

Key Points:
•Paleoart--techniques to restore the appearance of fossil organisms--should be rooted in scientific information.
•To recreate the appearance of a fossil dinosaur, several steps are necessary. One needs to fill in the missing parts of skeletons (based on other specimens of the same species or related ones); reconstruct the muscles and other organs supported by that skeleton, and drape the muscled skeleton with skin.
•A particularly problematic part of paleoart concerns what to put on the outside of the skin. We know that all clades of dinosaurs had scales on at least part of the body, and (based on skin impressions of "mummies") many clades were primarily scaled. But other clades (particularly among coelurosaurian dinosaurs, but also within Ornithischia) had fuzzy protofeathers or honest-to-goodness pennaceous feathers. The exact distribution of fuzz (both phylogenetically within Dinosauria and on the body of individual animals) remains poorly resolved.
•Restoring the color--or at least some aspects of color--is possible for some species. Scale and feather color is created by several sources: pigments (including color-bearing melanosomes and various elemental stains) as well as different structures within the scale or feather. Some of these can be recovered from exceptional specimens.
•Pigment-bearing structures are not the only fossil biomolecules to preserve in the rocks. Various other proteins, including collagen (the firm connective tissue widespread in animals) can survive for millions of years in certain circumstances.
•Even below the level of molecules, the isotopic composition of dinosaur bone and teeth contains information about the ecology and physiology of the animals.
Bones and Muscles: the Foundations of Paleoart
People are obviously interested in the life appearance of fossil organisms like Mesozoic dinosaurs. The practice of doing such restorations is called paleoart. Here are some basic observations about paleoart before we go into more detail:
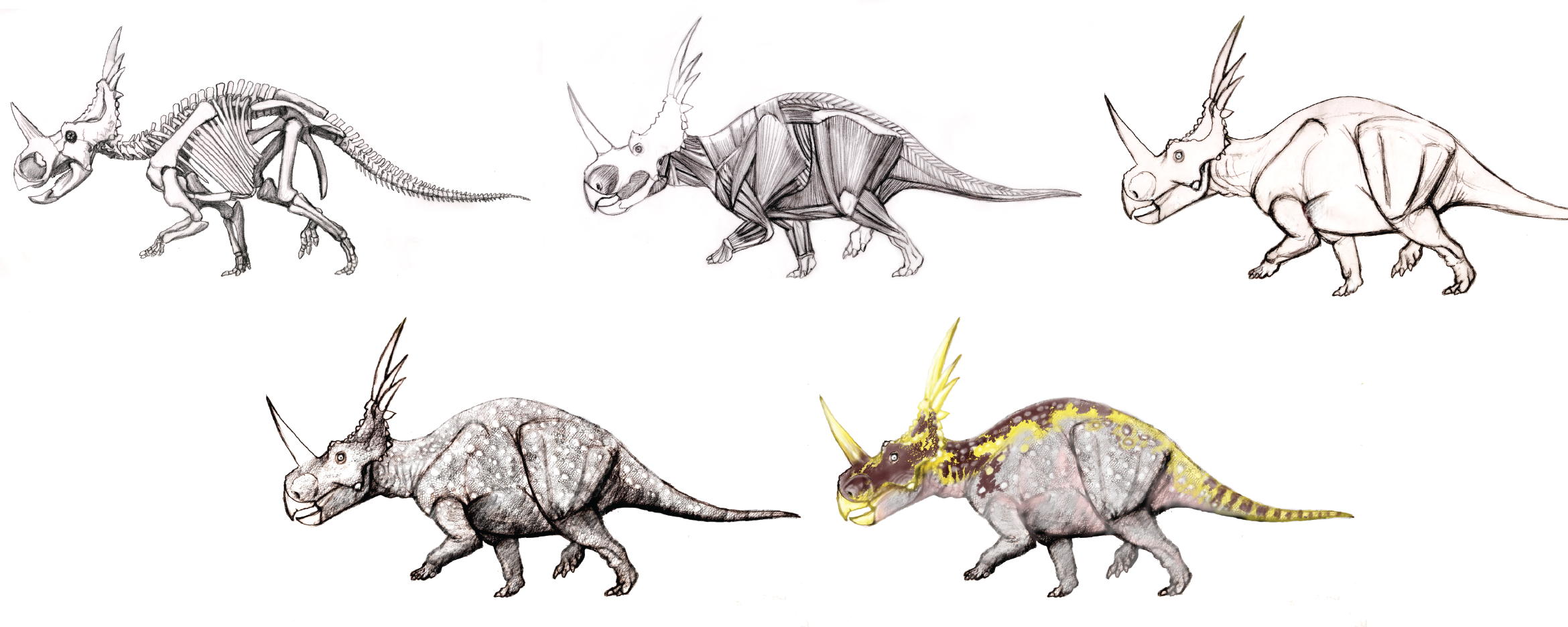
But how do you even begin paleoart? You have to start with the fossils themselves; and for dinosaurs, that means skeletons. With rare exceptions, you won't have a 100% complete skeleton (although sometimes you do!). So you need to complete the skeleton. If there are other individuals of the same species, you can use the non-overlapping bones to create a more complete skeleton. (However, you have to take into account allometric changes of proportion!)
When other individuals of the same species are not available, you can use the bones of close relatives to fill in the gaps. That said, this is less secure in detail (and the further from the species in question you have to go to find the bone, the more likely the ACTUAL bone that is missing would look different.)
The next step is to map the muscles onto the skeleton. Thankfully most of the major muscle groups in amniotes are shared in common, so we are generally pretty secure where the origins and insertions of the muscles fit. In fact, there are often attachment scars and crests on bones to indicate where the muscles fit. Where there are differences among amniotes, we can use phylogeny to look at the muscle groups in birds and crocodylians to see the most likely condition in extinct dinosaurs.
All that said, there is still some uncertainty. The exact size of the muscle in between origin and insertion is not known, and varies of the course of the life of an animal.
Skin Deep: Integument
Once the muscles are mapped out, the restoration can be draped in skin.
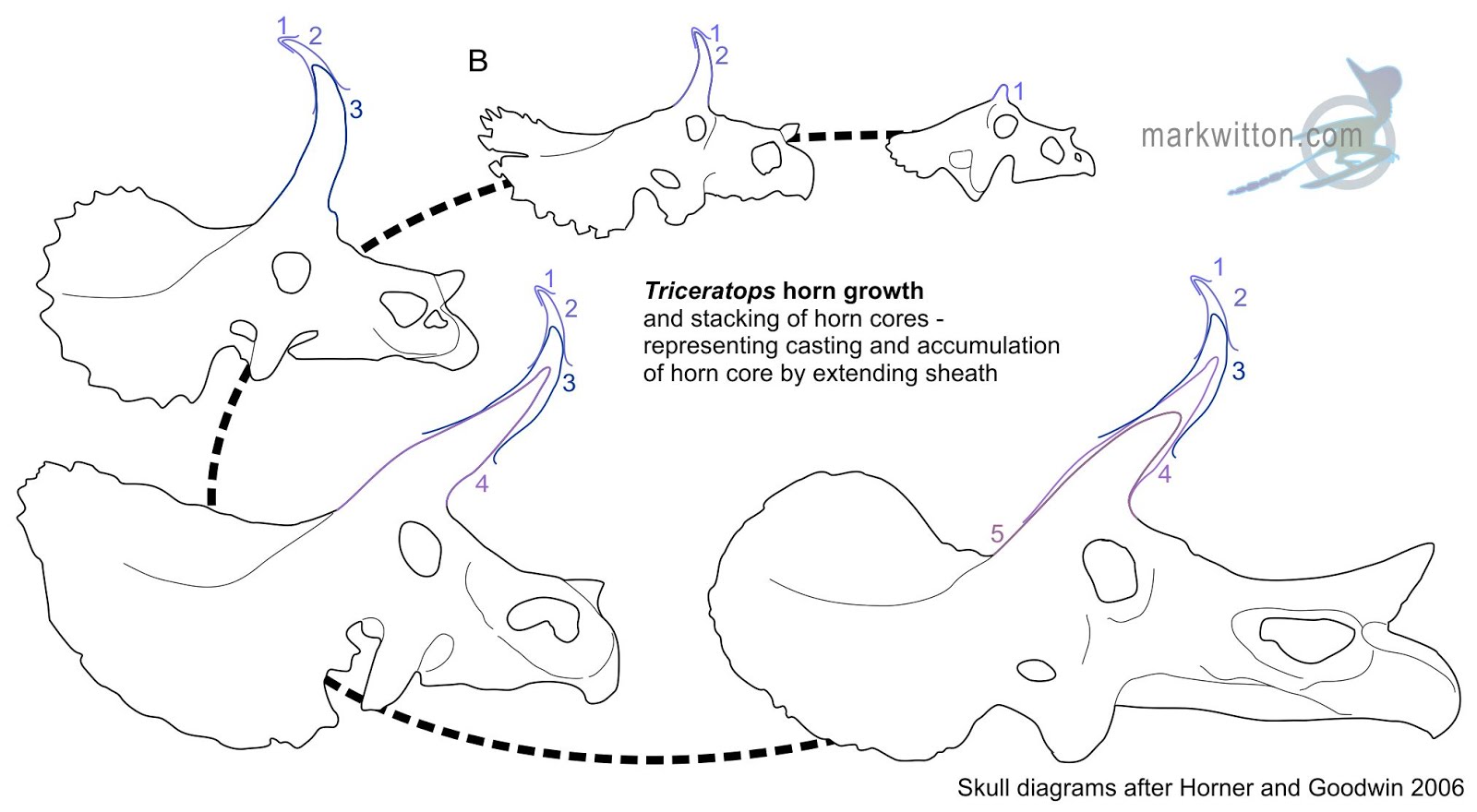
Before we get to the morass of what goes on top of the skin, we'll look at the special case of horny (keratinous) tissue. Various dinosaur species have horns and/or osteoderms. In life these were capped by keratinous sheaths. But how thick were these sheaths? That is, how far beyond the bone did they extend?
A study published on 29 November 2017 examined the osteoderms of the exquisitely-preserved type specimen of the nodosaurid Borealopelta. In this specimen it can be seen how far the sheaths extend beyond the osteoderms. Caleb Brown incorporated this information with other data from living reptiles and mammals with horns in order to see if general patterns exist. It appears that the sheaths grow with positive allometry, making large horns/spikes on larger animals even more exaggerated. Thus the actual horns or spikes of big dinosaurs like Triceratops and Stegosaurus were probably significantly larger than the skeleton alone shows.
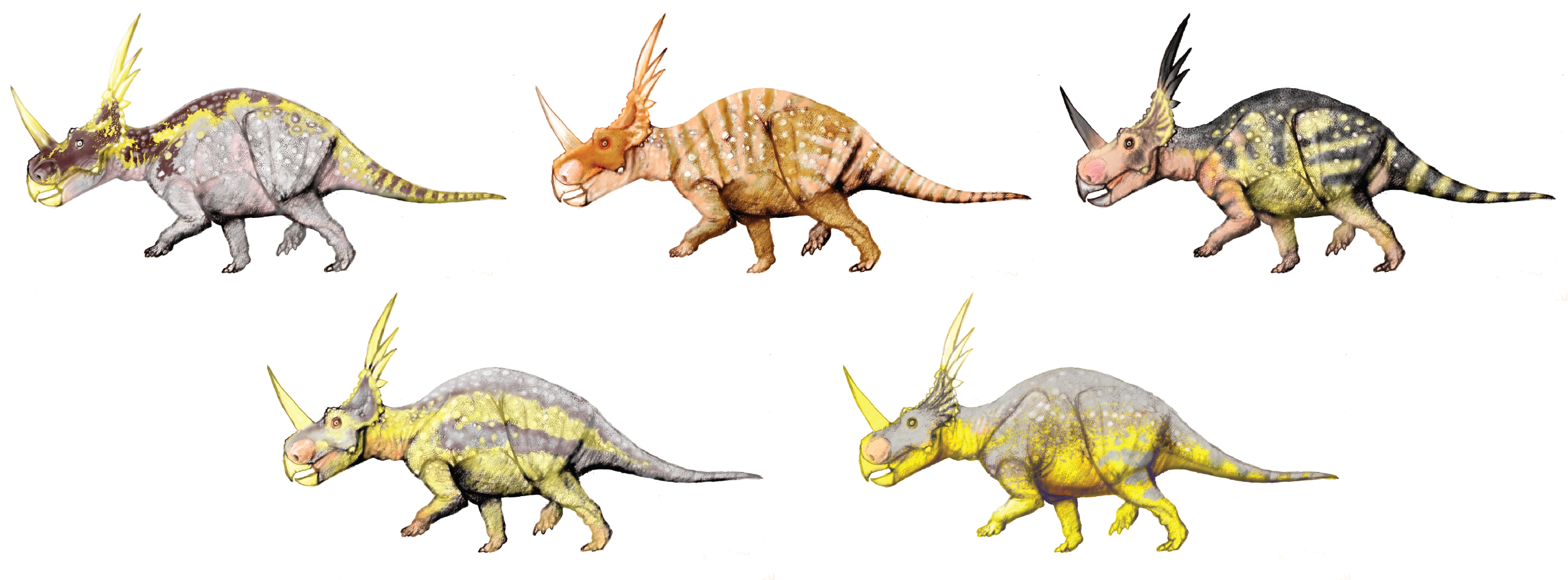
On the outside of the skin is some form of additional integument, which in dinosaurs would be scales or plumulose fuzz or pennaceous feathers. We know that all dinosaurs (including the living ones) have flat scales on at least part of the body (in birds, mostly just on the legs.) We have scaled skin impressions from many groups: hadrosaurids, other ornithopods, ceratopsids, other ceratopsians, primitive neornithischians, sauropods, ceratosaurs, carnosaurs, tyrannosauroids, and various maniraptorans. Those scales tend to be mosaic scales (like on the legs of crocodilians and turtles), rather than the overlapping scales of lizards and snakes.
However, we also have carbonized and/or impressions of other forms of integuments. We have quills in basal ceratopsians; plumulose protofeathers in primitive neornithischians, heterodontosaurids, megalosauroids (if Sciurumimus is a megalosauroid and not a coelurosaur), basal coelurosaurs, tyrannosauroids, and some maniraptoriforms; and pennaceous feathers in ornithomimosaurs, therizinosauroids, and pennaraptorans. If the ornithischian and theropod plumulose structures are homologous, this would mean that these features were present at the base of Dinosauria (if the Saurischia hypothesis is real) or of Ornithoscelida (if ornithischians and theropods are sister taxa).
But the phylogenetic distribution is by no means the same thing as the physical distribution of the features on an individual! There are many factors that might control the actual distribution on an animal:
The net result of this is that we still are not particularly secure on which dinosaur species were fully feathered, which fuzzy, which only scaled, and even on any given individual how much of the body had each. Thus, nearly all dinosaur paleoart has to be speculative with regards to the integument, except for those based on good dinosaur "mummies" (with skin impressions for most of the body) or with good total body coverage of feather impressions.
Other than simple skin covering, sometimes there is evidence of other soft tissue structures. For instance, an impression of the rear of the head of Edmontosaurus regalis shows it had a small crest. But this leads to lots of questions: did most hadrosaurines have such crests? Or just edmontosaurinins? Or just Edmontosaurus? Or was it limited to the species Edmontosaurus regalis? And did all individuals have it? Or just one sex? Or just adults? Was it present all year? Or just during one season?
A brand-new technique may show that more soft tissue records are preserved than meets the eye, at least under visible light. A technique called laser-stimulated fluorescence (where lasers of particular frequency are used to cause faint traces of organic material to light up) has shown that many soft tissue features, including foot pads are actual preserved on specimens of the basal avialian Anchiornis. Future work may show that such features are more commonly preserved than we currently think.
"Unknown Unknowns
But even the most spectacular preserved fossil is not the compete living animal. As any visit to the wild (or zoo, or even nature documentary) will show, animals have all sorts of wonderful, extravagant features that are not obvious from the skeleton. There are lots of anatomical structures and behaviors of fossil species were are unlikely to ever actually see. These are truly "unknown unknowns": we don't even know what it is we are missing (as opposed to a "known unknown", like when he have only the humerus of a skeleton and are secure that there was a forearm, even if we don't know what it looked like.
Some paleoartists have attempted to show how wonderfully weird dinosaurs might have looked, even though we can't possibly know if these cases are accurate. Here are some cases:
This whole paleoart movement is called the "All Yesterdays" movement (after an influential artbook by that name):
Even cartoonists are aware of these issues.
(Of course, one can take this too far...).
Fossilized Color
Color is one of the main features of the appearance of animals that we notice. And up until recently, the color of fossil organisms was considered one of the things we would never know. But in recent years we can begin to assess at least some preserved traces of color.
The first thing to recognize is that scales and feathers create color in various methods rather than just one. Some of the color is generated from actual pigment: chemicals with particular colors themselves. Some of these pigments include carotenoids (often yellowish chemicals generated by plants), porphyrins (modified amino acids expressing a vast spectrum of colors), and melanins (typical black, brown, blue-grey, or reddish brown.) Melanins in particular have been studied in fossils, as they are typically present in animals in melanosomes: subcellular bodies containing the pigment. Melanosomes can be preserved in certain circumstances. And different sized and shaped melanosomes contain different colors. So paleontologist have mapped out the type of melanosomes on some feathered dinosaurs to determine its possible colors and color patterns.
Or at least that is the model. There are paleontologist who disagree that the supposed "melanosomes" are really that, and instead are just bacteria. Others agree they are melanosomes, but do not accept that the 'shape = color' model applies outside of pennaraptorans and mammals. So cautions apply, as always.
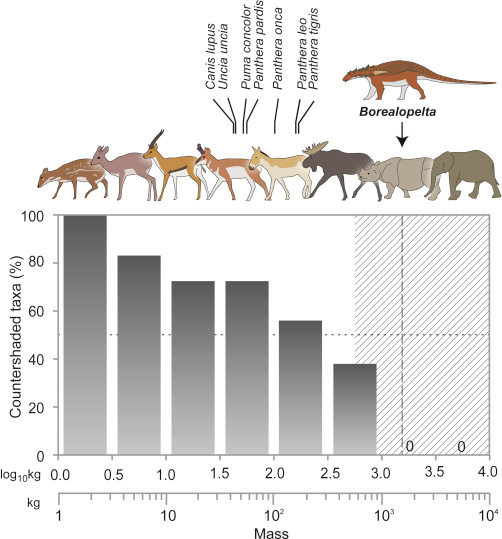
In the exceptionally well-preserved specimen of the nodosaurid Borealopelta there are no preserved melanosomes, but some traces of color have been shown chemically to likely represent the reddish-brown melanin. This has led some to say that it had a brown top. Lack of traces of any melanin on the ventral surface led them to suggest it was whitish underneath, giving the whole dinosaur a counter shade color. Countershading is a common camouflage pattern in modern prey. Curiously, Borealopelta is MUCH bigger than the largest countershaded modern herbivores, but living in a world with predators much larger and (especially) taller than are found today, this isn't that much of a surprise.
However, caution must again be asserted. The absence of preserved melanin on the ventral surface is not in fact evidence of the absence of melanin there! Corpses are never perfectly pristine, and this particular individual sank belly-upwards in the Cretaceous shallow inland seas: its ventral skin may have been scavenged prior to burial. The countershaded model is interesting, and quite plausible, but there are alternative hypotheses for the observed pattern.
Other than pigments, another main means color is achieved is by the structure of the integument. Various factors (including air bubbles in the feather) can render an otherwise brown feather blue. A particular case of structural color is iridescence, the faint (or sometimes not-so-faint) color highlights on a background of a different color. The purples and greens you can see on some starlings and crows, or the flashes of color on the necks of some male hummingbirds, is a byproduct of iridescence. (In some cases, the structure of iridescence is achieved by the use of very long black-pigmented melanosomes interacting with light in a different fashion than the keratin in which they are embedded. The feathers of the small dromaeosaurid Microraptor show this pattern, and so it was likely a black iridescent animal like a starling or crow!
Keep in mind that the ACTUAL color of an animal is a product of the combination of all these going on. For instance, melanins, porphyrins, and/or carotenoids all combine to form various additional colors. And pigments can combine with structures to make additional more (and to create iridescence, as mentioned). So even if we can figure out the presence of one of these aspects for a fossil, it is unlikely that this was ALL of the color that the animal had.
Molecular Paleontology
Beyond melanosomes and other pigments, there are other biomolecules that preserve in the fossil record. Collagen is one such. It is the main connective tissue in animals, and is found throughout dinosaur bone. Although it was thought for a long time that fossils had lost all of their original organic material, in the 1990s it was found that collagen can hang around for hundreds of millions of years if the taphonomic conditions are right.
There have even been supposed dried remains of dinosaurian red blood cells found in fossils. However, once again there are some who suggest these are actually decayed remains of bacteria rather than red blood cells.
Isotopic Paleontology
Below the level of whole molecules, there are actually individual atoms that contain important information about fossil organisms. This is because atoms of a given element might exist in different isotopes. Isotopes are variations of atoms with the same number of protons (and electrons), but differing number of neutrons. As a result of the different weight, different isotopes will be sorted out by various physical and chemical processes (what is called fractionation). Because the isotopes we are concerned with here (that is, non-radioactive isotopes) are stable, they can be preserved in the fossil record.
For one instance, we can use isotopes to model the migration of extinct species. The isotopic composition of water varies from spot to spot due to temperature, evaporation, and other factors. In a study geochemist Henry Fricke and colleagues compared the isotopic composition of clay minerals and the teeth of the macronarian sauropod Camarasaurus from several sites in the Morrison Formation (Late Jurassic, western North America). While the clays only had the isotopic composition of the lowland environment (not surprisingly, since that's where they formed and that's where the teeth were found), but the Camarasaurus teeth show a broader spread of values, including upland signatures. Looking within individual teeth, it was found that some succession of layers show lowland values and then switch to show upland values. This has been interpreted as indicating migration between uplands and lowlands within the species.
Nitrogen and carbon isotopes also get fractionated up a trophic pyramid. Some work has been used to determine (for instance) feeding patterns in fossil communities, including recognition that spinosaurids fed more from aquatic food than land food.
Additionally, oxygen isotopes can be used to estimate the temperature at which a bone crystal formed. Some studies have suggested that dinosaur body temperature was independent of the ambient environmental temperature, which would point to homeothermy (although whether from gigantothermy or metabolic regulation would not be discernible from this sort of analysis).
To Next Lecture.
To Previous Lecture.
To Lecture Notes.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
/https://public-media.smithsonianmag.com/filer/20110520083205Anchiornis-colors.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}