossil relatives that, like Hexapoda, have replaced the second antenna with an appendage-free intercalary segment. Of course, this makes traditional "Crustacea" paraphyletic.
Little guys: Thecostraca, Copepoda, and Branchiopoda are closely related in both results. Legg et al. refer to this as Entomostraca.
Significant Points of Disagreement:
Ostracoda: Either ostracods are the sister taxon to all other pancrustaceans, or they are nested within Entomostraca with the other little guys.
Cephalocarida: Members of Miracrustacea or Entomostraca?
Malacostraca: Are they closer to Miracrustacea or nested among the little guys? The molecular result supports a pleasing dichotomy between pancrustaceans closer to insects and those closer to the major marine groups (Vericrustacea) The morphological result is much more stratigraphically congruent.
The common ancestor was probably a shrimp-like creature.
Eight pairs of thoracic appendages broken down in various ways in different groups.
Hoplocarid Odontodactylus scyllarus from Wikipedia
Major malacostracan groups:
Phyllocarida: (Cam. - Rec.) Large bivalve carapace. Yet another entry in the bivalve arthropod ecospace.
"Eocarida" (Dev. - Per.) Shrimp-like crustaceans of Late Paleozoic coal swamps. (E.g. Angustidontus seriatus From Late Devonian - right.)
Hoplocarida: (Right) (Dev. - Rec.) Enlarged raptorial first walking limbs. Living members comprise Stomatopoda. Capable of fastest measured movements by any animal. Large eyes. Fossil record begins in the Devonian (Jenner et al., 1998).
Syncarida (Carb. - Rec.) Lacking carapace, thus revealing segmented thorax with biramous appendages. Thought to have gone extinct in Permian until living specimens found in Tasmania. (E.g. Acanthotelson stimpsoni from Mazon Creek.)
Peracarida: Include marine and terrestrial isopods - "pill-bugs."
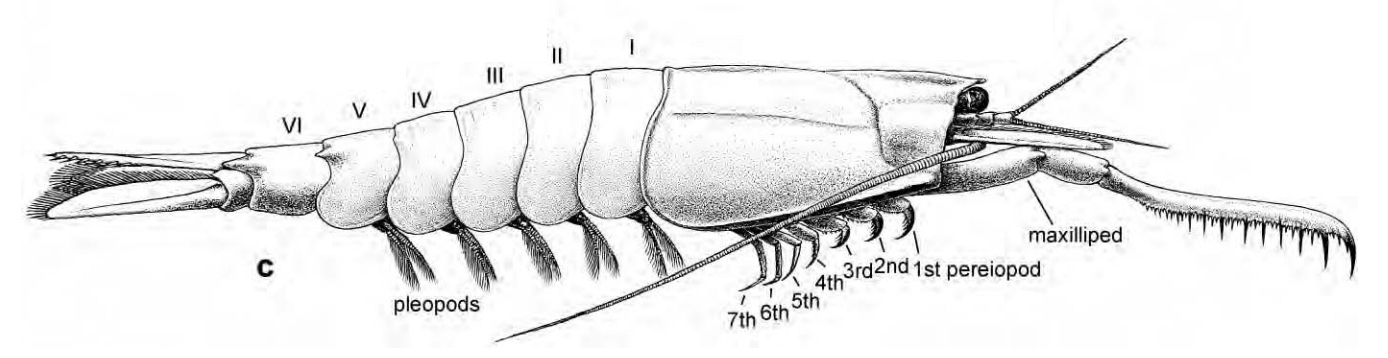
Decapoda: (Dev. - Rec.) Includes any crustacean you have ever considered eating.
Characteristics:
Body has 20 segments, six for the head, eight for the thorax, and six for the abdomen:
Thoracic limbs divided into:
three pairs of maxillipeds for food manipulation
five pairs of walking limbs. Note: these remain biramous, even though the gills are typically enclosed by the carapace.
up to three anterior pairs of walking limbs may bear chelae (lobster pincers are robust examples)
Occupy many habitats, from deep ocean to fresh water and land (almost)
A rogue's gallery of interesting decapods:
Brachyura: Crabs - Abdomen reduced and flattened. In adult, it is folded beneath the thorax, offering protection from dessication and facilitating movement. Delicious!
The weird part: Hexapoda (Dev. - Rec.): Proper insects and their close relatives. Closely related to Remipedia in current results. Ancestrally each group is adapted to life underground, the remipedes in saline aquifers, the hexapods in moist soils.
Alas, insect cuticle is not calcified. Thus, their fossil record is poor compared to that of crustaceans (though not as bad as you might think). What there is comes largely from Konzervat-Lagerstätten
Some have a median abdominal filament (silverfish, mayflies) that represents the telson.
Ventilation is exclusively by means of a tracheal system, in which tracheae take in air through spiracles and deliver it directly to the tissues. The tracheae are lined with cuticle that is shed during ecdysis. Note that by this means, insects are able to achieve aerobic scopes approaching 300 (compare to 10 for mammals) (E.G. Hoverflies mate while hovering. Can YOU do that?).
These are distinct from basal hexapods in being larger and spending time on the surface. Insecta is diagnosed by a list of technical anatomical synapomorphies.
Doomed adult mayfly
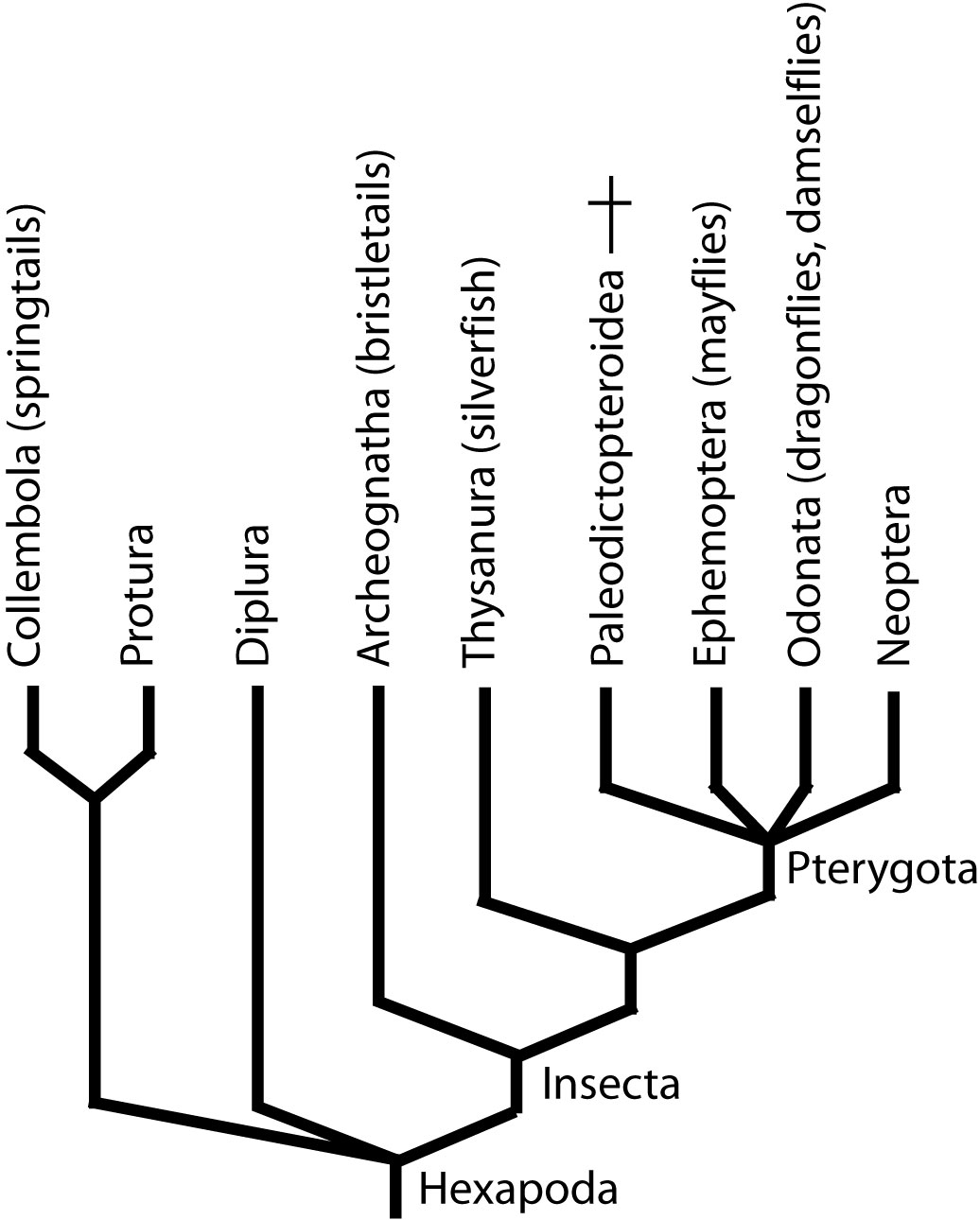
Within Insecta is Pterygota,
the winged insects (first organisms to slip the surly bonds of Earth). In most, a pair of thoracic wings (arguably derived from the exopodite ramus) is matched with the posterior two limb pairs. In some extinct forms, there was a third small pair anteriorly, making a wing pair corresponding to each leg pair. The evolutionary origin of wings is enigmatic, but a leading hypothesis holds that they are homologous to the dorsal ramus of ancestral biramous appendages.
Pterygota incudes:
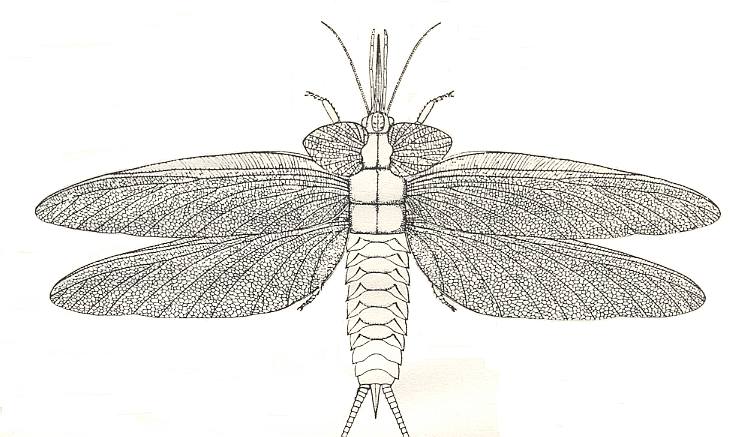
Paleodictyoptera: (Carb - Perm) Extinct group of large bodied insects (largest with 50 cm. wingspans.) with three wing pairs and piercing/sucking mouthparts. Possibly paraphyletic.
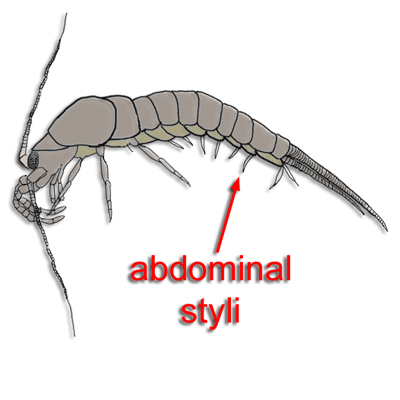
Ephemoptera: (Mayflies, Carb. - Rec.) Spend most of life as fresh-water aquatic opportunistic feeders. They breathe using gills derived from abdominal styli (I.e. the plesiomorphic arthropod pattern.) Mayflies metamorphose into winged adults for a brief life of mating. As adults (right), living members have no functional mouthparts. This is apomorphic. Indeed, Permian fossil mayflies appear to have had proper mouthparts (Prokop and Nei, 2011).
Tramea cophysa
Odonata: (Dragonflies and damselflies. Perm.-Rec.): Extremely skillful fliers and aerial predators. Spend their youth as aquatic predators (using specialized protrusible labium to snare prey), then metamorphose into winged adults. Unlike mayflies, they spend months as adults feeding and mating. Some odd things: It is tempting to assume that odonatans ancestrally had life cycles like mayflies, and yet:
Although fossil adult odonatans were around in the Pennsylvanian, we find no fossils of their aquatic nymphs until the Jurassic. (Indeed, fossils of crown-group Odonata occur only back to the Jurassic. Stem-group odonatans occur as early as the Carboniferous)
The most phylogenetically basal odonatans, the Petaluridae, lay eggs and spend their youth in moist soil and leaf litter, not in bodies of water.
Different structures perform the function of gills in different odonatan groups.
Is it possible that the ancestral odonatan had no aquatic stage?
The odonatan stem group - Protodonata (Carb. - Tri.) (paraphyletic) included creatures like Meganeura with wingspans up to 75 cm.
Note: before considering the last pterygote group, consider what the life cycle of these "primitive pterygotes" tells us: Unlike apterygotes, pterygotes undergo a partial metamorphosis from and aquatic nymph to a terrestrial imago (adult).
Evolved a hinged articulation by which the wings could be folded over the back. An obvious advantage if, (unlike dragonflies) you planned to spend time on the ground. This made possible a striking radiation of flying/walking insects. The real breakthrough came when Coleoptera (beetles) transformed their forewings into elytrae (wing covers) to protect the hindwings while on the ground. This made possible behaviors like burrowing and the invasion of microhabitats that might otherwise damage wings. Thus, beetles make up roughly 50% of animal diversity.
While all arthropods experience discrete instars (growth stages separated by episodes of ecdysis), many Neopterans are holometabolous undergoing a complete metamorphosis between totally dissimilar life stages:
The earliest known hexapod is Rhyniella praecursor, a collembolan (springtail) from the Early Devonian Rhynie Chert Konzervat-Lagerstätte.
Winged insects appear abundantly in the Late Carboniferous. The interval from the Middle Devonian through the Early Carboniferous is called the hexapod gap, in which hexapod fossils are unknown. This, interestingly, encompasses Romer's gap, an analogous gap in the land vertebrate record in the Early Carboniferous. (More on this later.)
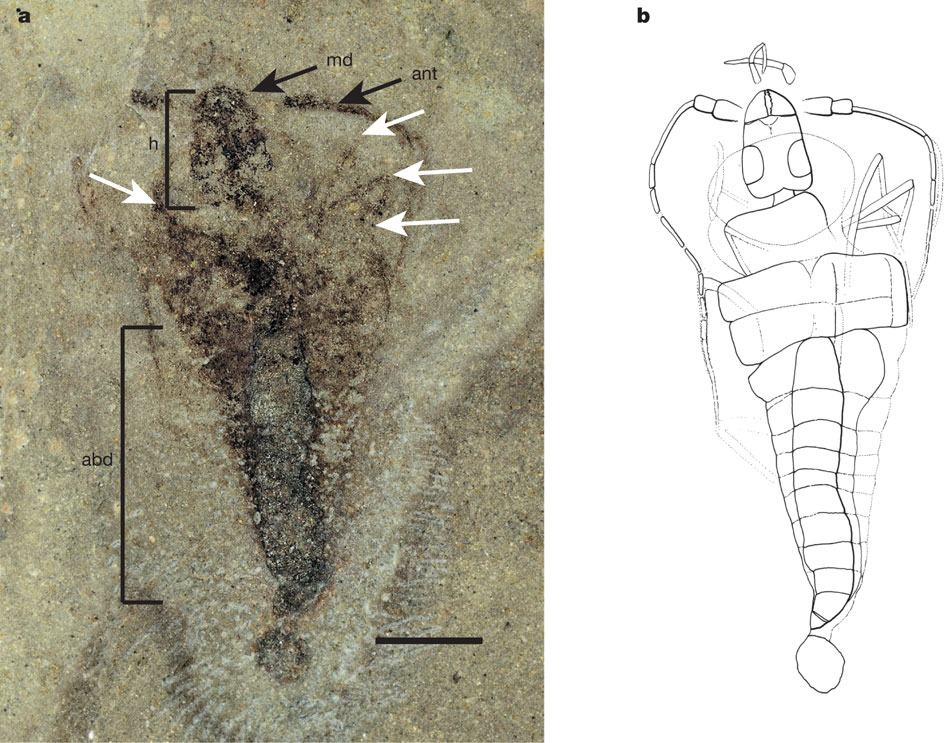
But wait! Garrouste et al., 2012 report on Strudiella devonica (right) from the latest Devonian, smack in the middle of the hexapod gap. Although wingless, its jaws resemble those of winged insects. Is it an adult or the nymph of a winged form?
Outstanding issues:
What allowed Carboniferous - Permian land arthropods to get bigger than later ones? (E.G. dragonflies like Meganeura with 75 cm wingspans, arthropleurid millipedes.) (E.G. Absence of flying vertebrates? Higher [O2]?) Arthropleurids seem to have tracked[O2] faithfully, vanishing in the early Permian. Paleodictyopteroids and protodonatans were out by the Early Triassic. Remaining odonatans had been reduced to current size range. Did the rise of flying vertebrates play any role?
What selective pressure facilitated the origin of flight? (Exaptation of gills or thermoregulatory structure? Exaptation of structure for surface tension locomotion?)
Dissertation, anyone?
Insect brain schematic.
The Arthropod Head Problem and the Power of Paleontology:
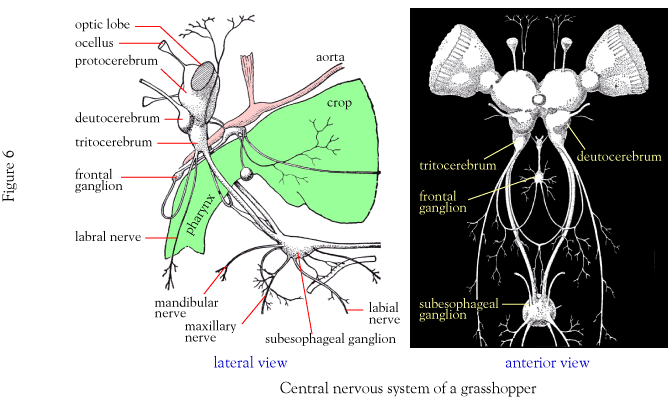
Questions about the homologies of various arthropod head appendages have vexed generations of zoologists. Today, they are finally yielding to the application of developmental, genomic, and paleontological data. First a review. The arthropod brain forms from the integration of the front three pairs of ganglia. Our first tool in homology assessment is which cerebral ganglion an appendage is innervated from. Terminology:
Protocerebrum the first ganglion pair
Deuterocerebrum the second
Tritocerebrum the third
In Euarthropods, we see these patterns of innervation:
Protocerebral: The eyes
Deuterocerebral: The antennae (in mandibulates) and/or chelicerae (in chelicerates)
Tritocerebral: The remaining mouth parts (in mandibulates) and/or pedipalps (in chelicerates), and the second antennae of crustaceans
Combining this information with informed hypotheses of phylogeny, it is becoming clear that:
The first antennae of just about all arthropods are homologous to the chelicerae of chelicerates
Chengjiang: During the last decade, gloriously preserved arthropods from Chengjiang that show the nervous system have enabled us to see that this arrangement has very deep roots. For example:
"Great appendages" of the "megacheiran" Alalcomenaeus are deuterocerebral and likely homologs to antennae and chelicerae (Tanaka et al., 2013).
Now, from the magic hats of Chengjiang and the Burgess Shale, emerge the radiodonts that offer conflicting interpretations:
Lyrarapax unguispinus (Cong et al. 2014). It, too, preserves traces of nervous tissue appearing to show that its raptorial appendages are also protocerebral, and are likely homologs to the antennae of onychophorans.
But remember the Burgess Shale radiodont Stanleycaris hirpex from Moysiuk and Caron, 2022. It's great appendage seems to be deuterocerebral - a possible homolog to mandibulate first antennae, chelicerae, and onychophoran mandibles. Also, Stanleycaris seems to possess a protocerebrum and deuterocerebrum, but no tritocerebrum. That, apparently, arose in Deuteropoda before the LCA of Euarthropoda.
Whom do we believe? Lyrarapax is known from one specimen, Stanleycaris from 268, preserved in various orientations.
For now, we believe Stanleycaris. Without the fossils, we couldn't have known.
Desmond Collins and David M. Rudkin. 1981. Priscansermarinus barnetti, a probably lepadomorph barnacle from the Middle Cambrian Burgess Shale of British Columbia. Journal of Paleontology 55(5), 1106-1015
Peiyun Cong, Xiaoya Ma, Xianguang Hou, Gregory D. Edgecombe, and Nicholas J. Strausfeld. 2014. Brain structure resolves the segmental affinity of anomalocaridid appendages. Nature 513, 538-542.
Gregory D. Edgecombe, Christine Strullu-Derrien, Tomasz G—ral, Alexander J. Hetherington, Christine Thompson, and Markus Koch. 2020. Aquatic stem group myriapods close a gap between molecular divergence dates and the terrestrial fossil record. Proceedings of the National Academy of Sciences 117 (16) 8966-8972.
Romain Garrouste, Gael Clement, Patricia Nel, Michael S. Engel, Philippe Grandcolas, Cyrille D'Haese, Linda Lagebro, ulien Denayer, Pierre Gueriau, Patrick Lafaite, Sebastien Olive, Cyrille Prestianni, and Andre Nel. 2012. A complete insect from the Late Devonian period. Nature 488, 82-85.
Ronald A. Jenner , Cees H.J. Hof , Frederick R. Schram. 1998. Palaeo- and archaeostomatopods (Hoplocarida: Crustacea) from the Bear Gulch Limestone, Mississippian (Namurian), of central Montana. Contributions to Zoology 67(3): 155-186.
David A. Legg, Mark D. Sutton & Gregory D. Edgecombe. 2013. Arthropod fossil data increase congruence of morphological and molecular phylogenies. Nature Communications 4(2485).
Jianni Liu, Rudy Lerosey-Aubril, Michael Steiner, Jason A Dunlop, Degan Shu, John R Paterson. 2018. Origin of raptorial feeding in juvenile euarthropods revealed by a Cambrian radiodontan. National Science Review, 5(6), 863Đ869.
Xiaoya Ma, Xianguang Hou, Gregory D. Edgecombe, and Nicholas J. Strausfeld. 2012. Complex brain and optic lobes in an early Cambrian arthropod. Nature 490, 258-261.
Christine Martin and Georg Mayer. 2015. Insights into the segmental identity of post-oral commissures and pharyngeal nerves in Onychophora based on retrograde fills. BMC Neuroscience 16, 53 (2015)
Marco T. Neiber, Tamara R. Hartke, Torben Stemme, Alexandra Bergmann, Jes Rust, Thomas M. Iliffe, Stefan Koenemann. 2013. Global Biodiversity and Phylogenetic Evaluation of Remipedia (Crustacea). PLOS|One May, 2011.
Jakub Prokop and Andre Nel. 2011. New Middle Permian palaeopteran insects from LodŹve Basin in southern France (Ephemeroptera, Diaphanopterodea, Megasecoptera). Zookeys (130): 41Đ55..
Jerome C. Regier, Jeffrey W. Shultz, Andreas Zwick, April Hussey, Bernard Ball, Regina Wetzer, Joel W. Martin, and Clifford W. Cunningham. 2010. Arthropod relationships revealed by phylogenomic analysis of nuclear protein-coding sequences. Nature 463, 1079-1083.
Gengo Tanaka, Xianguang Hou, Xiaoya Ma, Gregory D. Edgecombe, and Nicholas J. Strausfeld. 2013. Chelicerate neural ground pattern in a Cambrian great appendage arthropod. Nature 502, 364-367.
Dieter Waloszek. 1993. The Upper Cambrian Rehbachiella, its larval development, morphology and significance for the phylogeny of Branchiopoda and Crustacea. Developments in Hydrobiology 103: 1-13.
Heather M. Wilson and Lyall I. Anderson. 2004. Morphology and taxonomy of Paleozoic millipeses (Diplopoda: Chilognatha: Archipolypoda) from Scotland. Journal of Paleontology 78(1): 169-184.


























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
,_Mook,_the_Netherlands.JPG){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}