


Key Points:
•"Functional morphology" studies the connection between organismal form, behavior, and dynamics.
•Upon scrutiny, the term encompasses at least three distinct approaches: Biomechanics, theoretical morphology, and paleontological applications.
•Morphospace is the theoretical universe of possible organismal morphologies. Depending on one's approach, it can be defined in terms of measured anatomical features or through multivariate statistical analysis.
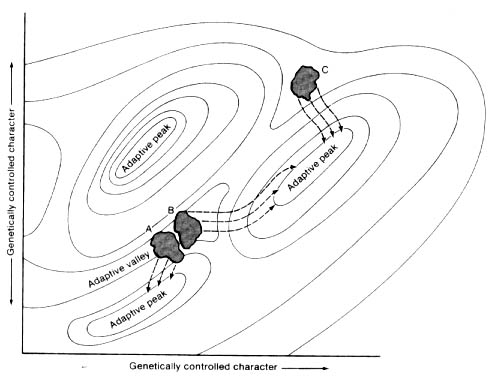
•Morphospace resembles the "adaptive landscape" metaphor of Sewell Wright.
•Ecological morphospace represents the morphospace occupied by inhabitants of a given environment.
•So much in functional morphology depends on the assumption that natural selection is the sole determinant of morphology, however this "adaptationist program" is regarded as naive by some. Structural constraints, evolutionary heritage, pleiotropy, competing selective pressures, and generic variation can all impart their own non-adaptive signal.
•Testable functional hypothesis can be difficult to develop.
•The comparative method - the interpretation of one organism's features by analogy with those of a better known organism - is a tradition in biology, but can be difficult to implement.
•Some hypotheses are clearly testable, including whether a particular function would require a biological material to exceed its known strength or a muscle to to exceed its known energy output.
"Whether it be the sweeping eagle in his flight, or the open apple-blossom, the toiling workhorse, the blithe swan, the branching oak, the winding stream at its base, the drifting clouds,
over all the coursing sun, form ever follows function, and this is the law. Where function does
not change form does not change."
(Louis H. Sullivan, 1896.)
Origins and biases: But why would we even expect there to be a connection between form and function? Our faith in this idea springs from three sources:
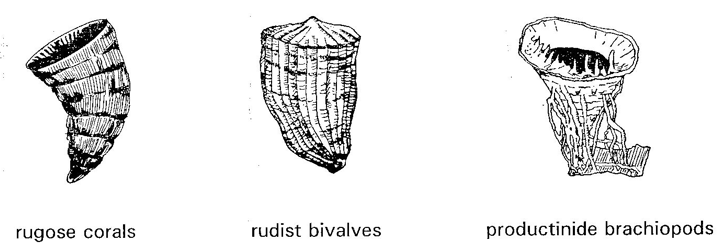
This only seems explainable in terms of the primacy of natural selection.
However, When examined in detail, people calling themselves functional morphologists don't all do the same things. In fact, three broad categories of related but distinct research tend to get conflated under this term:
Obviously, this only works directly on living organisms, however observations of them can indirectly illuminate interpretations of ancient ones.
As paleontologists, we are primarily concerned with the last two.
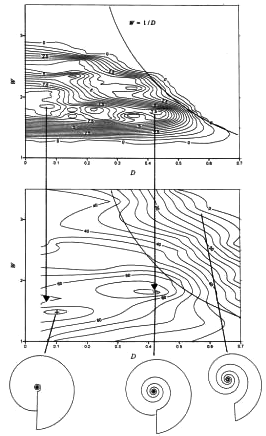
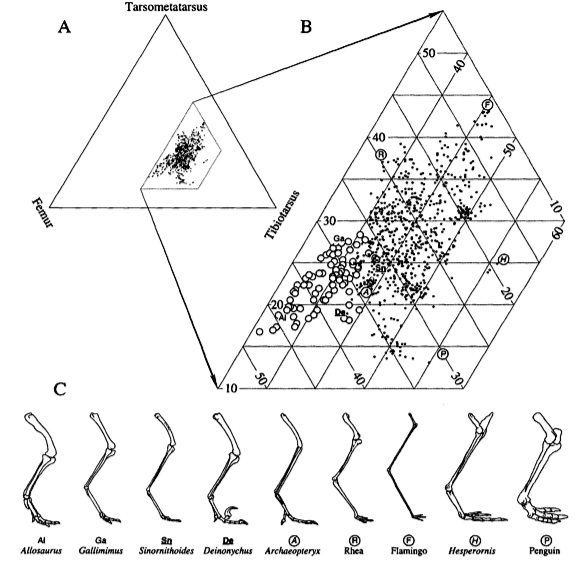
Theoretical vs Functional Morphology: The description, in abstract terms, of morphospace - the universe of possible organismal morphologies. Morphospace is, by necessity, an abstraction. Typically, the range of morphological variation in comparable organisms is reduced to two or three principle axes. These may be:
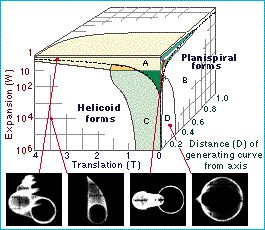
Caveat: The degree to which these favored measurements truly encompass morphological diversity is not knowable. Raup attempted to reduce coiled mollusk shell morphology to three parameters:
(A Tom Lehrer song inspired by Wright's exploits.)
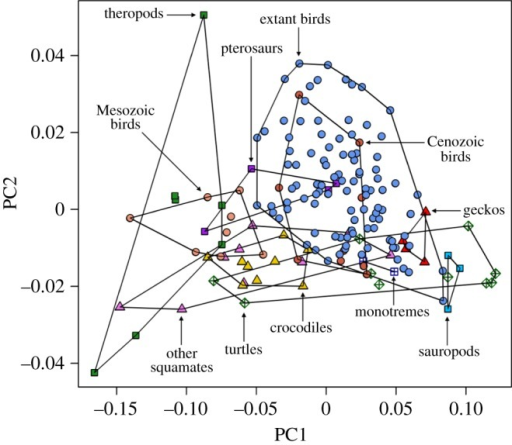
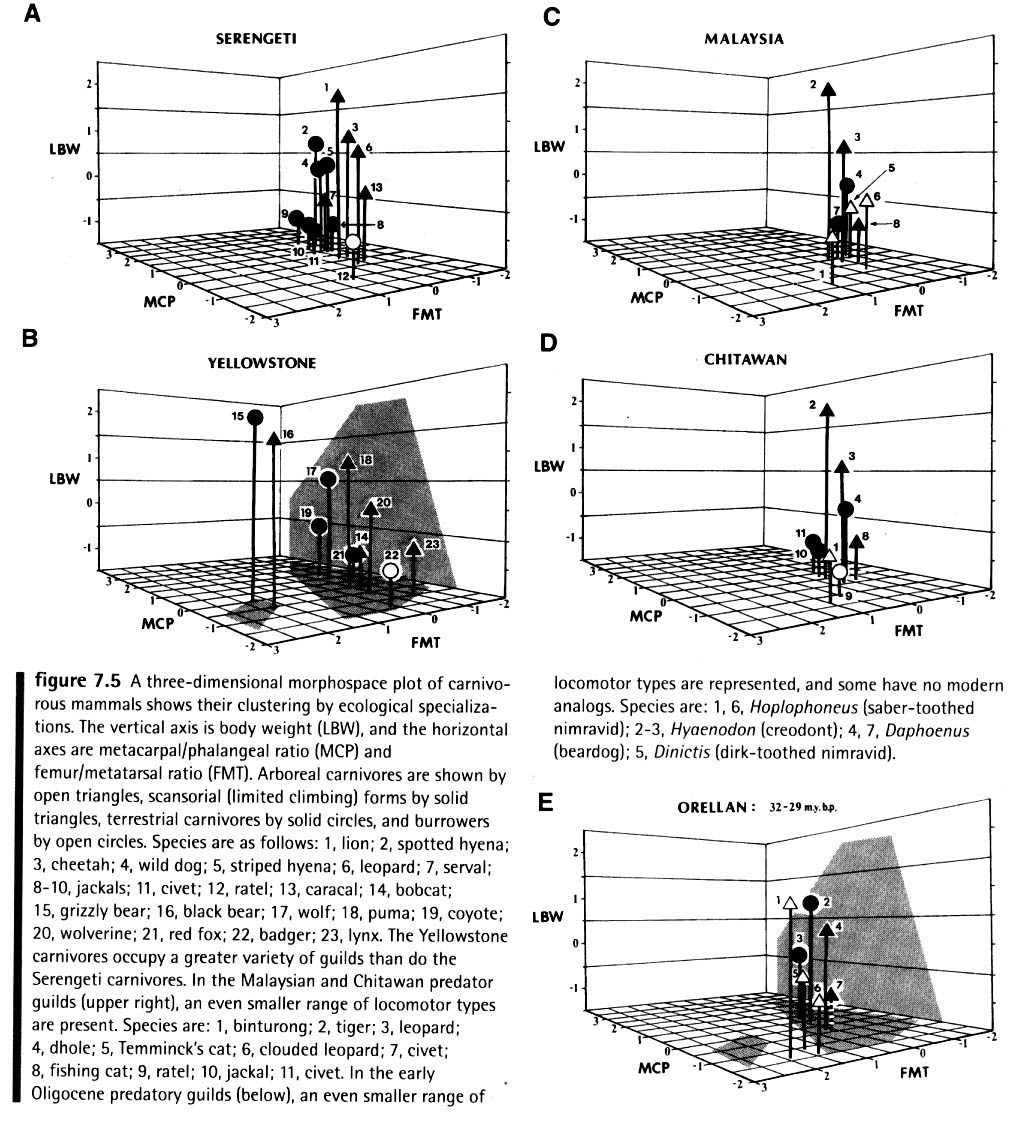
Ecological morphospace: Modern and ancient environments can be characterized in terms of the morphospace occupied by their occupants, as in Van Valkenburgh, 1988.

The adaptive landscape, and much of functional morphology, rests on the notion that adaptation under the influence of natural selection is the paramount determinant of morphology. For much of the 20th century, morphologists counted on this. Could they reasonably do so?
Gould and Lewontine (1979) took aim at the notion, and the reflexive tendency of "pan-selectionist" researchers to offer adaptive secenarios for every aspect of morphology. They compared this to attempts to infer structural function to spandrels, sections of wall filling spaces between load-bearing components like arches and domes in classic Medieval churches. (See the spandrels of San Marco right) In fact, spandrels are just space-fillers between functional elements.
A biological comparison: the human chin - a functionless feature that is a developmental consequence of the reduction of the anterior tooth row.
Non-adative influences on morphology include:
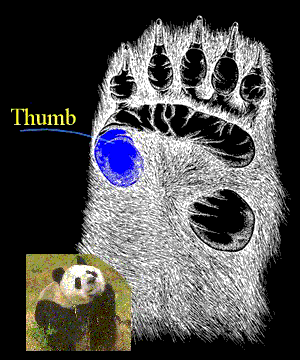
The result is that few structures are adaptively optimal, as in the classic panda's thumb. (Another structure constrained by evolutionary history.)
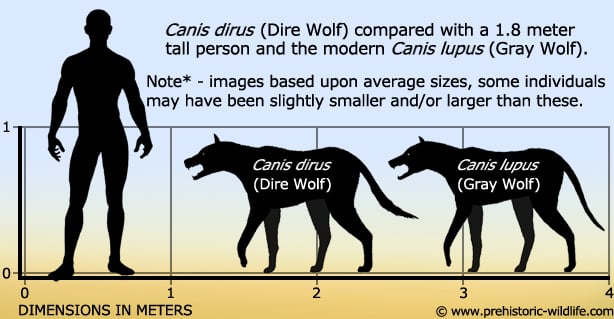
Structures that appear functionally analogous are our starting point for the development of hypotheses about life-style. E.G. numerous anatomical characters demonstrate that the extant timber wolf and the extinct native North American dire wolf (right) are closely related and generally similar. They differ mostly in that the dire wolf's skull and teeth were more robust - a little closer to what one sees in spotted hyaenas or other bone-crushing animals.
Thus, we speculate that the dire wolf was generally similar to the timber wolf, but better adapted to bone crushing. Because we are dealing with an organism and with biomechanical functions that are well understood, this contention can be evaluated with any of the methods listed above.

Alas, so do other, ecologically dissimilar reptiles, including vertical clingers and leapers like Boyd's forest dragon. In this light, speculations on Eudibamus' locomotion seem reckless without taking a wider range of variables into account.
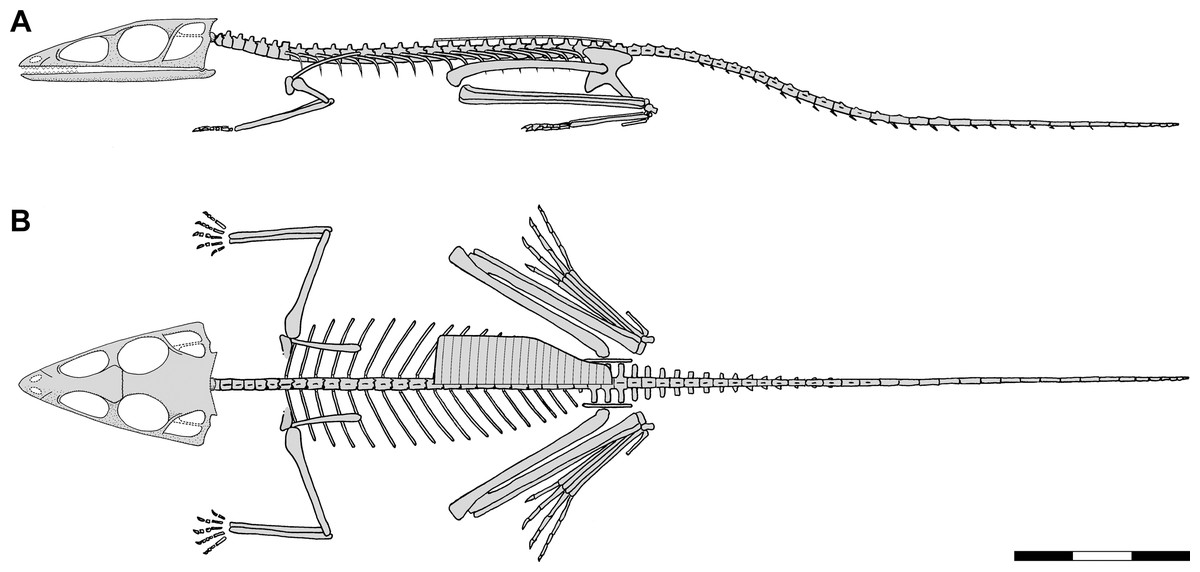
Doing it better: There are several multivariate statistical analysis methods including principal component analysis that can identify the major sources of variation among specimens. These will be discussed in a later lecture. For now, understand that like the examples given above, they provide a basis for comparison of extinct and living taxa. Unlike them, they also provide an algorithmic approach to identifying and weighting the variables of greatest interest. A recent example: Bennett, 2020 who determined that the enigmatic tiny archosaur Scleromochlus was specialized for leaping similarly to frogs.
Consider Parasaurolophus, a Cretaceous lambeosaurine ornithopod dinosaur. Like other ornithopods, it had a deep, laterally compressed torso and tail. The vertebral column of both was stiffened by ossified tendons. The comparison of the deep flat tail to those of swimming vertebrates, combined with the early 20th century conviction that large dinosaurs would have had trouble generating the energy to support their weight on land gave rise to reconstructions of Parasaurolophus as an aquatic creature. The idiosyncratic crest must, therefore, have been a snorkel.
This, at least, was a falsifiable hypothesis. More thoughtful biomechanical analyses of the vertebral column showed that the trunk and tail were inflexible from side to side. Furthermore, the tail vertebrae of actual aquatic reptiles like crocodilians generally had long lateral extensions to give axial muscles better leverage.
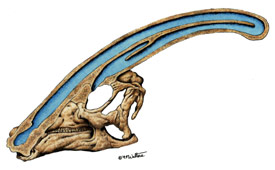
Worse, although the crest connected to the pharynx and nasal cavity though an elaborate system of passages, no specimen of the "snorkel" actually had a hole in the top to admit air. Eventually a revised interpretation of Parasaurolophus as a land animal emerged.
Still, its crest stimulates speculation, including.
Perceiving comparative similarities, like the perception of any pattern in a set of data, is a creative act. Ironically, of all paleontological methods, this one probably has the greatest tendency to push the limits of the proper Scientific Method. In the worst case, the result can be "Just so stories" like those of Rudyard Kipling - appealing speculation with no hope of ever being rigorously tested.
This allows us to make some inferences with confidence. Consider the maximum size of a flying bird: Selective pressure to develop maximum flight thrust is such that in most birds, the major flight muscles already occupy the maximum possible proportion of overall body mass, regardless of size. Thus, they cannot be scaled up allometrically. Huge birds that would require more muscle than this are forbidden. This places a limit on the overall size of an exclusively powered flyer of roughly 12 kg. - roughly the size of the largest powered flyers such as: are the limit. Any larger bird such as the 15 kg. Pleistocene Teratornis merriami must employ some kind of soaring strategy, at least some of the time. Thus, we are confident that a 40 kg. Pteranodon would also have to resort to soaring because of the same constraints.
Indeed, mapping pterosaurs onto theoretical morphospace (see Witton and Habib, 2010, for example) might support this scenario, provided our morphological assumptions are correct and in agreement.
Review: In paleontology, we give different levels of confidence to different hypotheses of function in descending order:
The big thing: Know when you are testing a falsifiable hypothesis and when you are speculating!
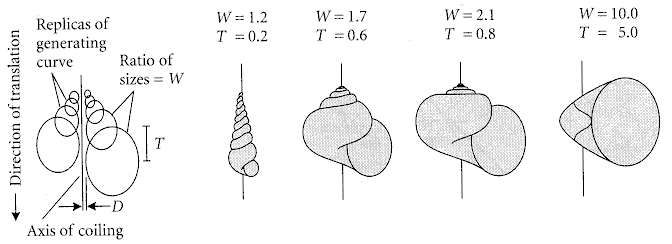








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}