
Vertebrata I: Paleoichthyology
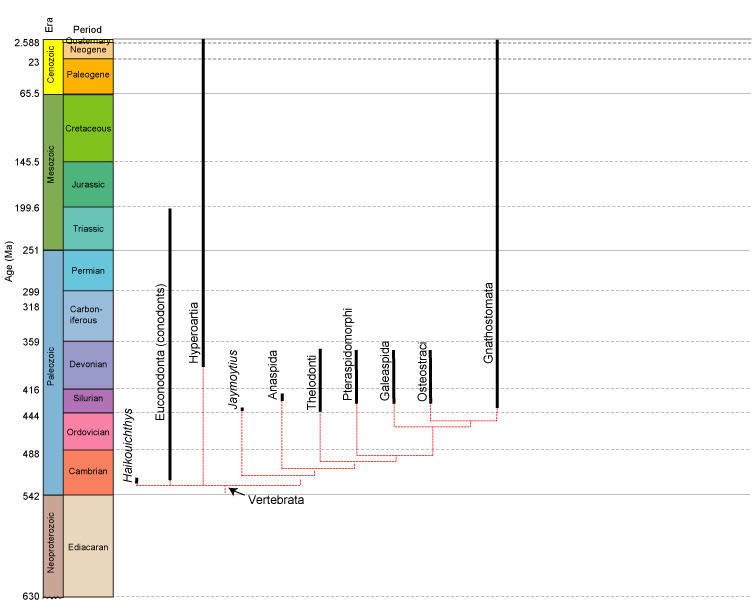 The evolution of Paleozoic vertebrates presents a paradox:
The evolution of Paleozoic vertebrates presents a paradox:
We have such a copious record of heavily armored Early Paleozoic forms that it is tempting to forget that the group's most basal members (like Haikouichthys and Myllokunmingia) essentially lacked hard tissues. Indeed, the early evolution of Vertebrata is marked by the diversification of bony tissues and their proliferation through the body. This pattern was recently illuminated by Sansom et al., 2010. (Synopsis at right.) So, we start with a review of bony evolution.

Bones
Hagfish and lampreys, as the only living jawless vertebrates, provide an interesting glimpse of early vertebrate evolution, however they lack the proper hard tissues by which we know the vast diversity of early vertebrates - bone.
Fossil vertbrates are mostly known from hard tissues - bone and teeth. Bone is composed of:
- A mineral component - made of calcium phosphate (i.e. the mineral hydroxyapatite).
- A protein component - made mostly of the fibrous protein collagen.
Among living craniates, bone in an form only occurs among members of Vertebrata - craniates with vertebral elements protecting their spinal cords. NOTE: although we tend to get sloppy, strictly speaking, Hyperotreti (hagfish) are considered the sister taxon of Vertebrata but not members of it. What does the study of fossil organisms tell us about the distribution of bony tissue?

A rogue's gallery of early Paleozoic vertebrates:
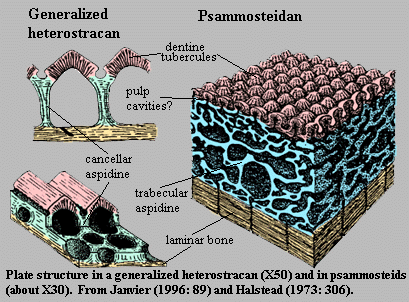
The earliest vertebrate hard tissues are small acellular elements: conodont elements, which show outer layers of enamel covering layers of dentin. Conodonts were not the only representation of craniate hard tissues in the Cambrian, however. Enigmatic, scale-like plates of bony armor called Anatolepis were also present. In this and similar creatures, histologically tooth-like denticles complete with enamel and dentin formed a composite superficial body armor.
Indeed, in many early vertebrates, there seems to have been little difference between teeth and scales, which took the form of little denticles with a pulp cavity, dentin, and enamel.
The most basal vertebrates, however, lacked any hard tissues (except for conodont elements.) A survey of early vertebrate evolution should focus on their acquisition:
Euphanerops (right) and Jamoytius (Silurian)

The phylogeny of Sansom et al., 2010 was made possible by reexamination of these important early vertebrate, often previously cited as the ancestral vertebrate or close to Hyperoartia. It seems to represent the first appearance of hard tissue elements outside the mouth: W-shaped bony acellular scales - composites of dentin and enamel.
Indeed, in many early vertebrates, there seems to have been little difference between teeth and scales, which took the form of little denticles with a pulp cavity, dentin, and enamel. A survey of the diversity of fossil jawless vertebrates tracks the proliferation of different bone types in different parts of the body.
Anaspida (Silurian)
Morphology:
- Hard tissue in the form of acellular plate-like scales, reminiscent of those of Jaymoutius, however these extend onto the head.
- Retain single median nostril (reminiscent of lampreys).
Overall, anaspids seem adapted for active swimming. Exactly how they ate is mysterious, but they lack obvious adptations to suspension feeding or to taking large prey.
Synapomorphy of Anaspida and jawed-vertebrates: Dermal skeleton of head.
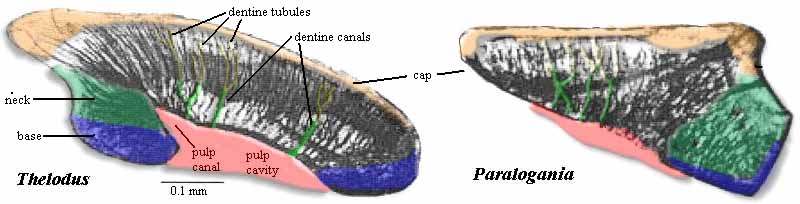
Thelodonti (Ordovician - Devonian)

Morphology:
- Hard tissue Entirely consists of small scales that usually disarticulate when the animal dies. These scales are distinctive, consisting of enamel and dentin layers around a pulp cavity, like a vertebrate tooth. Note: from this point on the tree onward, aquatic vertebrates generally retain scales of this sort or their derivatives, regardless of any other kind of skeletal ossification they may have.
Living chondrichthyans preserve a similar pattern.
Synapomorphies of Thelodonti and jawed-vertebrates:
- Dermal denticles with distinct root, crown, and pulp-cavity.
- Paired nostrils and nasal capsules.
Issue: True Bone:
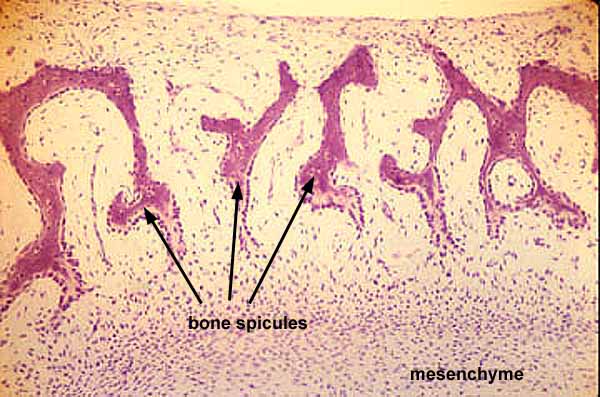
Before proceeding, a note: The cells that secrete and maintain hard tissue may be locked within it, yielding cellular bone. Seen in larger bony elements. Cellular bone forms in two ways:- Dermal bone: Laid down as a two-dimensional membrane. (E.g. human cranial bones). Ancestrally these formed near the body surface, but in derived vertebrates, their derivatives may invade deeper parts of the body.
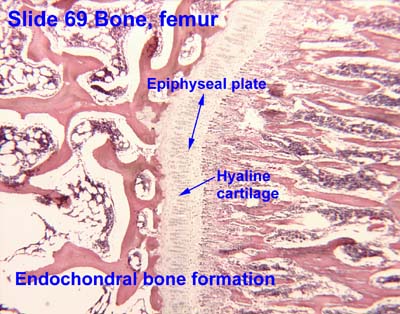
Endochondral bone cross-section from University of Oklahoma Health Sciences Center
Interactive Histology Atlas - Cartilage bone: Three-dimensional bone that is preformed in cartilage. (E.g. human limb bones). The cartilage, in turn, is preformed by condensations of mesenchyme cells, amoeboid mesodermal connective tissue of the embryo. (Remember them?) Cartilage bone makes its evolutionary debut in the skeleton of the braincase but is widespread in vertebral columns and the skeleton of the limbs.
However cartilage bones in living vertebrates actually form in two ways: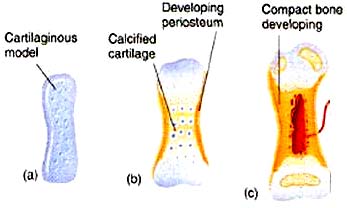
- Perichondral ossification: Bone formation begins at the perichondrium - the membrane surrounding the cartilage precursor.
- Endochondral ossification: Bone formation begins in the interior of the cartilage precursor.
Note: Among living vertebrates, both types of ossification occur in cartilage bone, so neontologists and human anatomists call cartilage bone "endochondral bone." Paleontologists must distinguish them, however, because perichondral ossification evolved first and many fossil vertebrates have perichondrally ossified bone without the endochondral component.
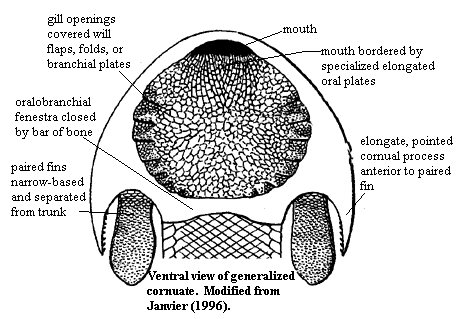
Pteraspidomorphi (Ordovician- Devonian).

The earliest well-preserved vertebrate, the Ordovician form Sacabambaspis, ironically represents a more derived form of hard tissue, in which individual denticles are integrated into broad head-shield composite elements and joined to one another through dermal layers of aspidin, a composite of thelodont-like denticles, lamina of dentin, and cellular dermal bone. These shields are the first vertebrate elements that we can call proper bone. these elements seem to have played the roles of:
- Armor
- A reservoir of calcium and phosphate for use by the animal's metabolism.
Morphology:
-
- The mouth is opened and closed by row of narrow parallel plates in the lower "lip."
- Impressions of otic capsules on the underside of the dorsal shield indicate the presence of two sets of semicircular canals - similar to lampreys.
- Paired nostrils
- The mouth is opened and closed by row of narrow parallel plates in the lower "lip."
Synapomorphy of Pteraspidomorphi and jawed-vertebrates:
- Plate-like cellular dermal bone.
Galeaspida: Restricted to southern China and Indochina, then a separate continent. (Silurian - Devonian)

Morphology:
- large flat head shield enclosing an perichondral bony braincase. The head shield is a composite of the perichondral braincase and composite dermal bone like that of pteraspidomorphs.
- large opening in upper front of head shield (maybe a large median nostril) leads to paired olfactory capsules and pharynx
- mouth is ventral
- Pharynx is large with many gill openings, suggesting suspension feeding.
Synapomorphy of Galeaspida and jawed-vertebrates: Perichondral bone in braincase.
Osteostraci (Silurian - Devonian): Resemble galeaspids but with differences:

Morphology:
- large head shield enclosing an perichondral bony braincase. As in Galeaspida, the head shield is a composite of the perichondral braincase and composite dermal bone like that of pteraspidomorphs.
- Head shield contains large sensory fields.
- Single median nostril leads to a single olfactory pouch (like in lampreys) and doesn't communicate with the pharynx.
- Notochord invades upper lobe of tail.
- Dorsal fin
- But the big thing: Paired pectoral fins (complete with endochondral skeleton elements and muscle) present.
Synapomorphies of Osteostraci and jawed-vertebrates:
- Paired pectoral appendages
- Heterocercal tail
- Distinct dorsal fin
Coping with history: You will encounter the term ostracoderms in the literature. This referred to the vast paraphyletic grade-group of armored jawless vertebrates whose diversity we have surveyed.
Gnathostomata: (Sil - Rec.) The jawed vertebrates. Quantum leap forward.

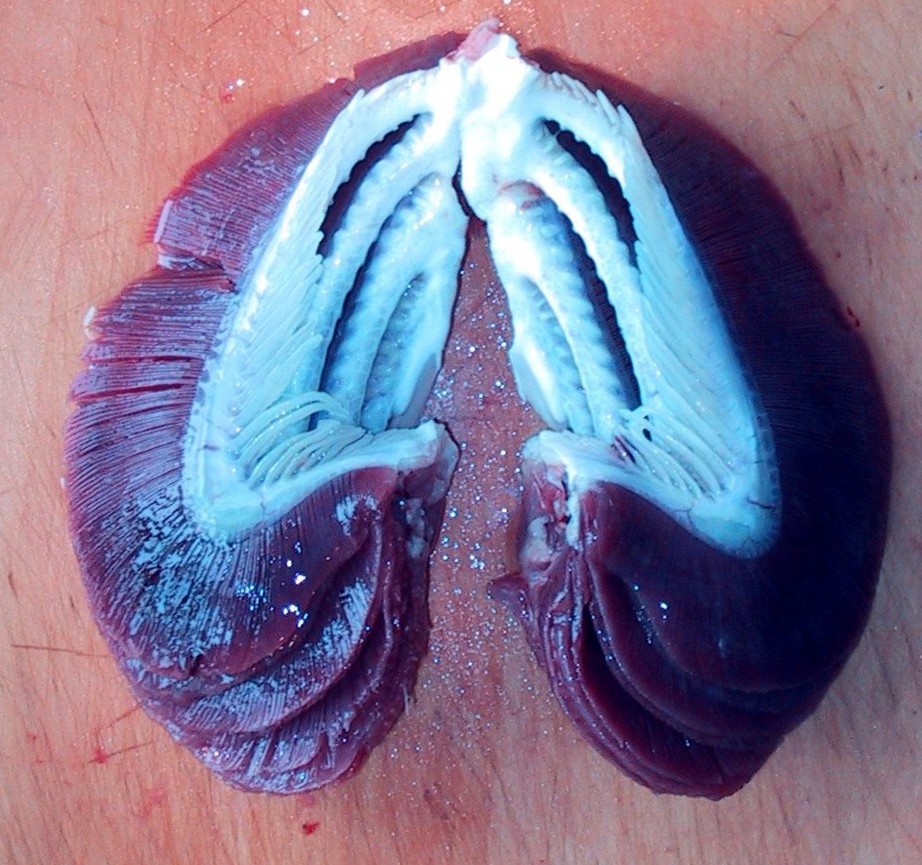
- Branchial skeleton internal to gills. (In contrast to the external branchial basket of lampreys.)
- The mandibular arch - i.e., the jaws, consisting of upper elements - the palatoquadrates and lower elements - Meckel's cartilages.
- The hyoid arch, that helps connect the mandibular arch to the braincase.
- Enlarged forebrain that is flexed ventrally with respect to hind brain. (Compare with ammocoetes larva of lamprey.)
- Paired pelvic appendages
- Horizontal (third) semicircular canal
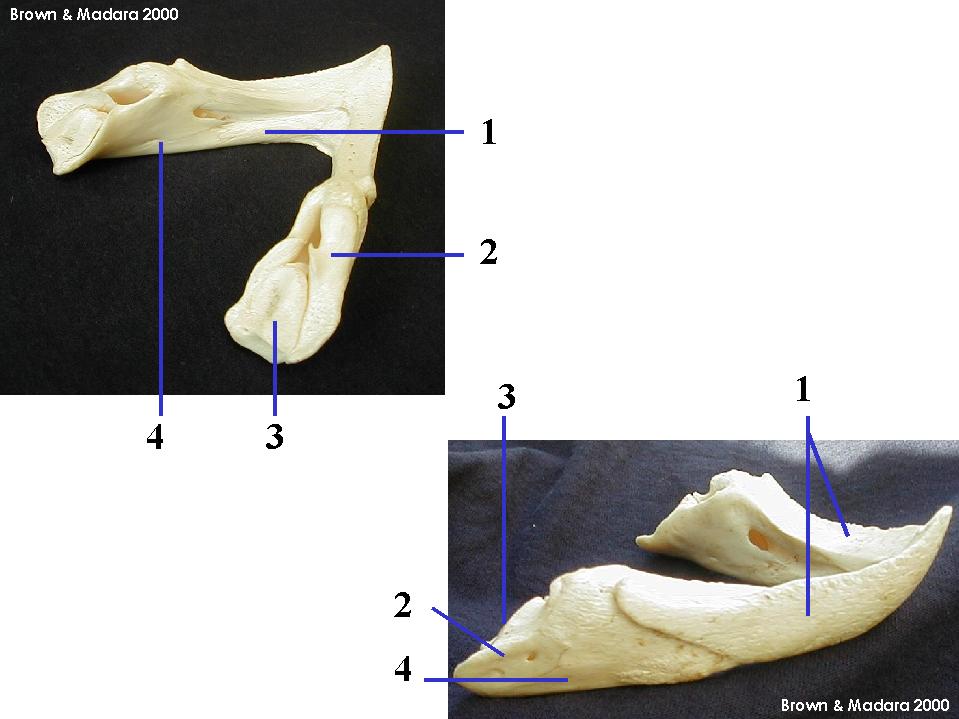
Jaws
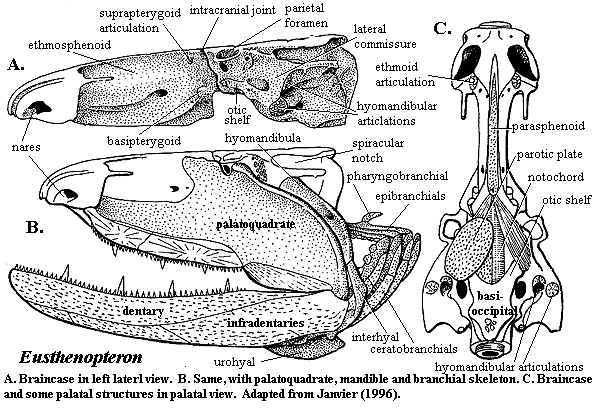
Jaw anatomy: Up until now, when we have spoken of a "skull" we have meant only a plating of dermal bone around the head, or, in the case of Galeaspids and Osteostraci, that plus in endochondrally ossified neurocranium or braincase. With the appearance of the gnathostome jaws and branchial skeleton, the skull becomes a complex composite structure.
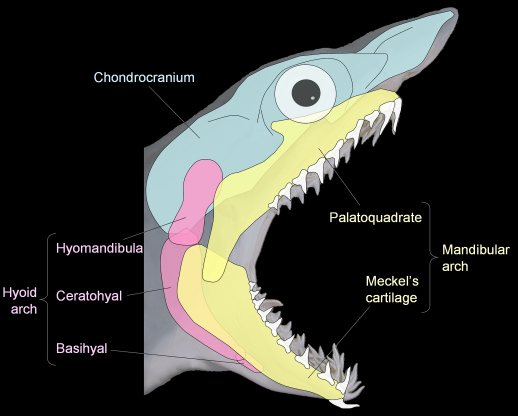
Note: In vertebrates with bony skeletons, these components can ossify in different patterns and into different numbers of elements, but the underlying identity of the cartilage precursors remains the same.
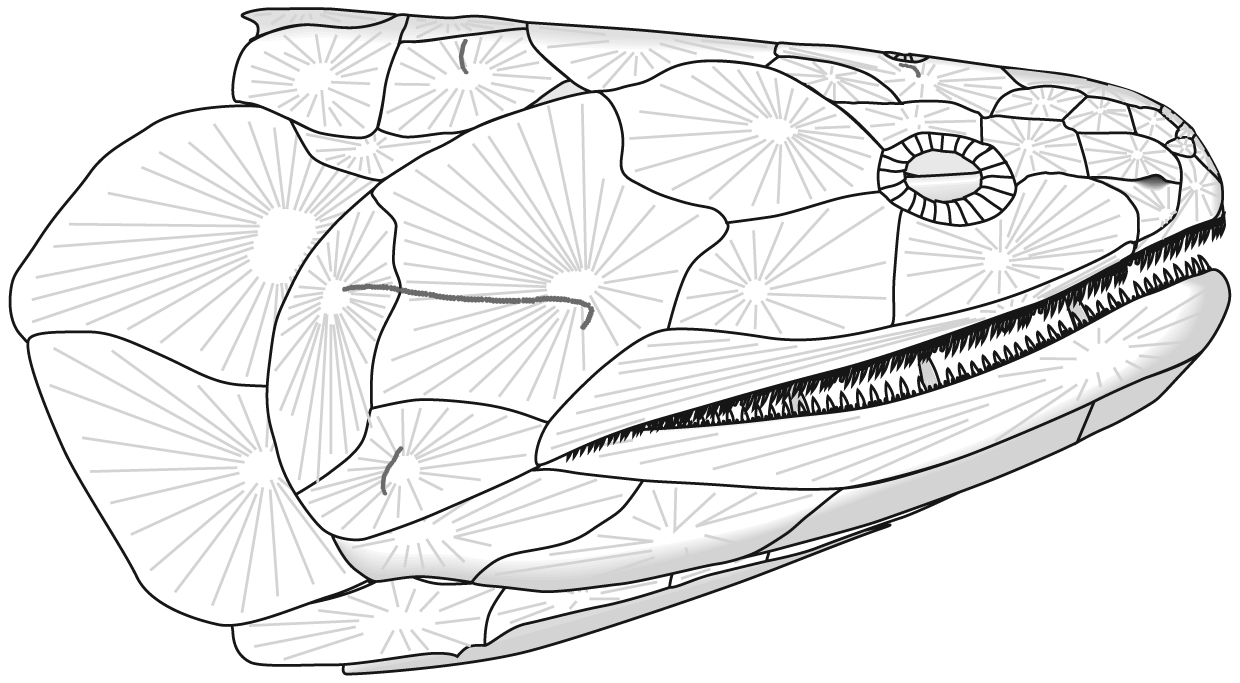
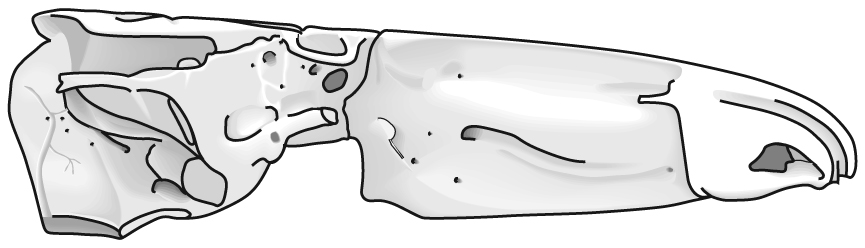
The following illustrations show its components using the fossil bony fish Eusthenopteron as an example.
- Start with the neurocranium:
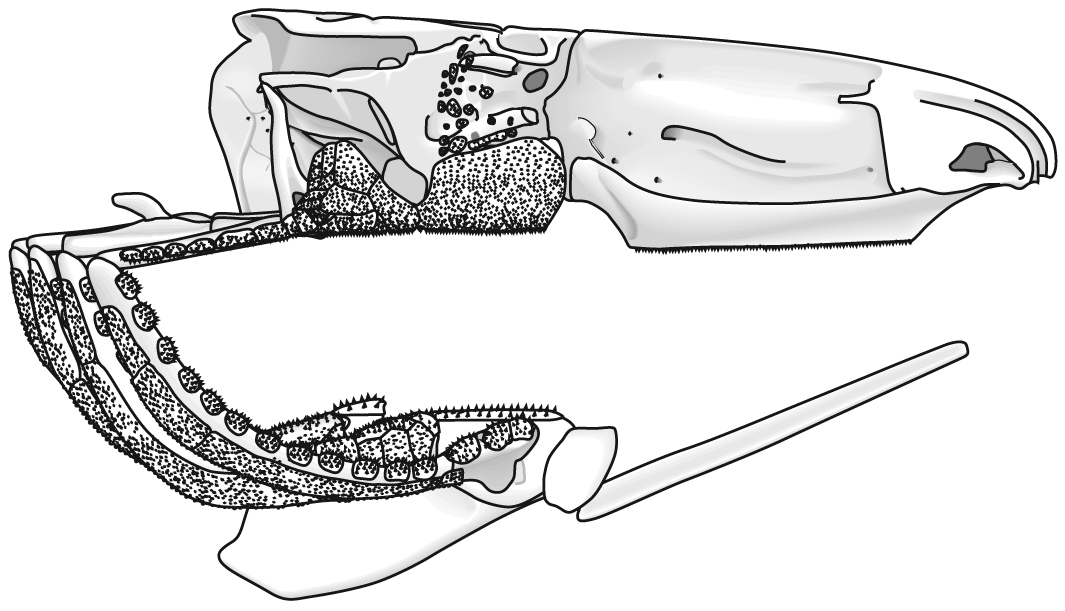
- Add the branchial arches:
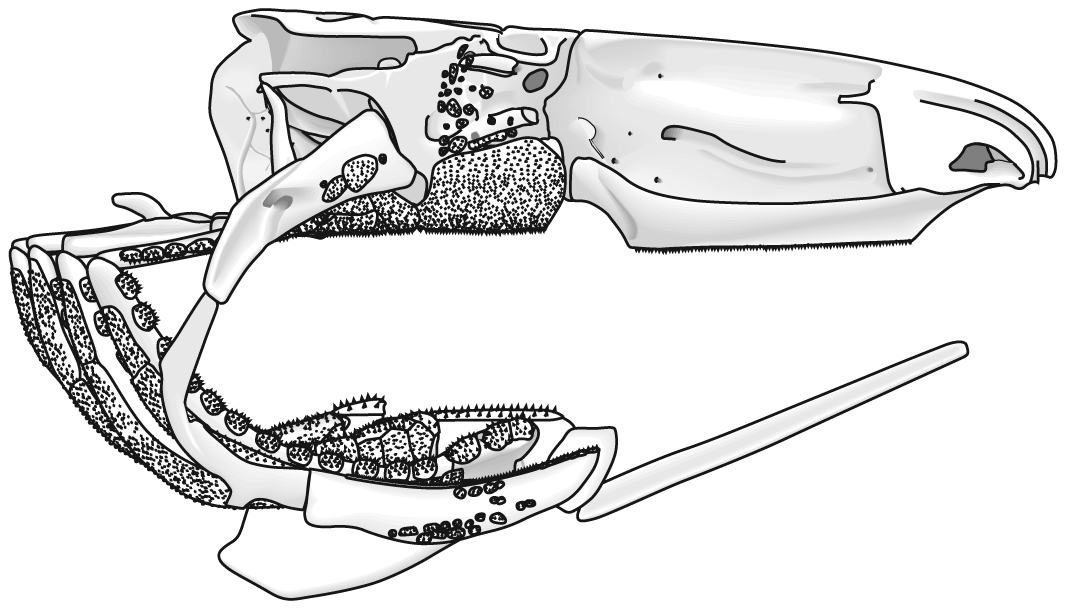
- Add the hyoid arch:
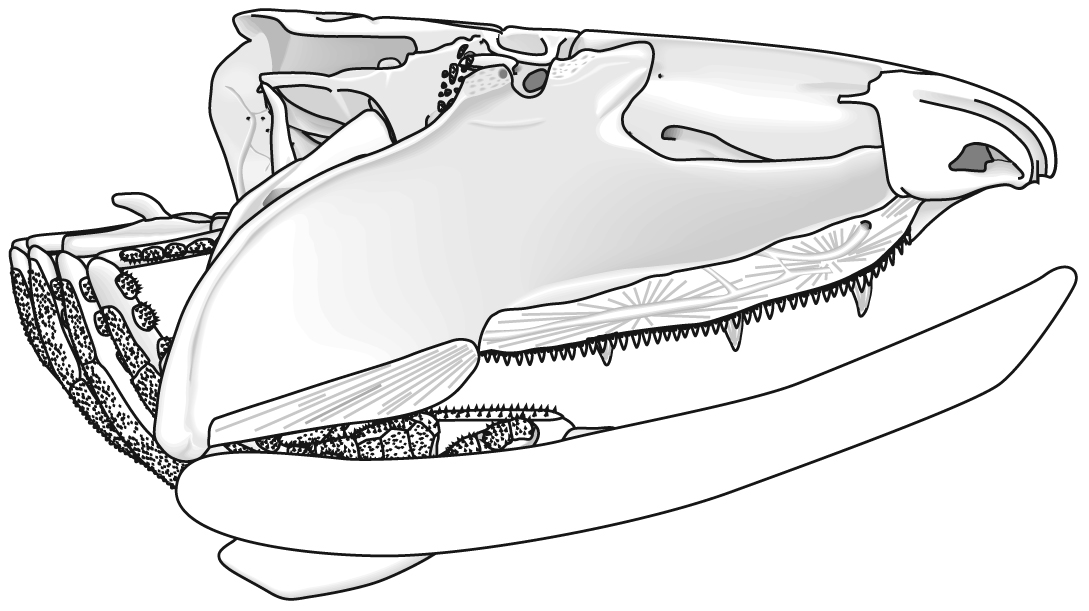
- Add the mandibular arch - palatoquadrate and Meckel's cartilage:
Note, whereas the branchial arches are separated from the hyoid arch and from one another by gill slits, the space between the hyoid and mandibular arches is either closed or perforated by the spiracle. (This is particularly conspicuous in rays, who breath in through it.)
- Finally, add the dermal bone of the skull roof:
Gnathostome diversity:
Traditionally, three major groups of unknown relationships were recognized:
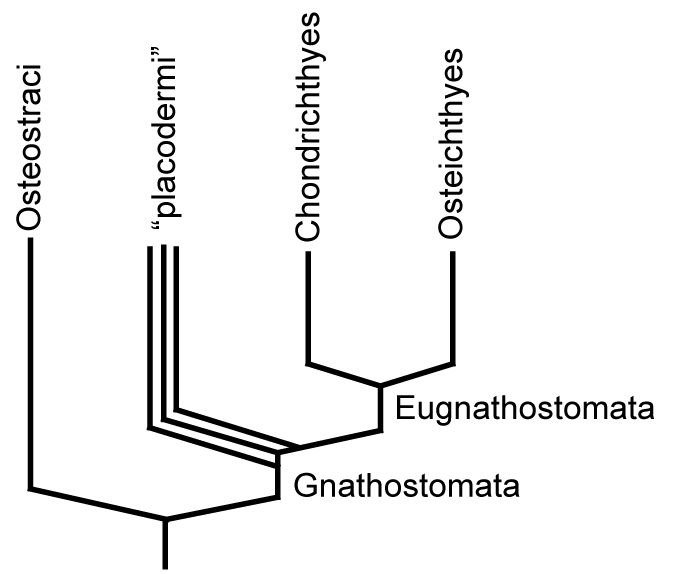
- Placodermi - Paleozoic forms with extensive bony armor
- Chondrichthyes - cartilagenous fish
- Osteichthyes - bony fish
Placodermi: Silurian to Devonian armored gnathostomes experienced a rapid worldwide diversification and sudden decline.

- Identifying features:
- Distinct cranial and thoracic armor with unique ossification pattern.
- Jaws lined with self-sharpening occluding bony plates. (Whether or not these represent fused teeth is open to debate. Some placoderms, at least, had proper teeth.)
- Adductor muscles (that close the jaw) pass medial to (inside) palatoquadrate.
- Distinct cranial and thoracic armor with unique ossification pattern.
- Predation; Some of the largest predatory fish ever , such as Dunkleosteus, belonged to the placoderm group Arthrodira. The name refers to the proper joint at which their head and thoracic shields articulated. Many seem suited to resting on the bottom and engulfing prey by rapidly raising their head and opening jaws. Some localities, however, preserve streamlined pelagic (i.e. open ocean) placoderms.
- Some placoderms were detritus feeders. One group, Antiarchi, had greatly expanded box-shaped thoracic armor and pectoral fins enclosed in arthropod-like armor as well. Serial sectioning of some specimens suggests the possible presence of lungs. Perhaps the weird pectoral fins were used for crawling briefly on land?

Janvier, Phillipe. 1993. Early Vertebrates
 Placoderm surprise:
Recently, Martin Brazeau, 2009 and Zhu et al., 2013. have analyzed basal gnathostome phylogeny and found that placoderms are paraphyletic, with some groups, including the arthrodires being more closely related to living gnathostomes and others, including the antiarchs, being more basal.
Placoderm surprise:
Recently, Martin Brazeau, 2009 and Zhu et al., 2013. have analyzed basal gnathostome phylogeny and found that placoderms are paraphyletic, with some groups, including the arthrodires being more closely related to living gnathostomes and others, including the antiarchs, being more basal.
Eugnathostomata: The gnathostome crown-group. The last common ancestor of Chondrichthyes and osteichthyes and all of its descendants.
Synapomorphies of Eugnathostomata include:
- Distinct anterior and posterior dorsal fins
- Horizontal septum of trunk muscles
Chondrichthyes - Cartilaginous fish (Silurian - Rec.)

Basic definitions:
- For our purposes, Chondrichthyes (cartilaginous fish) is defined as all organisms more closely related to modern sharks and chimaeras than to any other gnathostomes.
- Synapomorphy: It is often said that chondrichthyans are characterized by lack of internal bone. While true, it is not quite diagnostic. In fact, the unambiguous synapomorphy of Chondrichthyes is the presence of prismatic calcification of the cartilage. In contrast with bone, prismatic calcification takes the form of chains of tiny apatite crystals covering the surface of cartilage, likend together by collagen.
Note: Chondrichthyans do not lack other hard tissues. Various groups make teeth, fin spines, and dermal armor out of dentine and enamel. To recap:
Fossil Material: Chondrichthyans are arguably the oldest known gnathostomes, but their fossil record is poor because their bodies lack preservable hard parts that stay articulated when they die. Two notable exceptions:
- Teeth
- Scales - Skin denticles
These structures are similar in histology (microstructure) and differ mostly in size. Teeth are generated in a conveyor belt of gum tissue which moves them into place, then causes them to be shed and replaced by the next member of their tooth family in line. Modern sharks do this very quickly, with individual teeth being functional for a few days or weeks. This ability has evolved slowly and earlier sharks grew and shed teeth at a lower rate.

Synapomorphies:
- Unique pattern of dermal bones of head and pectoral girdle.
- Dermal skull bones with internal descending laminae that interact with endochondral elements.
- Lungs: The presence of lungs - outpouchings of the esophagus used to obtain suplimentary oxygen from "swallowed" air. As we will see, Actinopterygii and Sarcopterygii exploit this synapomorpy in very different ways.
- Operculum: A large plate of dermal bone suspended from the hyoid arch covering and protecting the gill arches. (Ancestrally consists of three bones, the preopercular, opercular and subopercular.)
- Extensive endochondral ossification of the postcranial body, including limb skeleton and vertebral column.
- Actinopterygii (ray-finned fish)
- Sarcopterygii (lobe-finned fish and land vertebrates.)
 Guiyu (Right. Silurian! Zhu, et al, 2009) is our closest well-known approximation of the last common ancestor of Osteichthyes. Remarkably, it is actually a basal sarcopterygian but also the oldest well-known gnathostome!
Guiyu (Right. Silurian! Zhu, et al, 2009) is our closest well-known approximation of the last common ancestor of Osteichthyes. Remarkably, it is actually a basal sarcopterygian but also the oldest well-known gnathostome!
This creature has features seen in both primitive sarcopterygians and actinopterygians, but retains others typical of primitive stem osteichthyans and stem gnathostomes, such as the stout pectoral spine.
Actinopterygii: (Dev - Rec)

The vast majority of "fishlike" bony fish.
- Identification:
- Fin rays articulate with rows of basal and radial elements.
- Loss of the anterior dorsal fin (see the basal actinopterygian Cheirolepis right. This character is reversed many times in actinopterygian evolution.
These have so vast a record from the Devonian on that it's hard to know where to start. Alas, they exceed the scope of the course.
Sarcopterygii: The lobe-finned fish and tetrapods.

- Identification
- Presence of single basal limb element followed by two. Humerus radius ulna, femur tibia fibula.
- A distinct pattern of unambiguously homologous skull roof elements.
- Bipartite cranium: Endocranium ossifies as front and rear parts that meet at a hinged articulation.
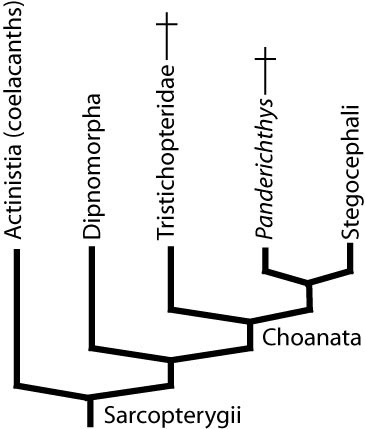
- Actinistia: Coelacanths. (Dev - Rec)
- Dipnomorpha: (Dev - Rec) Lungfish and relatives.

- Their disproportionately small eyes compare well with living fish that inhabit muddy bodies of water (Estuarine or near-shore). In that realm, they were probably powerful predators.
It is often suggested that they were able to move on land. Probably not.

Panderichthys rhombolepis
- Synapomorphies with land vertebrates:
- Complete loss of dorsal fins.
- Frontal bone in dermal skull roof
- Eye sockets located on the dorsal surface of the skull.
- Complete loss of dorsal fins.
- Biological interpretation: Panderichthyes, although still aquatic, was anatomically somewhat specialized for shallow water, possibly stalking prey on shore.

From Daeschler et al., 2006, this creature has become the new poster-child of vertebrate evolution. It is featured in:
- A new popular book
- The Colbert Report
- Southpark (rather vulgar. Search for "retard baby fish.")
In its general profile, it's similar to Panderichthys, there are two important differences.
- The wrist. Although fin rays are still present (and digits absent), its wrist is capable of flexing. Thus, it could support at least some of its weight on its forelimbs. No indication that it ever emerged completely from the water.
- Its skull and pectoral girdle were no longer in contact, allowing the head to be flexed with respect to the torso - the beginnings of the neck.
These earliest stegocephalians (including Panderichthys and Tiktaalik) display an interesting trend in the evolution of the spiracle, which is:
- Open
- Large
- Positioned on the upper surface of the skull
Link to additional info for all of your Tiktaalik needs.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Very well known from relatively complete skeletons.
- Breathing
- Open operculum and functional gills present in adult.
- Notch in rear of skull apparantly accomodated fish-style spiracle, not an impedance-matching ear.
- Open operculum and functional gills present in adult.
- Mechanical support
- Flexible wrist, ankle
- Slightly flexible elbow and knee.
- Fin rays lost from all but the caudal fin
- Fingers (eight per limb) and toes (seven per limb) present.
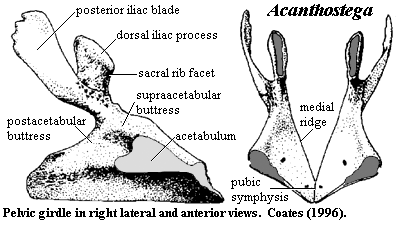
- accessory articulations between neural arches, but no zygapophyses to speak of.
- Pelvis is small but has a ligamentous connection with a sacral rib.
- Feeding:
- First vertebra is enlarged, but otherwise no specialization of the vertebral column, such as zygapophyses (intervertebral articulations) .
- First vertebra is enlarged, but otherwise no specialization of the vertebral column, such as zygapophyses (intervertebral articulations) .
- Senses:
- Extensive lateral line system
- Robust hyomandibula/stapes articulates with fenestra ovalis.
- Extensive lateral line system
- Overall impression is of an aquatic animal that perhaps primarily used hands and feet in walking on bottom or through underwater vegetation.
{kind=link}
{kind=link}
- Martin Brazeau, 2009 The braincase and jaws of a Devonian "acanthodian" and modern gnathostome origins Nature 457, 305-308.
- Edward Daeschler, Neil Shubin, Farish Jenkins, Jr. 2006. A Devonian tetrapod-like fish and the evolution of the tetrapod body plan. Nature 440, 757-763
- Robert Sansom, Kim Freedman, Sarah Gabbott, Richard Aldridge, and Mark Purnell. 2010. Taphonomy and affinity of an enigmatic Silurian Vertebrate Jamoytius kirkwoodi White. Palaeontology 53(6), 1393-1409.
- Min Zhu, Wenjin Zhao, Liantao Jia, Jing Lu, Tuo Qiao, and Qingming Qu, 2009. The oldest articulated osteichthyan reveals mosaic gnathostome characters. Nature 458, 469-474.
- Min Zhu, Xiaobo Yu, Per Erik Ahlberg, Brian Choo, Jing Lu, Tuo Qiao, Qingming Qu, Wenjin Zhao, Liantao Jia, Henning Blom & You'an Zhu, 2013 The braincase and jaws of a Devonian "acanthodian" and modern gnathostome origins. Nature 502, 188-193.
- Edward Daeschler, Neil Shubin, Farish Jenkins, Jr. 2006. A Devonian tetrapod-like fish and the evolution of the tetrapod body plan. Nature 440, 757-763
To Next Lecture.
To Previous Lecture.
To Syllabus.