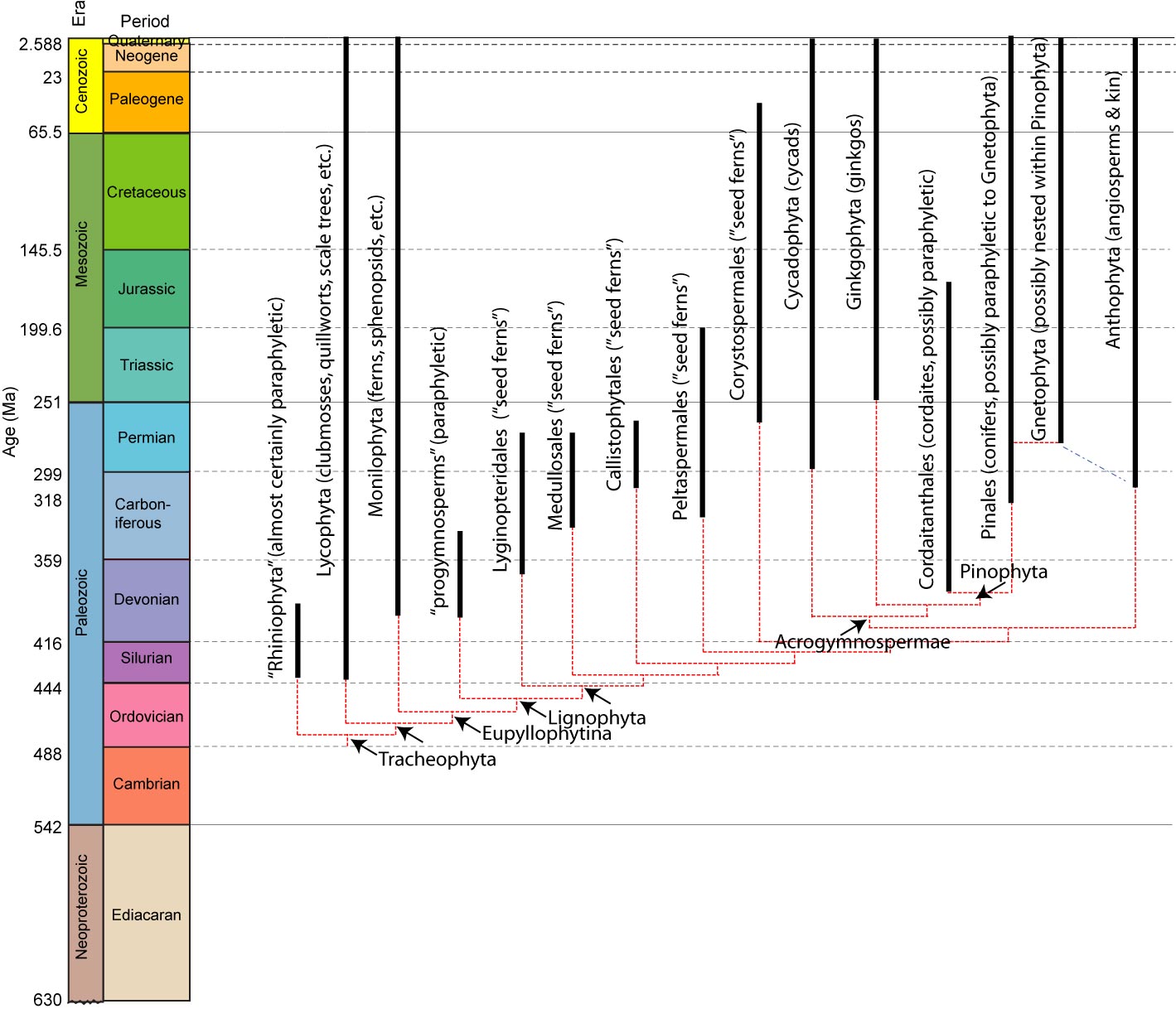
Key Points:
•Fossil plants are vitally important for many reasons.
•Their tendency to fall apart while alive presents special difficulties.
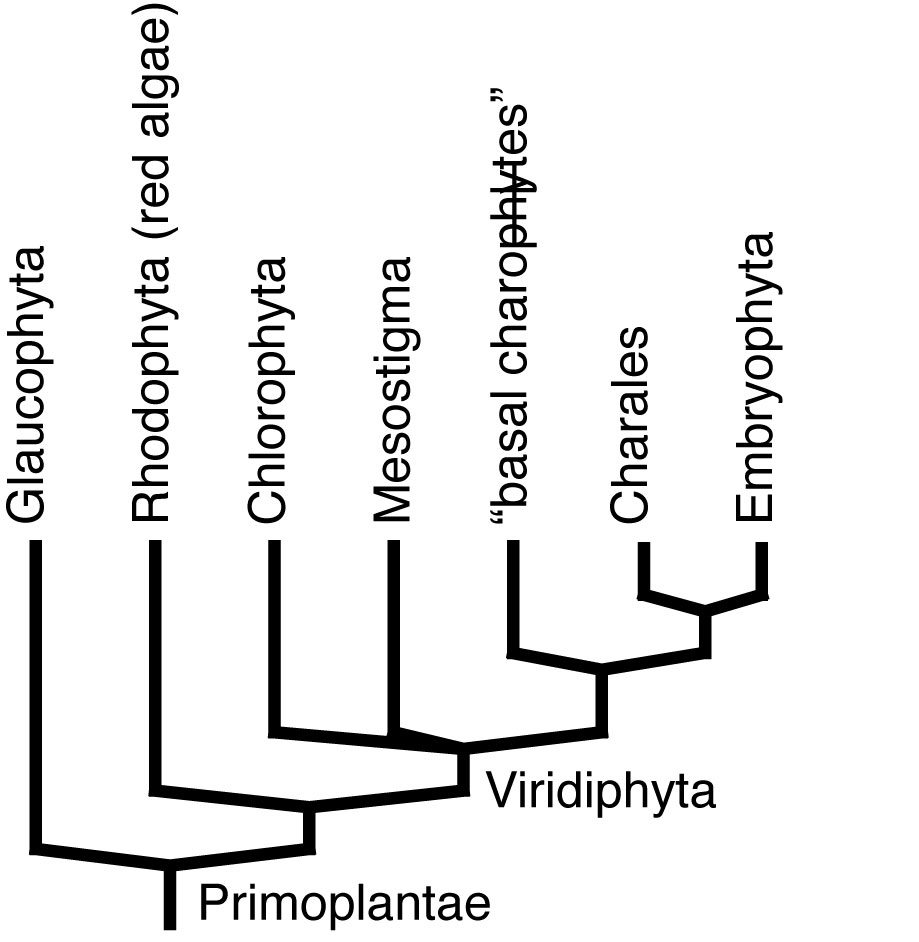
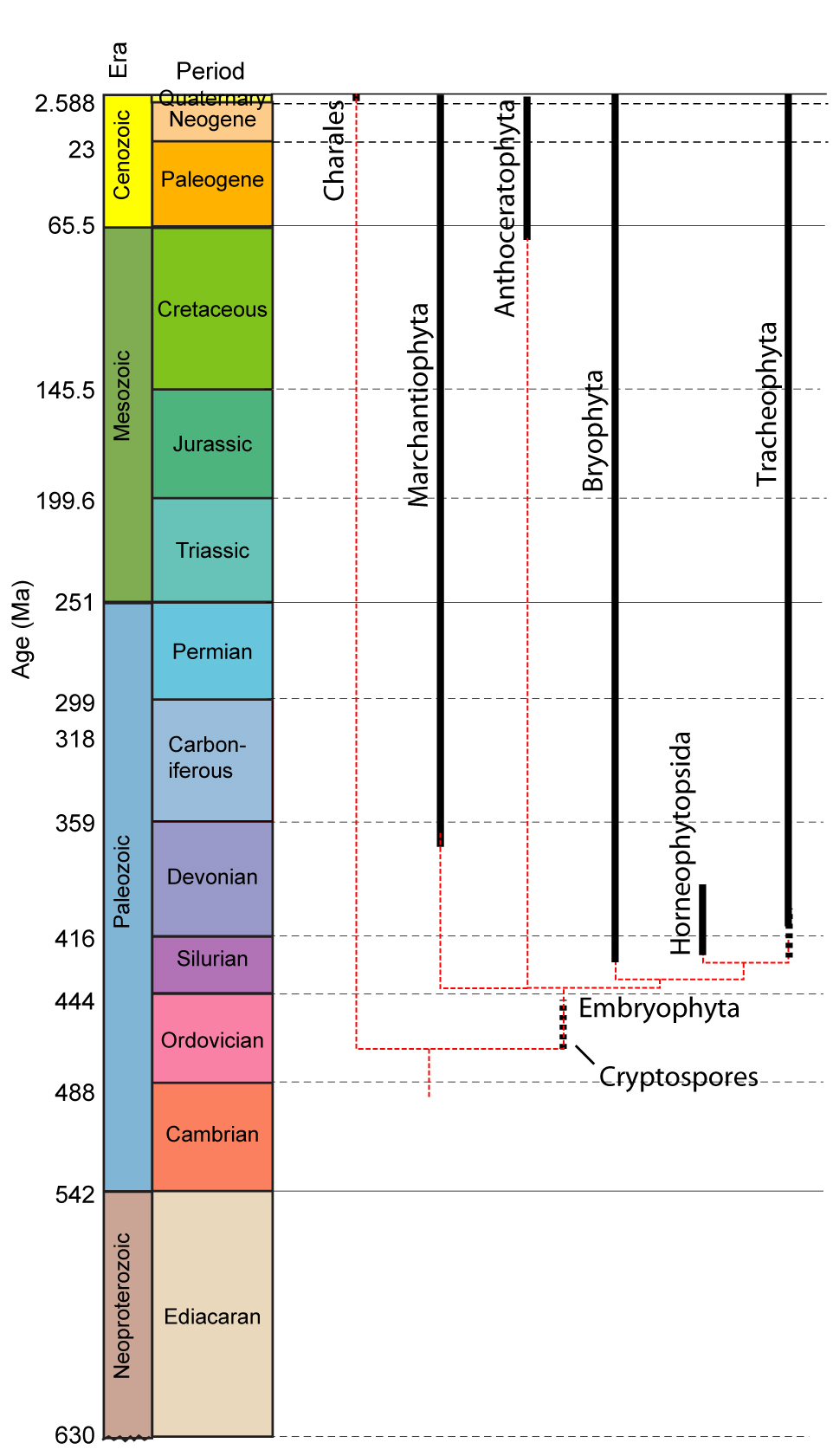
•Viridiphyta includes all multicellular plants. Originally aquatic without alternation of generations.
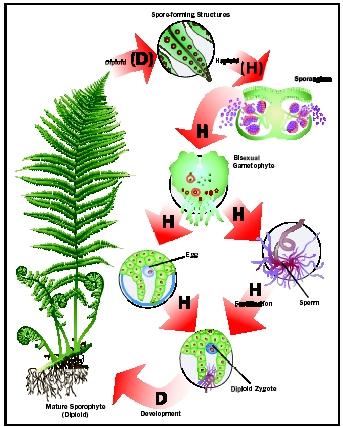
•Among Embryophyta - the clade of land plants, we see a range of adaptations to life on land, plus a struct alternation of diploid sporophyte and haploid gametophyte generations.
•Possible embryophyte body fossils occur as far back as the Cambrian, but their unambiguous record begins in the Silurian.
•Basal - bryophyte-grade plants emphasize the gametophyte generation.
•Tracheophyta - the vascular plants are the primary land plant radiation. They emphasize the sporophyte generation.
•Lignophyta - the seed plants - modify tracheophytes reproduction by retaining the female gametophyte within the body. When its ova are fertilized, the gametophyte becomes a seed.
•Anthophyta represents the radiation of plants with reproductive structures combining male and female components - flowers. Angiospermae is the crown group of living anthophytes.
"Land forests are the coral reefs of the ocean of air.."
(Steven Magee.)
We must put up with it.
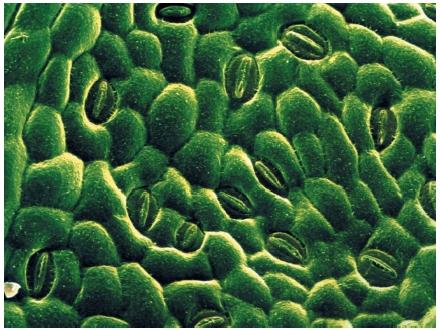
In order for plants to colonize the land, they had to overcome the challenges of retaining water, exchanging gasses, supporting their bodies, and reproducing out of water. Five adaptations facilitated their response:
Conventionally, we site the appearance of unambiguous plant macrofossils in the Early Silurian as the start of the land plant record. Gensel, 2008 has revealed microfossils including:
Monilophyta (ferns and horsetails): (Late Dev. - Rec.)
Ferns were the first plants with large vascularized leaves. Most, but not all ferns have very short trunks that support large pinnate leaves. Many ferns have independently evolved the "tree-fern" habitus, growing to the size of small trees. Post-paleozoic tree ferns seem to be monophyletic, but there were paleozoic ferns that separately evolved the tree-fern form. Ferns are plesiomorphic in bearing their sporangia on the bottoms of unmodified leaves.
 Equisetum arvense
Equisetum arvense
One monilophyte group has only recently been found to group within "ferns" - the Sphenopsida or "horse-tails. (Dev. - Rec). Like lycophytes, these plants protect their sporangia in strobili that develop at the end of stems. They are distinguished by the unique morphology of their stems. These consist of cylindrical sections separated by nodes. Whorles of slender leaves radiate from the nodes. During the late Paleozoic and Triassic, some sphenopsids such as Calamites attained tree size, but they were never the primary flora of their environments. Modern sphenopsids are known for rendering themselves inpalatable by incorporating silica phytoliths into their tissues.
Lignophyta - Seed Plants: (Dev. - Rec.)
Seeds: The second great radiation of land plants occurred during the Late Paleozoic, and was associated with the evolution of the seed. This occurred in stages.
- Ancestrally, gametophytes produced both sperm and ova.
- Some fossil as well as some recent non-seed bearing plants, display a dimorphism of between megaspores and microspores, which specialize in developing into gametophytes producing either ova or sperm, respectively and which resemble them in size dimorphism. This trend seems to develop primarily in plants with strobili which can, in principle, harbor the female gametophyte inside the sporangia.
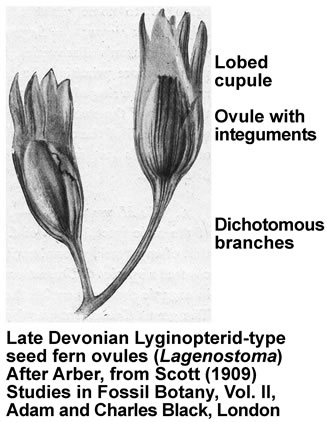
- When we pick up this trend again, it is in plants that retain the female gametophyte inside special reproductive structures. The gametophyte is now known as an ovule, and contains the ovum, nutritive tissue, a seed coat, and structures for conducting sperm to the ovum. Once the ovum is fertilized, the ovule becomes a seed.
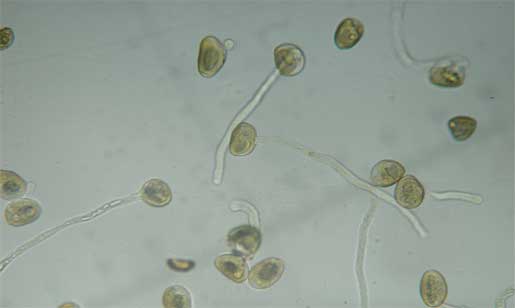
- How does the sperm get to the ovum? When a microspore lands on the sperm-capturing portion of the ovule, it develops into a specialized male gametophyte called a pollen-tube which grows into the ovule. Sperm is conducted down the pollen-tube to the ovum. In such a set-up, we call the microspore a pollen grain.
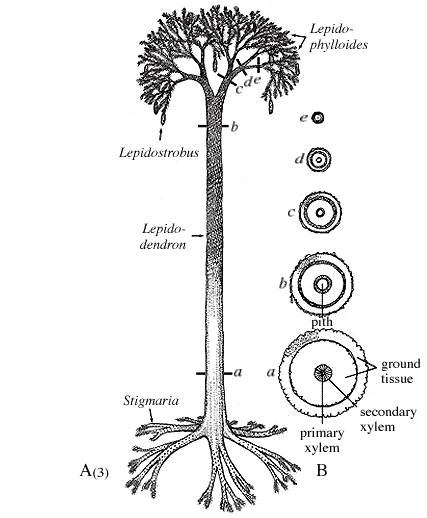
Wood: A second major evolutionary novelty was the ability of seed plants to lay down secondary tissue. Ancestrally, land plants could create new tissue only in the apical meristem tissue at the tips of their stems. In seed plants, however, we see the appearance of vascular cambium, a sheath of tissue that generated new vascular tissue around the circumference of the stem. New tissue laid down by vascular cambuim appears as growth rings in cross sectioned stems. The ability to thicken existing stems allowed seed plants to attain greater size than arborescent lycopods.
Lignophyte diversity - a sampling:
"Progymnosperms" Paraphyletic grade group of plants bearing vascular cambium but lacking ovule-like reproductive structures:
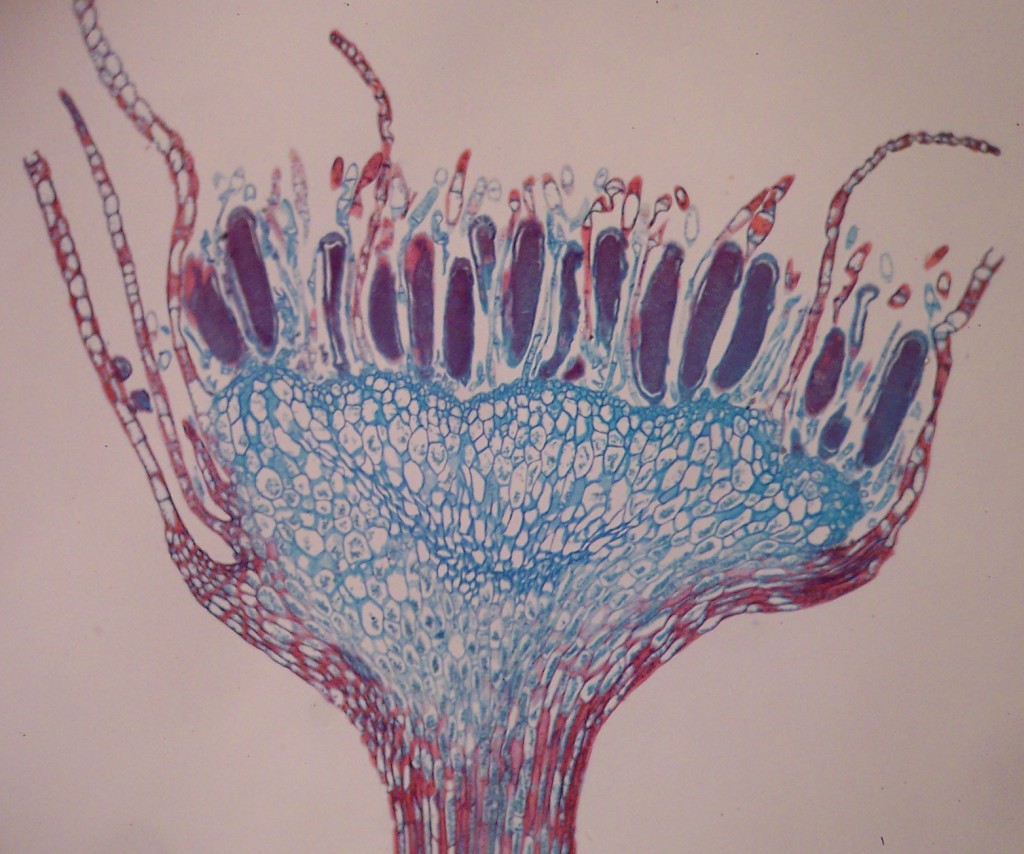
- Archaeopteridales: (Devonian - Early Carboniferous) Archaeopteris, is typical. Major component of the first forests of the Late Devonian, including the first large (redwood-sized) trees. Ovules and pollen organs were absent, but displays distinction between mega and micro spores. Note: Forests of Archaeopteris and its kin lined the banks of waterways inhabited by the likes of Acanthostega.
 Runcaria
Runcaria reproductive structure from
Wikipedia
- Runcaria: (Middle Devonian). Preserved megasporangia suggests that megaspores were wind pollinated, but show no seed coat or means to conduct the pollen tube to the megaspore/ovule. An early stage in seed evolution.
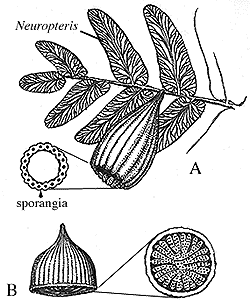
"Seed ferns" Paraphyletic grade group including numerous Late Paleozoic land plants characterized by distinct ovules and pollen organs born on separate leaves. The leaves were typically like those of ferns.
A rogue's gallery of seed fern groups:
- Lyginopterida: (Dev. - Carb.) Vine-like plants (possibly epiphytes) bearing ovules and pollen organs on separate leaves. typically each ovule theca contained several ovules.
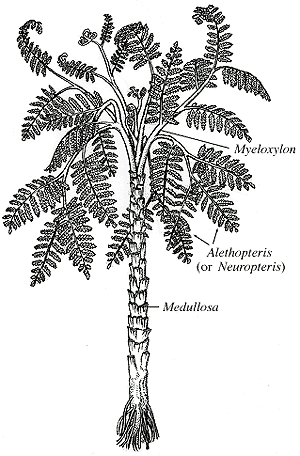
- Medullosales: (Late Paleozoic) Superficially fern-like plants bore distinct ovules (one per theca) and pollen organs on separate leaves.
- Callistophytales: (Carb. - Perm.) Vine or shrub-like plants also bearing ovules and pollen organs on separate leaves.
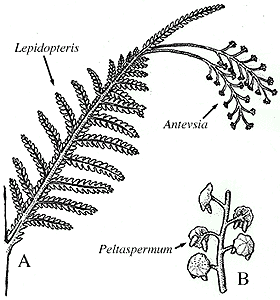 Lepidopteris
Lepidopteris and
Peltaspermum
- Peltaspermales: (Carb. - Tri.). Seeds occur on the lower surface of an umbrella-like organ.
- Corytospermales: (Permian - Paleogene) Includes Dicroidium (right) a major component of Triassic Gondwanan flora. The form taxon Umkomasia - short branches terminating in ovule-bearing cupules - appears to be the fruiting organ of Dicroidium.
Seed plants did well enough with separate pollen organs and ovule-protecting fruiting bodies for the late Paleozoic and most of the Mesozoic. Many living seed plants still use this method (E.G. the pollen organs and ovulate organs of pines.) And yet, during the Mesozoic, one group evolved toward their consolidation in a single reproductive structure - the flower. We follow that story in the next lecture.
Seed review:
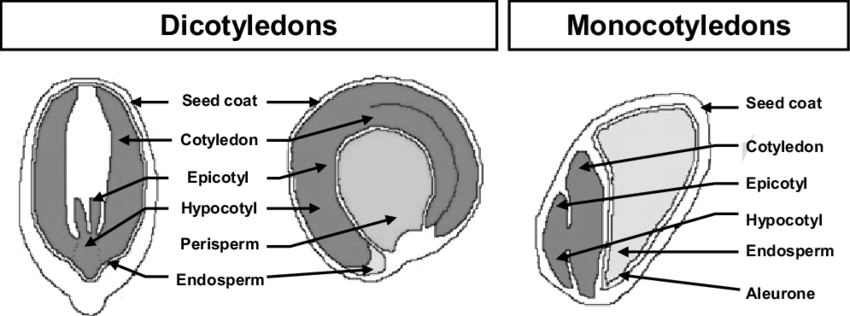
- The ovule: The derived haploid female gametophyte is retained inside a special structure of the sporophyte, producing ova.
- Fertilization by sperm produces a zygote that develops into an embryo.
- After fertilization, the ovule develops into a seed, and haploid cells of the ovule develop into "endosperm", nutritional tissue for the embryo.
Ancestrally, the ovule develops on the surface of a specialized leaf, such as the cone scales of living acrogymnosperms.
When we get past the paraphyletic grade group of "seed ferns," Lignophyta contains two major extant groups:
- Anthophyta: Flowering plants and their kin
- Acrgymnospermae: The traditional - "Gymnospermae" (naked seed) - non-flowering seed plants.
Acrogymnospermae: (Devonian - Quaternary) Deemed paraphyletic in earlier literature, the traditional "gymnosperms" have emerged from recent molecular analyses as a monophyletic group. An interesting fact for its own sake, but also because it implies that the evolutionary "roots" of flowering plants go back to the mid-Paleozoic.
 Dioon spinulosum
Dioon spinulosum
Cycadophyta: (Cycads) Permian - Quaternary Shrub-sized superficially fern or palm-like plants with short trunks and very tough pinnate leaves. They are very slow-growing and long-lived. Like conifers, they produce male and female strobili. In this case, however, the plants are dioecious i.e., each individual plant is fully male or female, producing cones of only one sex. Cycads were very common understory plants during the Mesozoic but are uncommon today.
 Ginkgo biloba
Ginkgo biloba
Ginkgoales (Ginkgos) Permian - Quaternary Common in the Mesozoic and Paleogene, they are now reduced to a single species, Ginkgo biloba. Like cycads, ginkgos are dioecious - have separate male and female individuals. The ovule is enclosed in a malodorous fleshy capsule.
Represented by a single species, Ginkgo biloba, but more speciose during the Mesozoic. Ginkgoes began to decline during the Cretaceous and were significantly reduced by the end of the Paleocene. (Possibly because the animals on which they depended to disperse their seeds had also declined.)
With the exception of a small region of southwestern China, ginkgoes are extinct in the wild. Thankfully, they are common under cultivation.
 Pinus nigra
Pinus nigra
Pinophyta: (Devonian - Quaternary) carry their seeds on the scales of cones. Most conifers are tree-sized and produce both male (pollen bearing) and female (seed bearing) cones. In many cases, their leaves are reduced to needles, allowing them to withstand cold and dry conditions. This ability allows many to retain their leaves year-round. Earliest known representative belonged to "Cordaitanthales" (Devonian - Jur.) Paraphyletic. Trees with whorls of strap-like leaves superficially like modern Pandanus.
Crown-group Pinophyta includes of the Pinales (Carboniferous - Quaternary) Important canopy trees of late Paleozoic and Mesozoic forests. Representatives include:
A fourth group requires notice:
Gnetophyta: (Ephedra, Welwitschia, and Gnetum.) Triassic-Quaternary. Weird arid adapted plants with flower-like strobili that enclose the ovule in an integument that is open at the end.
Until recently, we would have presented Gnetophyta as close relatives of flowering plants, however strong recent molecular evidence indicates that they are actually nested within Pinales - counterintuitive but strongly supported.
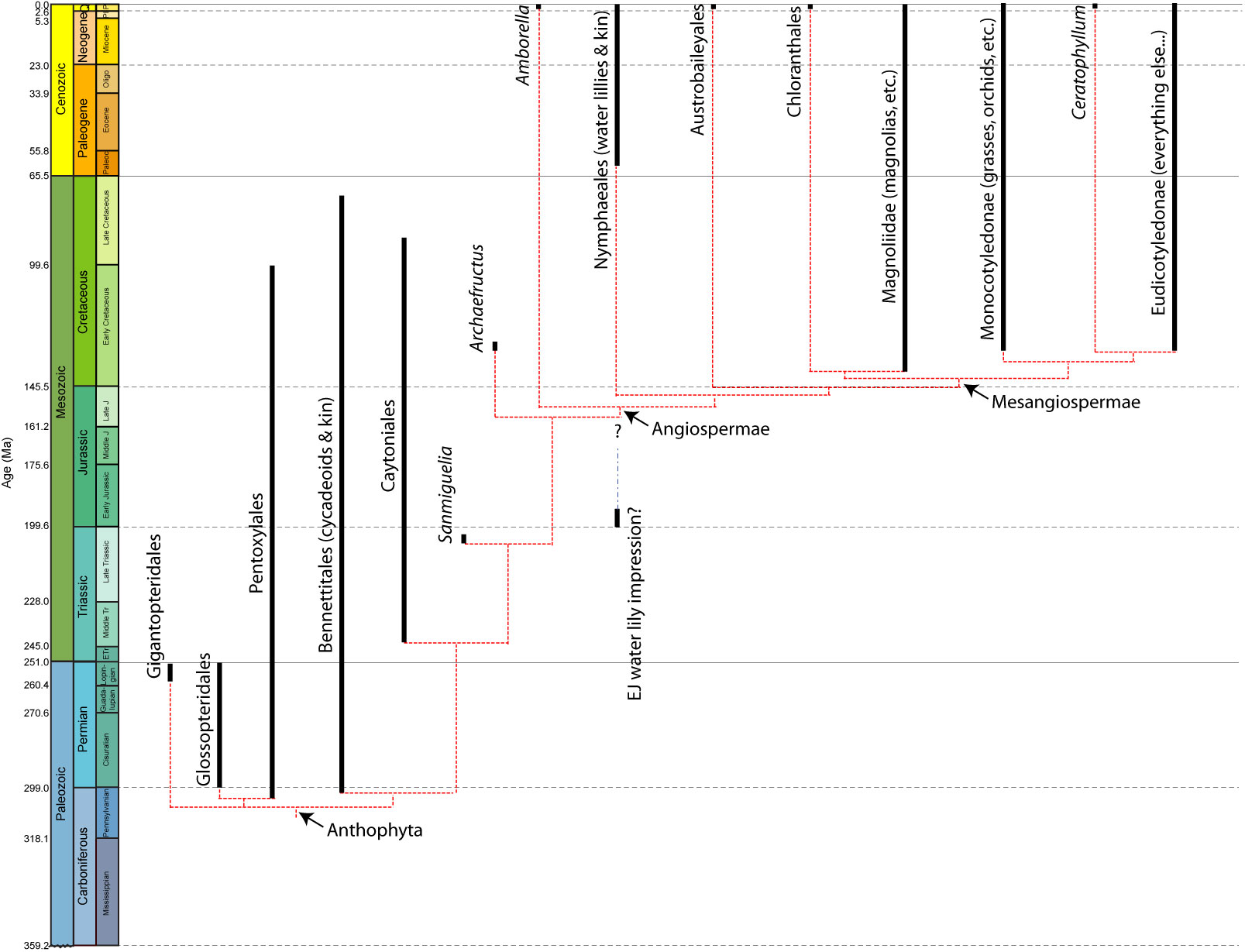
Anthophyta: Carboniferous - Quaternary) The total group of flowering plants and their kin. The familiar term Angiopspermae refers to the crown-group.
Before addressing the evolution of the innovations, of flowering plants, best to have a sense of where we are headed. Simply to say that flowering plants are distinguished by flowers would be sort of dumb. In fact, flowers represent three major innovations:
- Unlike other seed plants, in which pollen and ovule producing structures are completely separate, flowers combine them in a single complex structure.
- The pollen-organ is the stamen
- The ovulate organ is the pistil, a composite containing:
- One or more carpels - the specialized leaves from which the ovules are suspended (homologous with the scales of a conifer ovulate cone.)
- The stigma, to which pollen grains attach
- The style, with guides the pollen tube to the ovule.
- Perianth, the circle of specialized leaves (petals and sepals) inclosing the pistil and stamens.
- Unlike in other seed plants, the ovule is completely concealed by the carpel. When the ovule is fertilized, the carpel of many species swells to form a fruit, whose primary purpose seems to be to entice animals into consuming the seed. This allows the seed to hitch a ride in the animal's gut and insures that it will be deposited with a blob of fertilizer. Indeed, some seeds can't germinate without first passing through someone's gut.
- The specialized leaves (petals, sepals) of the perianth along with secretions of nectar function to attract pollinators, usually insects. In the latter two adaptations, the plant "pays" animals to transport pollen directly to other flowers, and to disperse seeds, resulting in an increase in reproductive efficiency.
-
But that is tame. The angiosperm seed is derived in that the endosperm - the tissue that nourishes the embryo - forms from the fusion of sperm with a special diploid cell of the ovule. After fertilization, this triploid cell develops into the endosperm. (Think of it as if everyone shared the womb with a fraternal twin whom they were intended to eat.) In non-anthophytes, this nutrient tissue is haploid.
 Nymphaea
Nymphaea sp.
- Angiosperms tend to have broad leaves with branching venation, in which the veins show multiple orders of branching and cross veins.
Anthophyte Evolution:
-
 Delnortea abbottiae
Delnortea abbottiae
Gigantopteridales: (Permian) Major portion of Late Paleozoic and Triassic Gondwanan flora. These bore distinct ovulate and stamenate reproductive structures on separate specialized leaves - the ancestral lignophyte pattern. Were derived in their development of broad non-pinnate leaves.
-
Glossopteridales: (Permian - Triassic) Major portion of Late Paleozoic and Triassic Gondwanan flora. These, again, bore distinct ovulatte and stamenate reproductive structures on separate specialized leaves. Note, the Permian Glossopteris was a particularly significant member of the Gondwanan flora.
-
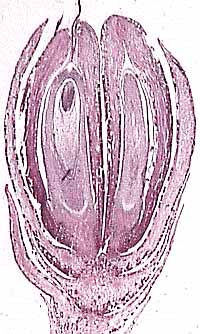
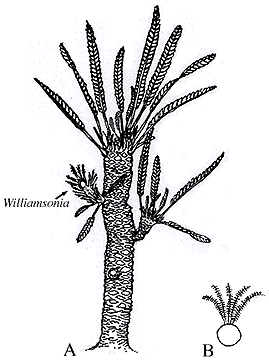
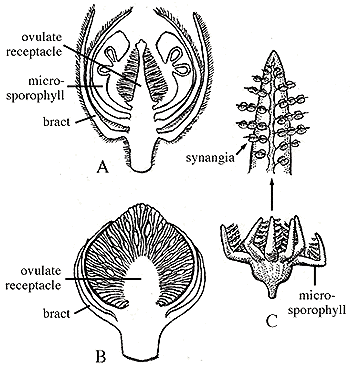
Bennettitales: (Triassic - Cretaceous) (Sometimes called "Cycadeoids.") Superficially resembling cycads with short trunks covered by leaf scars and pinnate leaves. Their reproductive structures are different. First, like most flowering plants, they are hermaphrodites. Second, some display flower-like cones in which the ovule is contained in a central recepticle surrounded by specialized leaves (bracts) that might be homologous to petals. In some, microspore (pollen) producing structures are arrayed radially around it, like stamens - a truly flower-like arrangement. Alas, not all bennettitaleans show this, so this bisexual reproductive structure seems to be convergent with angiosperms.
-
Sanmiguelia lewisii: (Triassic) Described by Cornet 1989 as having:
- Separate stamenate and ovulate reproductive structures in which the latter were very similar to those of angiosperms.
- Leaf venation patterns similar to those of angiosperms.
Some have put Sanmiguelia forward as "the earliest angiosperm." Probably not, but it is getting very close.
-
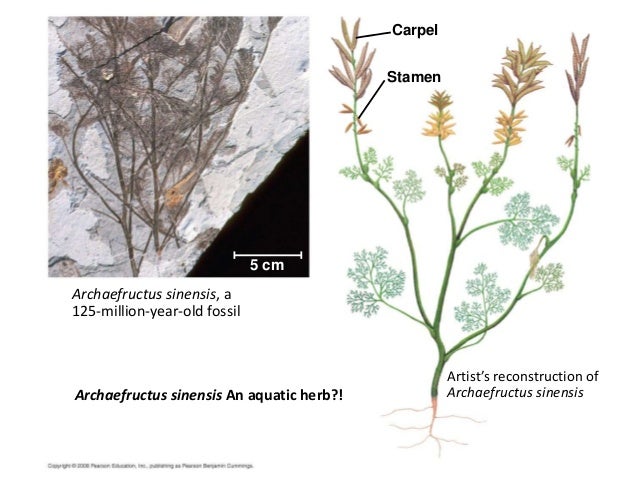
Archaefructus eoflora: (Early Cretaceous): An aquatic plant. In contrast to Sanmiguelia which seems to have male and female structures on distinct branches, Archaefructus eoflora is described by Ji et al., 2004 as having a single enflorescences (branches covered with small reproductive structures) in which:
- the distal structures were stamenate
- the proximal ones were ovulate
- But where they met were a few rows of composite bisexual structures.
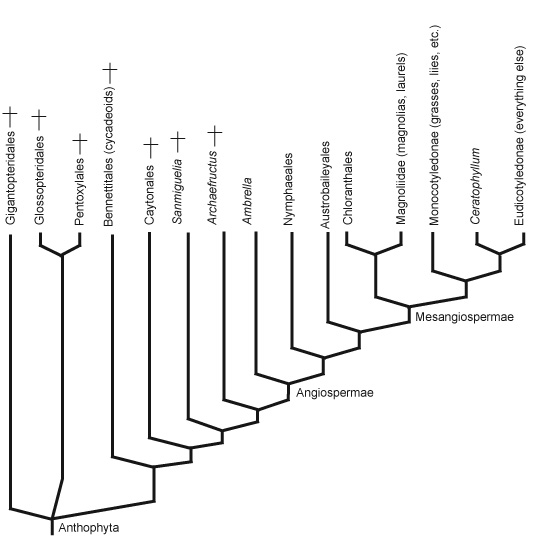
Angiospermae:
(Cretaceous - Quaternary) The crown group of living anthophytes.
Angiosperms appear unambiguously in the fossil record in the Early Cretaceous. Today, they are the dominant plant type, with over 200,000 known species (compared to 550 species of gymnosperms).
Both the ecologies of their fossil relatives Sanmiguelia and Archaefructus, and of their basal living members suggest that they inhabited wetlands.
Angiosperm partners:
We've noted the big angiosperm innovation - the use of animal commensalists to pollinate flowers and disperse seeds. Who were these? Some speculation:
Fruit eaters:
Arthropods might like to eat the fleshy parts of fruit, but they destroy them in the process. Plants develop fruit to select for something that can eat the whole fruit and spread the seeds. During the Mesozoic, that could only be a vertebrate.
- Herbivorous mammals and squamates of the Early Cretaceous were mostly small, but could have eaten berry-sized fruit.
- Herbivorous crocodyliforms were rare, but did exist
- But herbivorous dinosaurs were quite common, and could easily eat and transport a plenty of fruit! Since most early angiosperms were built low to the ground, it would be small and/or low-browsing forms that would have been the main agents of transportation for fruit.
Flower sniffers:
- No Early Cretaceous archosaur shows nectar-eating adaptations.
- Mammals were small, and may have been pollinators, but bats - the great modern mammalian pollinators - did not exist.
- Insects (including beetles, dipterans (flies), lepidopterans (butterflies and moths), and hymenopterans (wasps, bees, and ants)) were present, and were almost certainly the main pollinators in the Mesozoic just as they are today.
- In fact, the major groups of holometabolous insects that are the major pollinators today tend to diversify roughly contemporaneously with angiosperms ()Hu et al., 2008).
Was anyone else on the scene to fill this role? In fact, mecopteran insects (today represented by scorpionflies) appear to have served that role according to Ding , 2014. Rather than being predators, like the living members, fossil Cretaceous scorpionflies did not have biting jaws, but instead butterfly-like probing snouts (right). Similarly, Khramov and Lukashevich, 2019 describe a Jurassic dipteran (fly) with an improbably long proboscis, probably also a pollinator.
Angiosperm diversity:
 Amborella trichopoda
Amborella trichopoda male flower from
David Tng
The most primitive living angiosperm:
- Amborella trichopoda most primitive. Although weird for being dioecious (plesiomorphy?), otherwise suggests the structure of the ancestral flower:
- very small
- petals and sepals undifferentiated
- ovum development resembles that of nonflowering plants
Additional Reading:
- Bruce Cornet, 1989. The reproductive morphology and biology of Sanmiguelia lewisii, and its bearing on angiosperm evolution in the Late Triassic. Evolutionary Trends: In Plants 3(1)..
- He Ding, Chungkun Shih, Alexei Bashkuev, Yunyun Zhao, Dong Ren, 2014.
The earliest fossil record of Panorpidae (Mecoptera) from the Middle Jurassic of China. ZooKeys. 431: 79–92..
- Patricia Gensel, 2008 The earliest land plants. Annual Review of Ecology, Evolution, and Systematics
Vol. 39: 459-477.
- Shusheng Hu, David L. Dilcher, David M. Jarzen, and David Winship Taylor, 2008. Early steps of angiosperm–pollinator coevolution. Proceedings of the National Academy of Sciences 105 (1) 240-245
.
- Ji, Qiang; Li Hongqi; Bowe, L. Michelle; Liu Yusheng;
and Taylor, David Winship , 2004. Early Cretaceous Archaefructus eoflora sp. nov.
with Bisexual Flowers from Beipiao, Western Liaoning, China. Acta Geologica Sinica 78(4) 883–896.
- Alexander V.Khramov and Elena D.Lukashevich, 2019.
A Jurassic dipteran pollinator with an extremely long proboscis. Gondwana Research
71, 210-215.
- Robin B. Kodner and Linda E. Graham, 2001.
High-temperature, acid-hydrolyzed remains of Polytrichum (Musci, Polytrichaceae) resemble enigmatic Silurian-Devonian tubular microfossils. American Journal of Botany
88(3):462-466.
























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}