


Key Points:
•True multicellularity involves specialized cell types, the specialization of reproductive cells, and apoptosis.
•Possible multicellular eukaryotes may extend back to 2.2 Ga
•Multicellularity arose five times among eukaryotes.
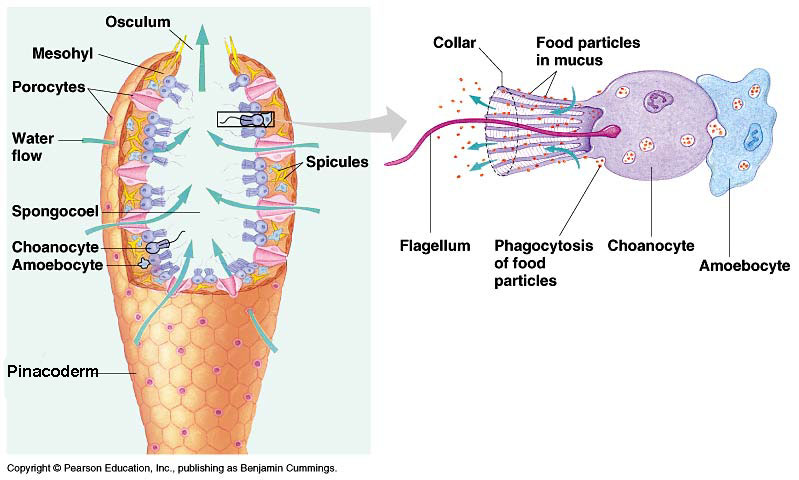
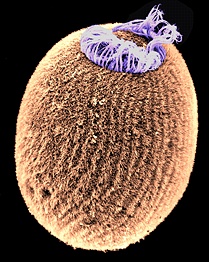
•Porifera (sponges) are the most primitive grade of metazoan.
•There are three major poriferan taxa (Hexactinellida, Demospongea, Calcarea) and one oddball (Homoscleromorpha).
•There is no strong consensus on whether poriferans are monophyletic.
•The earliest potential sponge fossils are Ediacaran but geochemical evidence for them goes back to 1.8 Ga
•Several enigmatic Phanerozoic fossil groups are likely to be poriferans.
"Well, it's not secret that the best thing about secrets is telling someone else your secret, thereby adding another secret to your secret collection of secrets, secretly.
(SpongeBob Squarepants, Season 2, Episode 35a.)
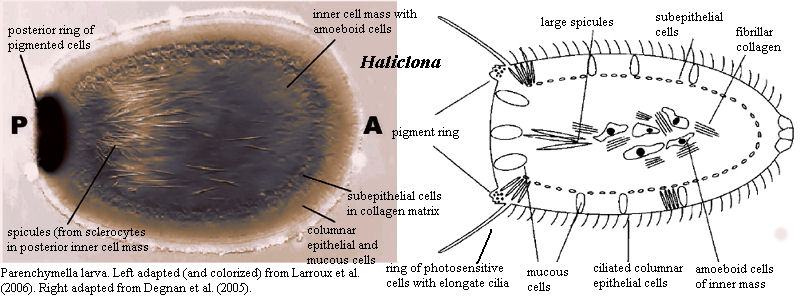
Most of the semester remaining will address the fossil record of Metazoa (animals.) Metazoans are one of five groups of multicellular eukaryotes to arise during the Neoproterozoic. Unlike simple cell colonies like Volvox or slime molds (right), each of these groups forms organized bodies in which:
The earliest fossil evidence for colonial eukaryotes comes from the Proterozoic:
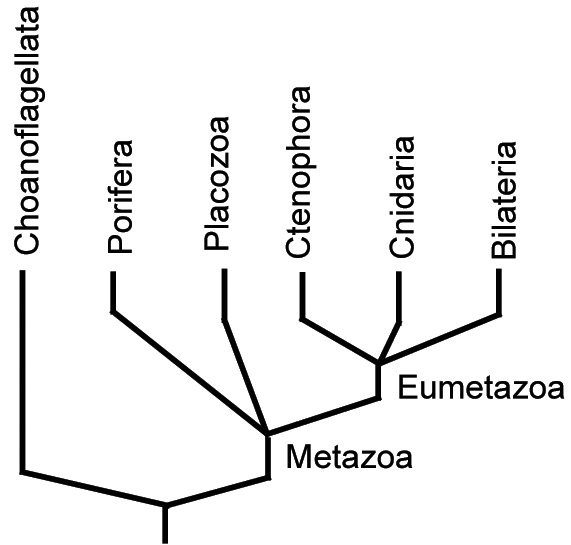
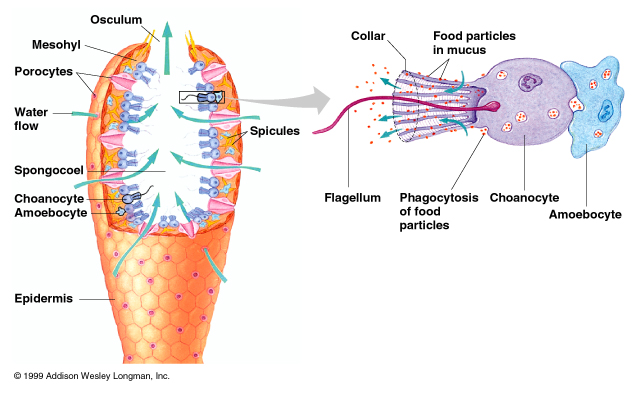

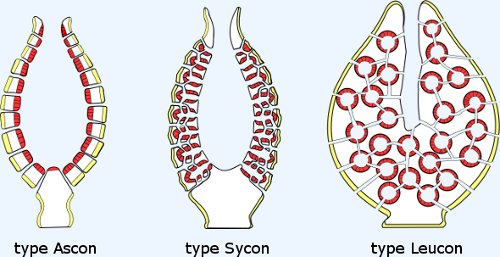
Diversity: Numerous groups. Of special interest:

Diversity: Too great to explore here, but likely to include some oddballs such as:
Hexactinellida: (Ediacaran - Quaternary)
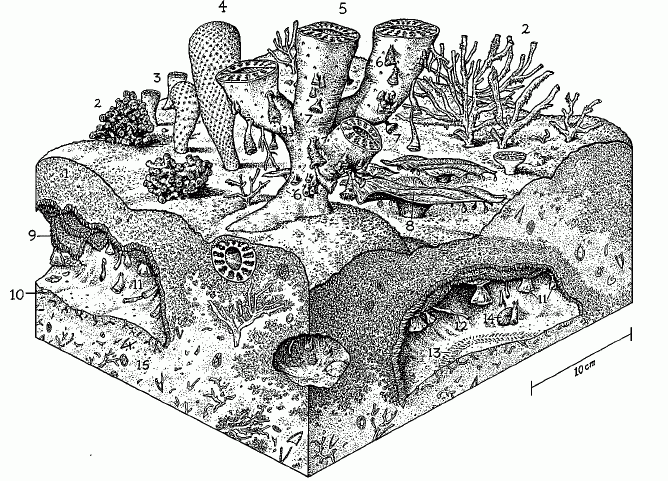
Ecology:
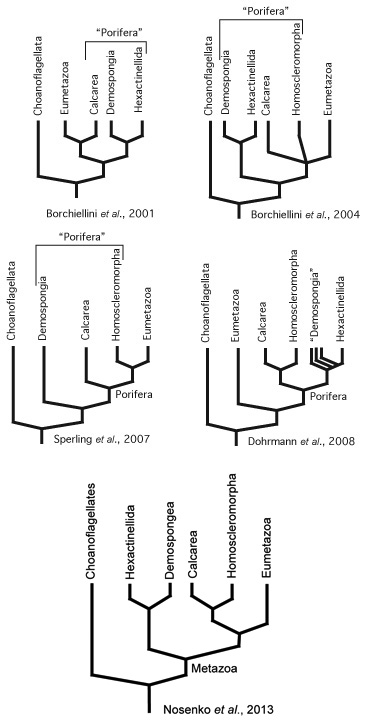
An unsettled question. Complicating factors include:
The techniques of molecular phylogeny have yielded ambiguous results. For example:
Given the morphological similarity of Homoscleromorpha to some demosponges, it is odd that molecular phylogenies consistently find Homoscleromorpha close to Calcarea with whom they lack morphological or cytological synapomorphies and pair Demospongea with Hexactinellida.
This issue is far from settled. Stay tuned!
Before we get too excited, Antcliffe et al. 2014 reject these claims. Their conservative approach identifies the earliest sponges as hexactinellids from the earliest Cambrian of Iran.

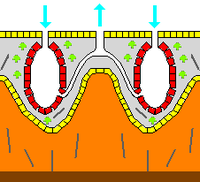
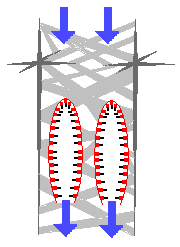
Characteristics:
Ecology:
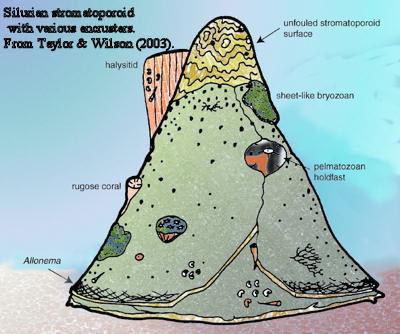
Stromatoporoidea: (Ordovician - Quaternary?): Monophyletic group or polyphyletic assemblage of Elvis taxa? Stromatoporoids, massive calcareous encrusting organisms, were important Paleozoic reef-builders that require our attention as primary organisms of mature reefs of the Silurian and Devonian.. Note: Don't confuse with stromatolites, even when they occur together.
Characteristics:


Before you conclude that all enigmas have been resolved...
Archaeocyatha: (Early - Middle Cambrian). Considered "poriferan grade" based on general body plan, but there is no consensus on their phylogeny. Reitner, 1990 found them to be nested within Porifera. If "Porifera" is paraphyletic, then GORK. At best, we assert that they branch close to, if not within Porifera.
Ecology:
Evolutionary trends:
A Final Thought:
I withheld one hypothesis of poriferan phylogeny - the minority view of Maldonaldo, 2004, who regards choanoflagellates as being derived from Demospongea through a process of paedomorphic simplification. Sounds strange, but is based in part on the observation that some choanoflagellates are capable of secreting internal structures of amorphous silica and the quite reasonable argument that in sponges, the ability to secrete silica was probably derived secondarily from the secretion of calcite.
Is it insane? Our aesthetic of evolutionary progress compels us to view choanoflagellates as precursors to poriferans, but nature isn't obliged to cooperate. Maybe Proterospongia is a drastically simplified poriferan, and choanoflagellates are choanocytes that have shaken free of their association with other cells. More data, please.
To Next Lecture.
To Previous Lecture.
To Syllabus.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}