Fall Semester 2022 Overview of Animal Phylogeny & Cnidarians
Great Barrier Reef - Queensland.
Key Points:
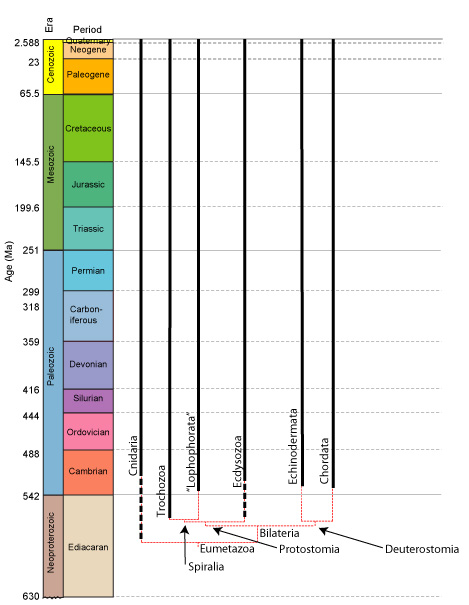
•Eumetazoa - metazoans closer to Bilateria than to "Porifera" includes creatures with proper endoderm and ectoderm, displaying gastruation.
•Basal eumatazoans, including Placozoa and Ctenophora have negligible fossil records.
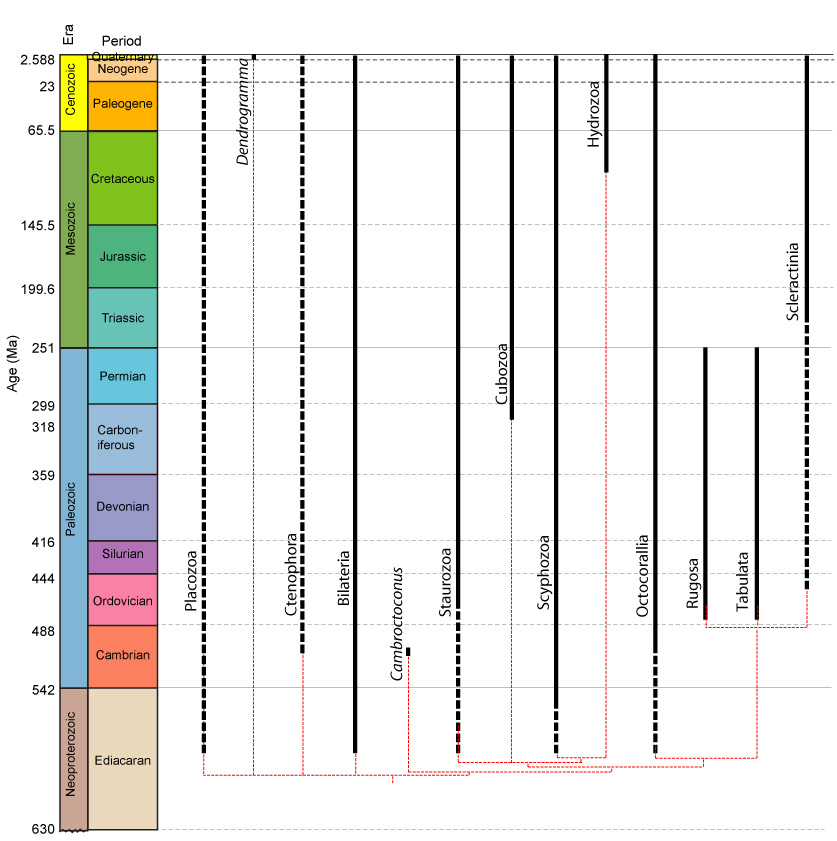
•Cnidaria are characterized by distinct body plans, cell types, and reproductive strategies.
•Cnidarians ancestrally alternate between polyp and medusa generations, however different branches may modify this pattern, reducing or eliminating one stage or the other.
•Only the cnidarian groups whose members secrete calcareous skeletons (Hydrozoa and Anthozoa) have abundant fossil records.
•Rugosa and Tabulata are limited to the Paleozoic. They were significant elements of Silurian and Devonian reef environments, but less common in the Carboniferous and Permian.
•Scleractinia have been present since the Triassic but only came to dominate reef environments during the Cenozoic.
•The earliest definite cnidarian fossils are from the Cambrian, however Ediacaran organisms such as Namapoikea and Cloudina might be stem cnidarian.
•Conulariids are enigmatic Paleozoic organisms that have been allied with Staurozoa and Scyphozoa, but also considered not to be proper metazoans.
"The coral that grows at the edge of the reef is always the strongest and most colourful because it faces the greatest battering. It's the same if you're called Honeysuckle. I'd have had a totally different life if I'd been called Mary."
(Honeysuckle Weeks.)
Eumetazoa:
(Ediacaran - Quaternary) Includes Cnidaria and Bilateria. (Many regard Ctenophora as members.)
Major taxa:
Cnidaria: (Ediacaran - Quaternary) corals, jellyfish, etc.
Bilatera: (Ediacaran - Quaternary) Animals with bilateral symmetry, and distinct mouth and anus.
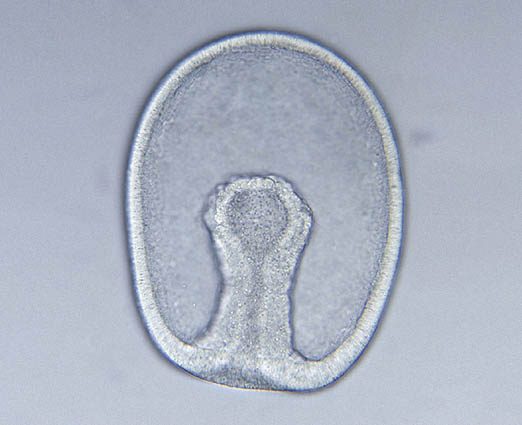
result from the process of gastrulation (right), in which cells of the unilayered blastula stage embryo invaginate to form a two-layered gastrula with a gut cavity that communicates with the exterior through a blastopore. But there is more than this to creating an organized body.
Hox genes.
We now turn to the eumetazoans with substantial fossil records.
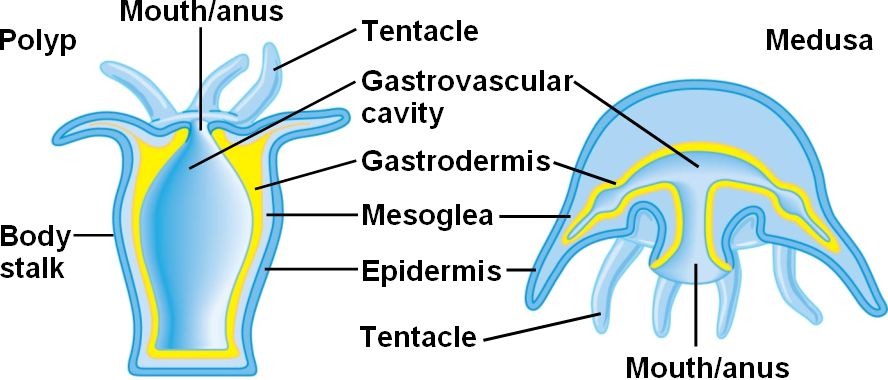
Diploblastic - having endoderm and ectoderm only, separated by gelatinous extracellular mesoglea.
Radially symmetrical body plan (right) with single opening to the gut, termed the "mouth" for the sake of politeness. Cnidarians have no front, back, left, or right, but we can refer to their oral and aboral aspects.
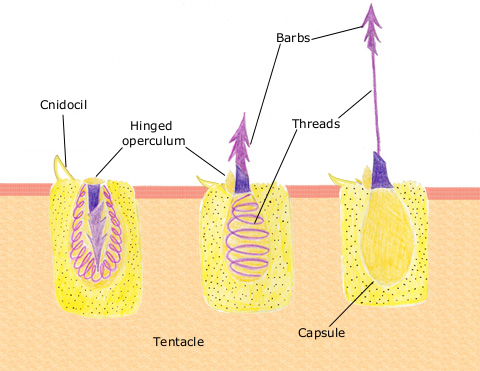
Prey capture by Cnidocytes (stinging cells) born on tentacles surrounding mouth.
One typically hears that cnidarians have tissues but not organs. Typically true, except that gametes are produced in proper gonads. A few other scattered exceptions noted below.
Major cell types: Cell types among cnidarians are no more diverse than in sponges, but are clearly organized into distinct tissues. These include:
Epitheliomuscular cells forming organized muscle fibers in both ectoderm and endoderm.
Nerve cells, including sensory and motor neurons, organized into a decentralized nerve net.
Gland cells of the endoderm secrete digestive enzymes.
Interstitial cells - undifferentiated totipotent cells that can transform, as needed, into other cell types.
Cnidocytes (right) containing nematocysts capsules - the stinging apparatus. The nematocyst stores Ca2+ ions. When triggered by a mechanical disturbance, these are released into the cytoplasm, creating an osmotic gradient that causes water to flow into the cell, inflating the capsule and extending the barb.
Reproduction: Cnidarians typically alternate between asexually reproducing polyp and sexually reproducing medusa generations (although in various groups, one or the other of these generations may be brief or absent.)
In sexual reproduction, the zygote develops into a planktonic gastrulated planula larva that settles down to develop into a polyp.
Asexual reproduction can take the form of:
Fission: where single polyps divide in two then regenerate lost parts.
Budding: New individuals bud off of parents. Polyps routinely produce medusae in this way. In some (E.G. hydrozoans) budding is off of the side, in others (Scyphozoans) it occurs along the body axis at the oral end (right).
Although generations alternate in most studied taxa, the polyp stage is reduce and medusa predominates. Medusae bud off of the oral ends of polyps (in contrast to hydrozoans.)
Fossil record: Restricted to Konzervatlagerstätten such as Burgess Shale (Cambrian), Holtzmaden (Jurassic), and Solnhofen (Jurassic).
Claims of Ediacara hills (Ediacaran) scyphozoans are controversial, as the fossils (E.G. Mawsonites) are impressions that might represent the holdfasts of other organisms.
Body plan shows four-fold radial symmetry with four tentacles or groups thereof.
Medusa profile is square when viewed from above.
Four eyes, complete with retinas and corneae, (organs!) facilitate more active pursuit of prey and obstacle avoidance.
Medusae appear to practice internal fertilization. Polyp generation reproduces asexually, but eventually, rather than budding medusae, polyps undergo a metamorphosis to become sexually reproducing medusae.
Fossil record:
Restricted to two Konservat-Lagerstätten, Mazon Creek (Carb) and Solnhofen (J).
Hydrozoa: Hydras, fire coral (Cretaceous - Quaternary)
Characteristics:
Marine and fresh-water. The freshwater Hydra is typical.
Generations alternate, but medusa stage is reduced almost to the point of functioning as gonads for the polyp. Polyps may reproduce asexually by budding from the side of the polyp (in contrast to the scyphozoan pattern.)
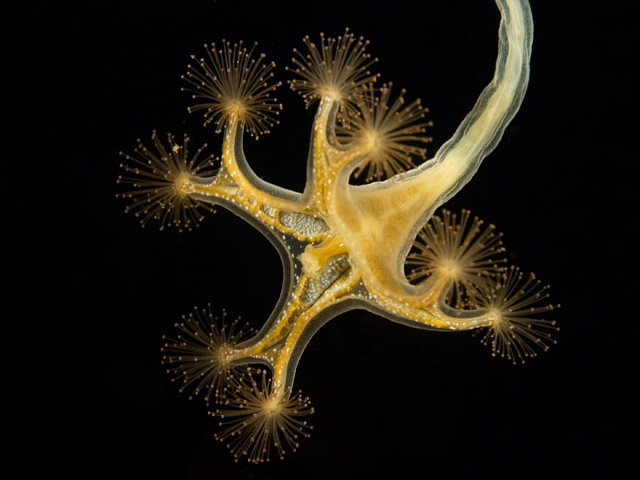
Solitary or colonial. Marine colonial forms include blade fire coral (right), which secretes calcareous skeleton.
Major groups: Here we must simply punt. Hydrozoan systematics are in a state of profound disagreement and revision. The following are interesting hydrozoan ecomorphs whose monophyly is entirely uncertain:
Milleporina (right) and Stylasterina (hydrocorals): Marine colonial forms include fire coral, which secrete calcareous skeleton.
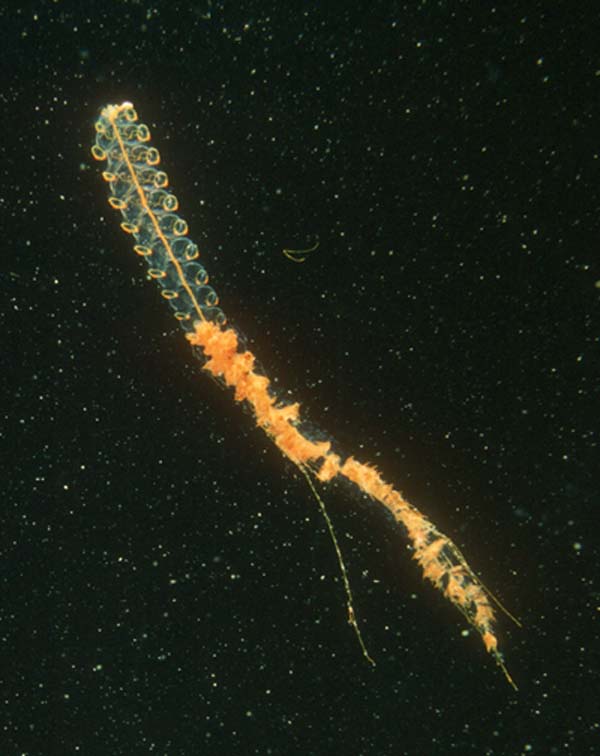
Siphonophora: Pelagic (free-swimming) colonial marine forms, such as Portuguese Man-o-War, showing radical specialization of zooids (both polyp and medusa in one colony) into:
Pneumatophores: Bell or sail for floatation.
Nectophores: For swimming.
Gastrozoids: For feeding.
Etc.
Fossil record:
Milleporids and stylasteroids known from Cretaceous onward.
Body plan shows four-fold radial symmetry with four tentacles or groups thereof.
Lack polyp generation.
Medusae are stalked.
Fossil record: Either:
Haootia quadriformis (Liu et al. 2014) From the late Ediacaran, this is interpreted as a cnidarian-grade eumetazoan that is suspiciously similar to a staurozoan. That would make it the oldest staurozoan and the oldest cnidarian.
But see conulariids (Cam. (or is that E?) - Tr) below.
Thus, the octocorallian record securely begins in the Middle Cambrian.
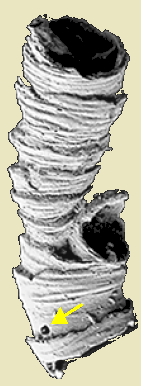
Rugose corallite from Hooper Museum. (PSST! Something is wrong!)
Zoantharia:(Cambrian - Recent) This group contains the vast majority of calcareous skeleton secreting cnidarians. Major issues:
Each major group seems, independently to have descended from a soft-bodied ancestor that achieved the ability to secrete a calcareous skeleton.
Hermatypic members of each group hosted photosynthesizing zooxanthellae, enabling them to grow and secrete CaCO3 much more rapidly. These tend to be the major reef-formers. They are restricted to the photic zone.
Ahermatypic members lack zooxanthellae and tend to be solitary and less abundant, but can colonize below the photic zone.
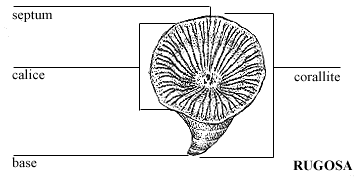
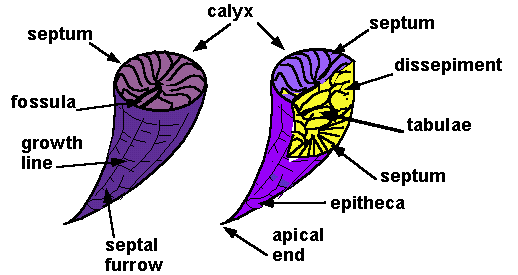
Corallites of the major zoantharian groups display the same characteristic features but in different combinations (see right):
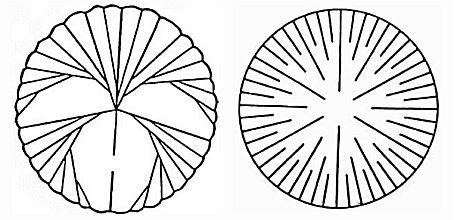
Septa: Vertical partitions projecting from the sides that support pleated infoldings of the polyp, that increase the surface area of its gut. Different taxa have distinct patterns of septum growth visible in the calyx (aperture of the corallite). Plural = calices. Note: "calice" is not a word!
Tabulae: As the polyp grows it moves upward in the conical corallite, then secretes a new floor to support it. The result is a series of horizontal sheets perpendicular to the long axis of the corallite.
Dissepiments: Small upwardly convex "bubbles" of CaCO3 that fine-tune the shape of the corallite.
Largely ahermatypic, not major reef framework builders, but crucial pioneers of soft sediments, as their skeletons provided hard substrates for later framework builders.
Suffered greatly in Devonian extinction. Persisted at lower abundance and diversity to the Permian extinction.
Paraphyletic with respect to Scleractinia? Possible, although equally likely that scleractinian skeleton is convergent.
Attach to substrate by a basal plate, and can secrete aragonite onto outer surface of corallite, facilitating solid attachment and growth of large colonies (in contrast to rugosans.)
In living members, the ability to form large reefs depends on zooxanthellae. Thus, major reefs limited to photic zone of tropics. Major reef building corals (with zooxanthellae), thus hermatypic.
In many scleractinians, the polyp and calyx, in oral view, becomes greatly elongate, with colony members forming convoluted interlocking patterns. E.g. "brain-coral" (right).
Minor reef formers in Late Triassic and Jurassic, however their fortunes waned in the Cretaceous, when the major reef-building was done by rudistid bivalves.
But bounced back after the K-P extinction as the Primary reef-builders of Cenozoic.
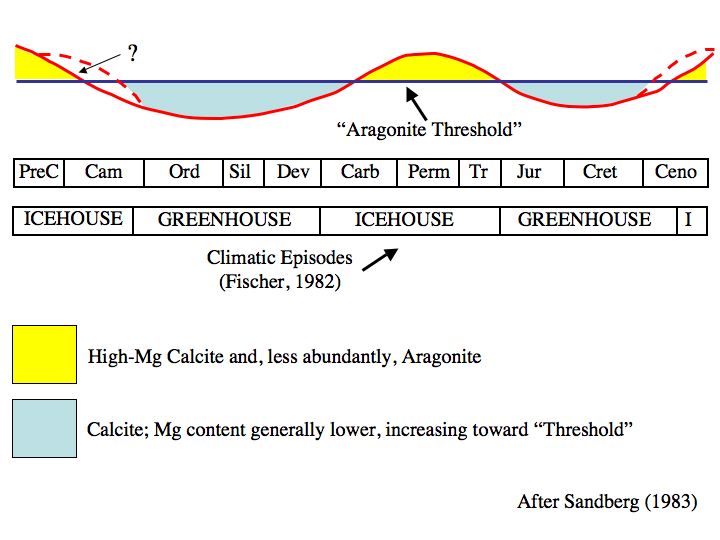
Indeed, the abundance of the aragonite-secreting scleractinians roughly tracks the tendency of CaCO3 to precipitate directly as aragonite or calcite. This deposition is sensitive to the ratio of Mg/Ca, as aragonite can accommodate more Mg through cation substitution. Normal ocean environments are very close to the boundary, such that minor changes in Mg concentration result in global shifts in carbonate deposition. Thus, Earth history has seen alternations between periods of aragonite seas and calcite seas. This, in turn reflects rates of sea floor spreading:
Indeed, the abundance of the aragonite-secreting scleractinians roughly tracks the
Calcite seas: During intervals of rapid sea floor spreading, hydrothermal activity near spreading zones pumps Ca into the oceans, but tend to withdraw Mg through hydrothermal reactions with ocean bedrock. Thus, oceans are Mg-poor and primary deposition is of low-Mg calcite. These also tend to be intervals of higher sea level.
Aragonite seas: During intervals of slow sea floor spreading, oceanic concentrations of Ca are lower and Mg are higher, favoring aragonite and high-Mg calcite deposition. These tend to be intervals of lower sea levels.
Ries et al., 2006 demonstrated that scleractinians raised in water with the Ca/Mg balance of the Cretaceous spontaneously secrete calcite, but grow much more slowly.
Scleractinian origins: All skeleton-secreting zoantharians probably derived from soft-bodied forebears like the Early Cambrian Archisaccophyllia of Chengjiang (Han et al., 2010). Thus, their early history is encrypted. In the case of scleratinians, we know of rare Paleozoic anthozoans with the scleractinian septal pattern, including Kilbuchophyllum (Ord). Unlikely that later calcified scleractinians are derived from it. More likely, Kilbuchophyllum testifies to the existence of the scleractinian septa pattern in soft anthozoans of the time, but represents an independent derivation of calcification.
Cloudina
Cnidarian origins:
A cloudy picture. The last common ancestor of Porifera and Cnidaria is difficult to visualize. Calibrated molecular clocks suggest:
Cryogenian (~660 ma) LCA for Metazoa
Early Ediacaran (~630 ma) LCA for Eumetazoa
Latest Ediacaran/ Earliest Cambrian (~543 ma) LCA for Cnidaria
Some tantalizing hints:
Namapoikea, a meter-scale modular colonial organisms of the Ediacaran Nama Formation of Namibia.Whether this was closer to sponges or cnidarians is completely unclear.
Cloudina (right), calcareous tubes of internested narrow cones found worldwide in the Ediacaran. The presence of some with budding structures leads to speculation that they were cnidarians. (Although some consider them likely to be bilaterians.)
Inconclusive but not out of line with molecular clocks. An aside: (Penny et al., 2014) report that Cloudina in addition to being the first widespread mineralizer, was also the first reef-builder, in association with bacterial stromatolites. They are currently a research focus of UMD's own Dr. Jay Kaufman!
Even the ancestral condition of cnidarians is problematic.
Some questions that arise:
Was the ancestral cnidarian colonial or solitary?
Was it nektonic or sessile?
Did it experience alternation of generations?
If not, was it a polyp or medusa?
Did polyps bud asexually in the manner of hydrozoans or scyphozoans?
Did it have fourfold symmetry (as in cubozoans, scyphozoans, and staurozoans) eightfold (as in Octocorallia) or none?
Available information is ambiguous:
The oldest fossils seem to be of solitary forms, but the most basal branch, Anthozoa, is primarily colonial.
If we assume that Ediacaran scyphozoan and conulariid fossils are legitimate, then both free and attached forms were present throughout the Phanerozoic.
The oldest fossils seem to be medusae, but the most basal branch, Anthozoa, has only polyps.
But for conulariids, with fourfold symmetry, there is no clear pattern of symmetry.
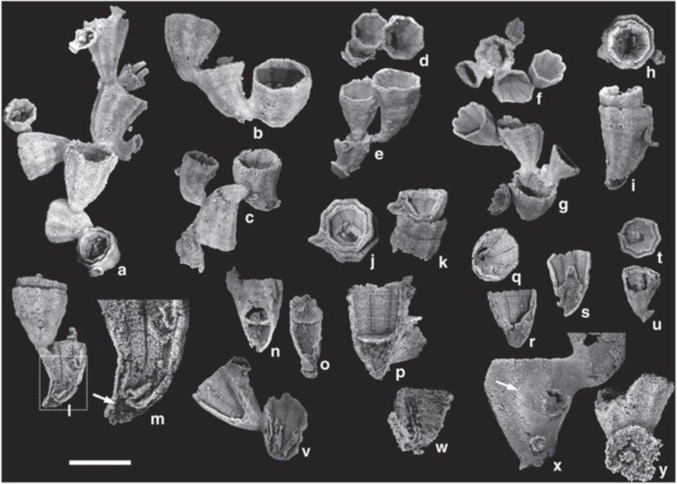
Some illumination has been provided by Park et al., 2011, with the description of the Middle Cambrian Cambroctoconus orientalis (right scale is 1 cm.) Argued to be a "stem cnidarian" (i.e. the sister taxon to proper Cnidaria) it appears to be:
Colonial with a hard skeleton (the earliest known from the cnidarian stem or crown.)
Consisting of polyps in which individual polyps bud asexually off of their parents from the side, like modern Hydra (or Cloudina), not like scyphozoans.
Possess septa but no dissepiments and at most one tabula.
Van Iten et al., 2013 describe an unnamed three-sided conulariid-like animal from the Ediacaran of China.
Relationships?: Two substantive hypotheses:
Staurozoa , Traditionally seen as closer to Staurozoa based on four-fold symmetry and general similarity to staurozoan medusa.
Scyphozoa, Van Iten et al. 2006 regard conulariids as close to the Coronatae within Scyphozoa.
Vendobiont based on the arguable presence in the Ediacaran. That these early conulariids (if that's what they are) have three-fold symmetry undermines the argument for a relationship with Cnidarians. Ivantsov and Fedonkin, 2010 ask if they be independently derived from other three-fold symmetric Ediacarans like Tribrachidium
Stay tuned.
Bilateria:
Sometimes called "Bilateralia" in older literature. Monophyletic group containing most animals. In simple terms, bilaterians are animals with a distinct front, back, top, and bottom.
Fundamental synapomorphies:
An antero-posterior axis (rather than an oral-aboral axis)
Bilateral symmetry (although this may be suppressed or overprinted in some)
Presence of a flow-through gut with distinct mouth and anus.
Triboblastic: I.e. a third germ layer, mesoderm is present in addition to the endoderm and ectoderm of cnidarians. Mesoderm replaces the passive extracellular mesoglea of cnidarians with three-dimensional living tissue.
Body segmentation.
Bilaterian Development:
Cleavage from A. S. Romer. 1977. The Vertebrate Body.
Consider the basic steps by which we get a proper embryo with a top, bottom, front, and back from a zygote:
Polarity: In all ova yolky matrial tends to concentrate at one end, yeilding:
"Animal" pole, the non-yolky side
"Vegetal" pole, the yolky part.
The amount of yolk greatly influences developmental dynamics. For now, we consider an ovum with relatively little yolk.
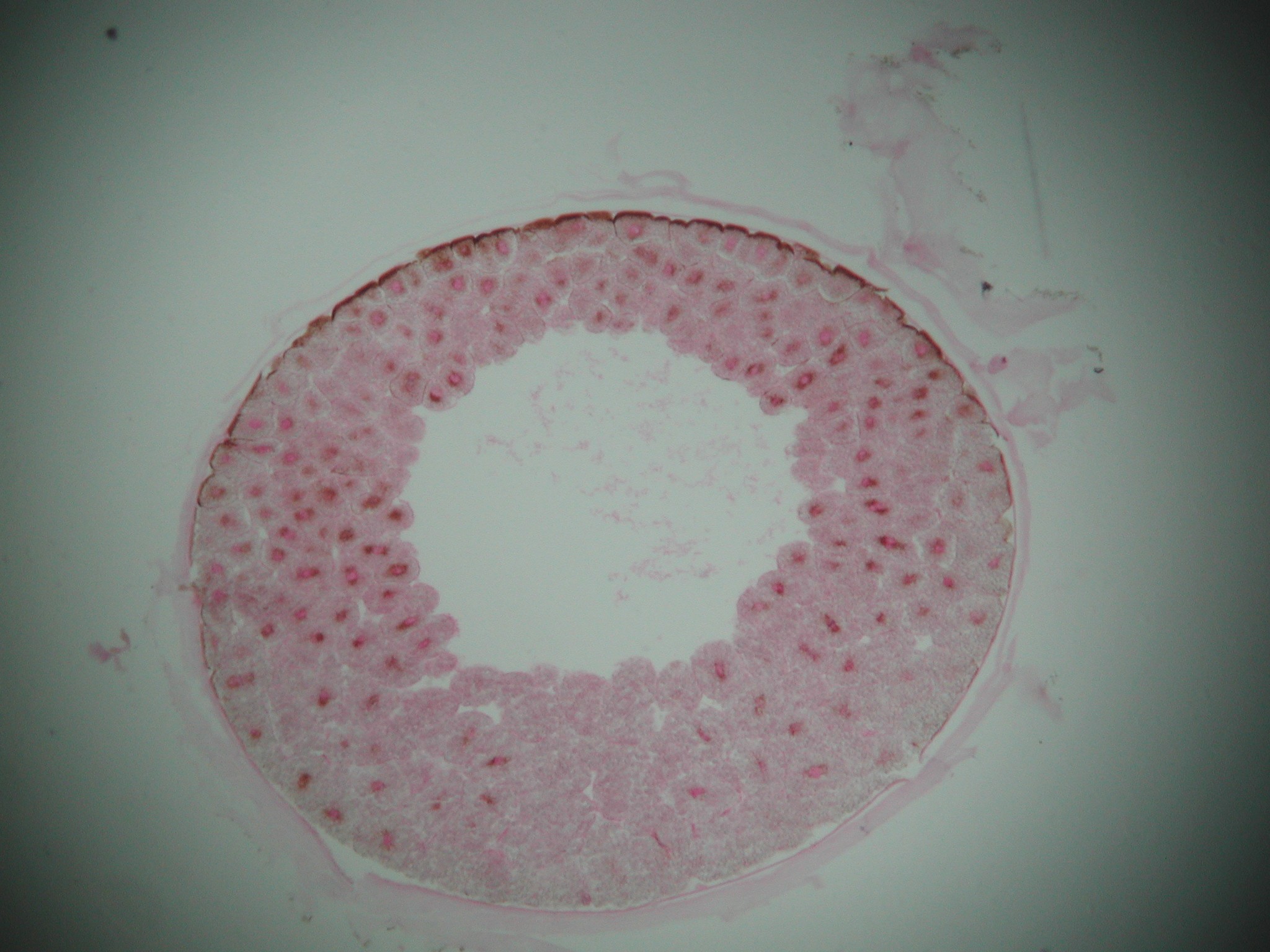
Cleavage Phase of rapid cell division with little overall growth. The zygote transforms into a hollow sphere of cells, the blastula. The space in the middle is the blastocoel
The blastula has three cell types:
Smaller cells nearer the "animal" pole.
Larger, yolky cells nearer the "vegetal" pole.
A third type of cell girdling the large cells of the "vegetal" pole.
Gastrulation from A. S. Romer. 1977. The Vertebrate Body.
Gastrulation:
Cells of the "vegetal" hemisphere form a flat plate then invaginate inward, yielding a two layered hemisphere.
The rim of this concavity qickly closes into a small opening, the blastopore. While this is occurring, cells from the third cell type stream in through the blastopore and move forward along the inner surface. These become mesoderm. Depending on the taxon, blastopore will become either mouth or the anus.
The embryo is now a gastrula. It possesses three basic germ layers:
Endoderm: Destined to give rise to the gut tube and associated structures.
Ectoderm: Destined to give rise to the skin and, in vertebrates, central nervous system.
Mesoderm: Destined to give rise to a large range of internal organs (depending on the taxon.)
Note: This should look slightly familiar, as it is basically a cnidarian planula larva with mesoderm added.
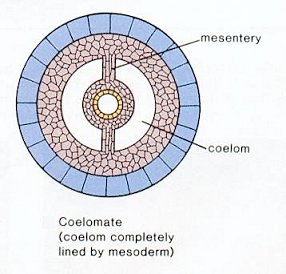
The Coelom: A characteristic feature of bilaterians is the presence of a coelom or body cavity. This feature allows for:
increased gut expansion,
hydrostatic structure against which muscles may act, facilitating purposeful locomotion
use of coelomic fluid in gas and nutrient transport, breaking dependence on simple diffusion and allowing larger body size.
Developed as:
Eucoelomate: In which a proper coelom develops within the mesoderm. (right)
Pseudocoelomate: In which the coelom develops between the endoderm and mesoderm as a remnant of the blastocoel and is properly called the pseudocoel. Rotifers, kinorhnchs, nematodes, and a few other minor taxa.
Acoelomate: In some critters, including platyhelminth flatworms, the coelom is drastically reduced or absent.
The evolution of the coelom opened many vistas for animal evolution, including significantly expanded locomotor strategies. Cnidarians show the limits of what a hydrostatic skeleton can do for an animal with a single module. Bilaterians, however, display body segmentation in which separate modules of the hydrostatic skeleton can lengthen and shorten, facilitating much more complex movement. This makes possible activities like:
Purposeful locomotion
Burrowing into the substrate
Indeed, non-bilaterians are deemed incapable of burrowing.
Specialized organs: These capabilities came at a price. Animals with only endoderm and ectoderm don't need to worry about gas exchange and elimination of nitrogenous waste, because no living cell is so far from the body surface that simple diffusion can't do the trick. Bilaterians, in contrast, usually require specialized organs for functions like:
Gas exchange
Nitrogenous waste elimination
Circulation
Coordination of the nervous system
Sensing the environment (useful to motivate and manage locomotion)
Fortunately, the presence of mesoderm and a coelom seems to bestow the developmental plasticity needed to allow these to evolve. Indeed, the gut tube, kidneys, and gonads are ancestrally suspended inside the coelom.
Hox Genes:
First, fully appreciating the significance of segmentation as a synapomorphy of Bilateria, and the deep relationship shared by bilaterians requires an excursion into the realm of genomics:
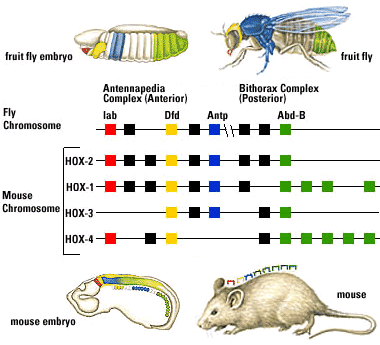
Hox genes: During the late 20th century it became known that segmentation in bilaterian bodies is governed by a special class of regulatory genes. The story:
Fruit flies are a favorite model for geneticists, with short generation spans and interesting mutations that often effect entire sections of the body (modifying or eliminating body segments and/or the appendages that grow from them). Investigation into these segmentation-altering mutations revealed that they can be caused by mutations to eight genes. What makes this interesting:
The genes are close to one another on a single chromosome, and are physically lined up in the same order as the segments of the body they effect.
Each gene (of ~ 1000 base pairs) contains a nearly identical sequence of about 60 base-pairs called homeoboxes.
The homeobox codes for a protein called the homeodomain that is functionally similar to regulatory proteins that block or allow transcription of other genes. It appears that the cluster of eight genes controls the identity of body segments in fruit flies. These are called Hox genes, after the homeoboxes they contain. The homeodomain protein regulates transcription of the Hox gene, whose protein products, themselves, promote or inhibit the synthesis of other regulatory proteins that govern body segmentation. Thus, Hox genes and the Hox gene clusters are special classes of operon. But that is the tame part!
But it gets better: The search for homologs to fruit fly Hox genes found them in almost every animal surveyed. Mammals, for example, have four clusters of Hox gene homologs, in each of which the genes occur in the same order on the chromosome as the regions for which they code.
This is huge.
The fruit flies and mammals belong to the two major clades of bilaterians, and their last common ancestor lived during (or before) the Ediacaran, and yet they share important regions of the genome.
The fruit flies and mammals belong to the two major clades of bilaterians, and their last common ancestor lived during (or before) the Ediacaran, and yet they share important regions of the genome. Moreover, two Hox genes have been identified in the cnidarian Nematostella and are involved with the patterning of its oral/aboral axis. We will meet Hox genes again, but know that they are present from the beginnings of Eumetazoa.
















{kind=link}
/planulaPR-5829d4013df78c6f6ae44a9d.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}