Gnathostomata and Placodermi
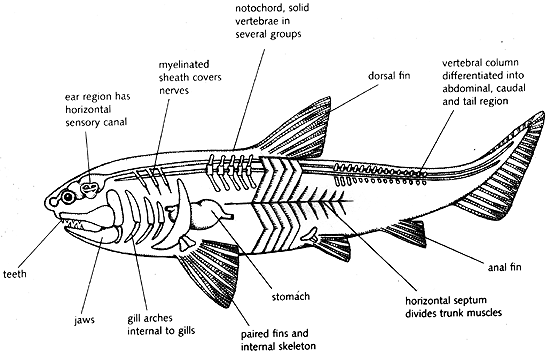
Radinsky, Leonard, 1987 Evolution of Vertebrate Design
Chronology:
-
The earliest well-preserved gnathostomes are a placoderm and an chondrichthyan from the Early Silurian Huixingshao Formation of Chonqing, China. Prior to this, the record consists of individual placoid scales dating as far back as the Late Ordovician that are boldly assigned to Chondrichthyes. (Zhu et al., 2022)
Synapomorphies:
-
Developmental:
- Four Hox gene clusters (compared with two ancestrally in Vertebrata and one in invertebrates.)
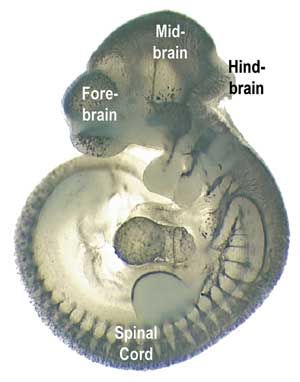
- Enlarged forebrain that is flexed ventrally with respect to hind brain. (Compare with ammocoetes larva of lamprey.)
- Paired nasal capsules communicate with the outside through paired nostrils. Their bony openings are the nares (sing. "naris")
- Hypophyseal duct opens onto roof of mouth
- Trabeculae present as initial chondrification of sphenoid region of braincase. (See braincase development schematic from Palaeos) (Kuratani and Ahlberg, 2018)
- Postorbital process of neurocranium (right)
- Horizontal (third) semicircular canal
- braincase incorporates occipital arch posteriorly.
- Pectoral fin supported by a pectoral girdle with:
- Paired endochondral scapulocoracoids
- Various dermal elements.
- Paired pelvic appendages with internal skeleton and endochondral pelvic girdle. (Only the most basal gnathostomes are slightly odd with dermal pelvic elements and possibly lacking endochondral ones (Zhu et al., in press).)
- Branchial skeleton internal to gills. (In contrast to the external branchial basket of lampreys.) Although it was long believed that the gill filaments were ectodermal in gnathostomes and endodermal in lampreys, Gillis and Tidswell, 2017 have demonstrated that they are endodermal so we are confident that they are homologous in all vertebrates.
- The visceral skeleton (gill arches and related structures) consists of jointed elements:
- epibranchials - dorsal
- ceratobranchial - ventral
- Pharyngobranchials dorsal to the epibranchials
- Paired hypobranchials ventral to the ceratobranchials
- Mid-line basibranchials
- Ancestrally there are five or fewer branchial arches.
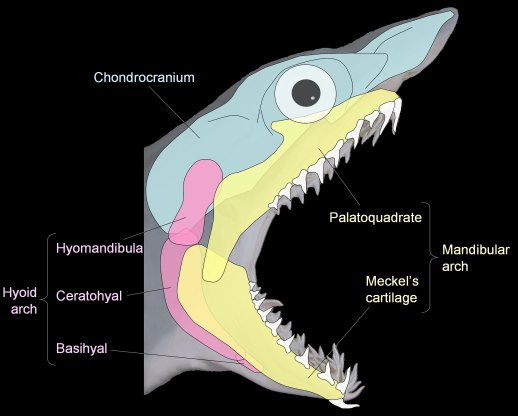
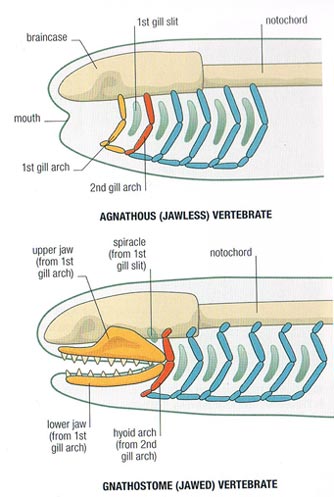
- The jaws made of the mandibular arch consisting of paired:
- palatoquadrates - upper elements
- Meckel's cartilages - lower elements.
- The hyoid arch, that helps connect the mandibular arch to the braincase. It breaks down into:
- Hyomandibulae (sing. hyomandibula) - upper elements that articulate with the braincase
- Ceratohyals - lower elements that support Meckel's cartilages
- Basihyal - ventral mid-line element.
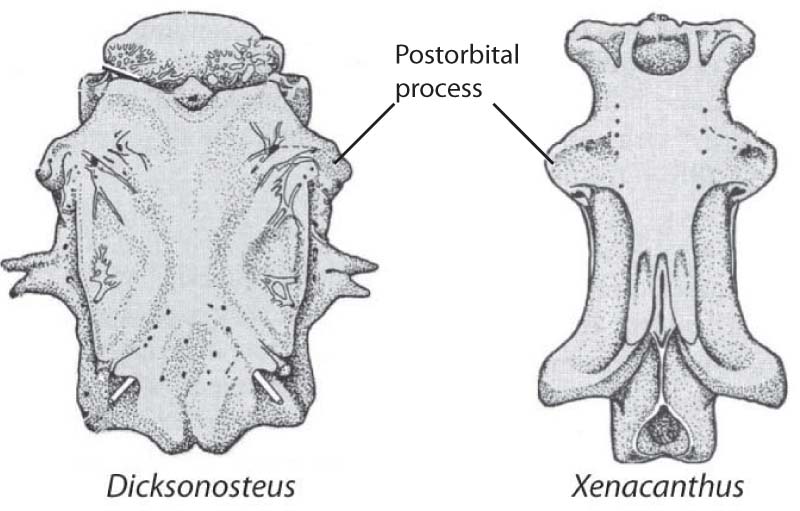
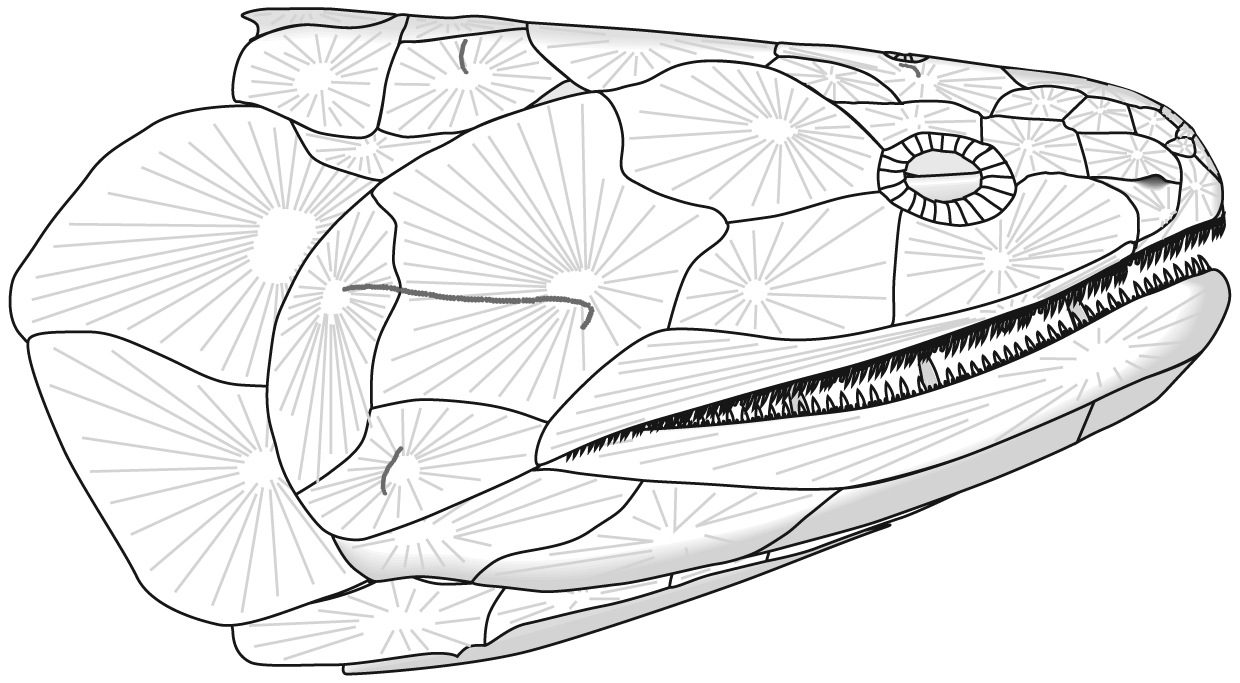
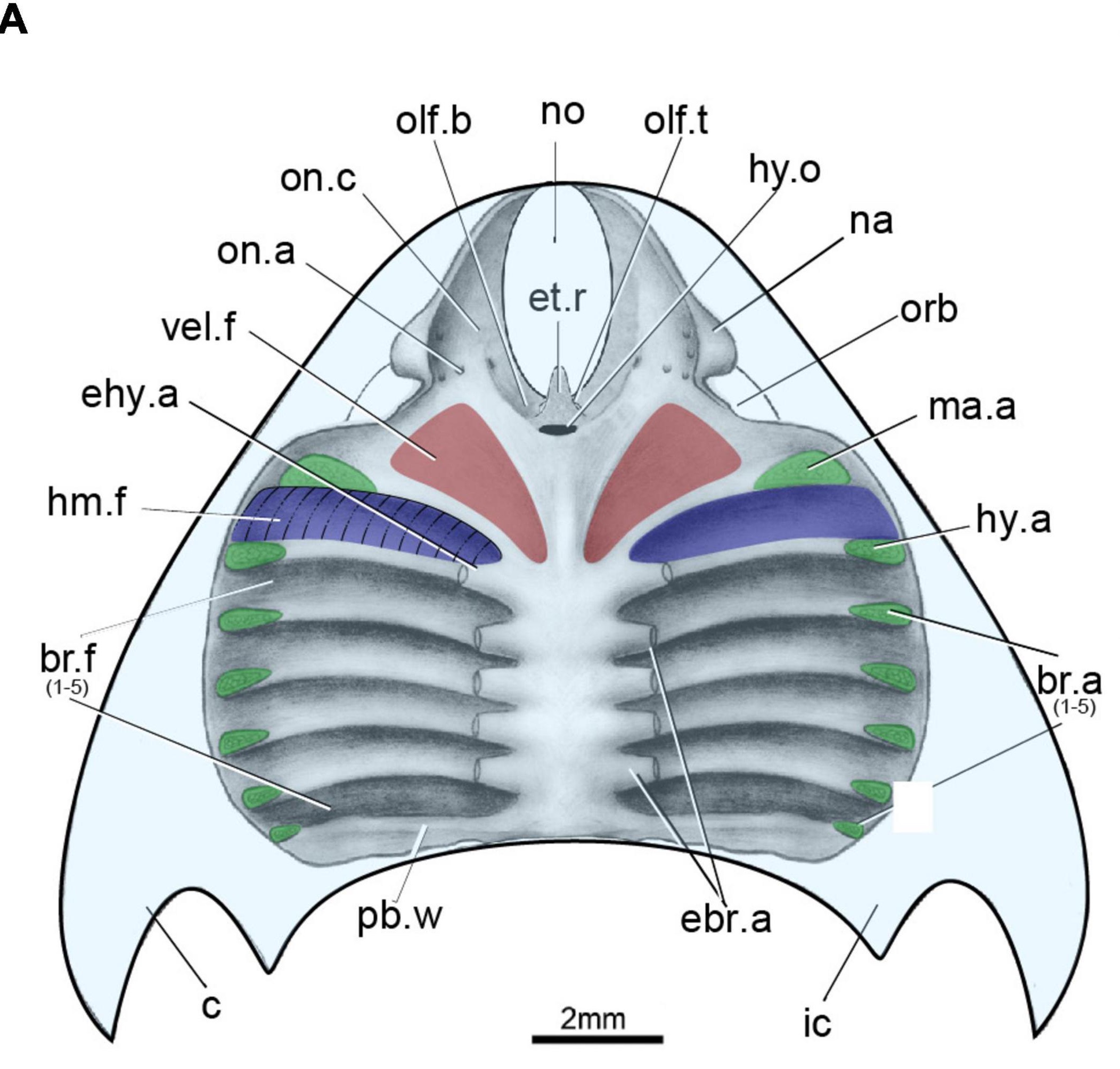
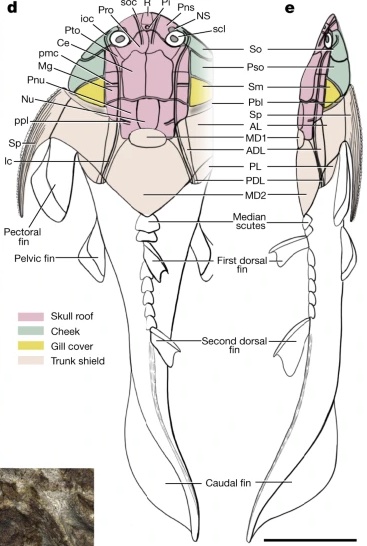
Placoderm (left) and chondrichthyan (right) neurocrania in dorsal view.
Girdles:
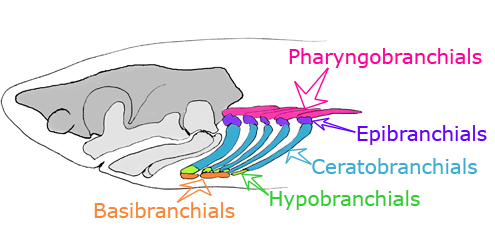
Branchial arches:
The Big News: The first two arches are specialized
This list invokes some challenging problems in gnathostome evolution. These are take up below, after an anatomy review.
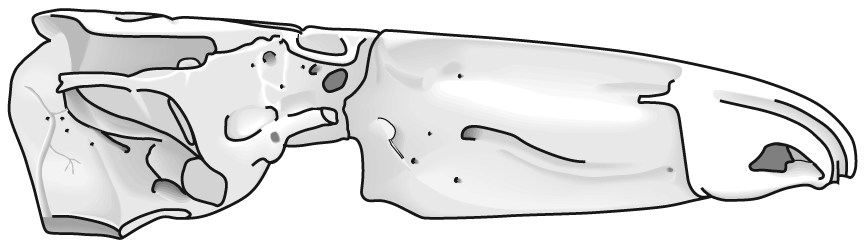
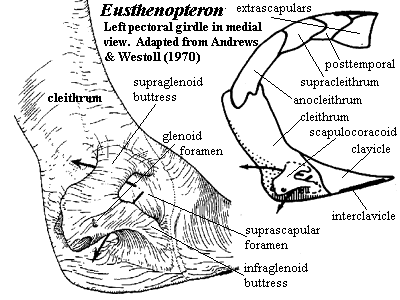
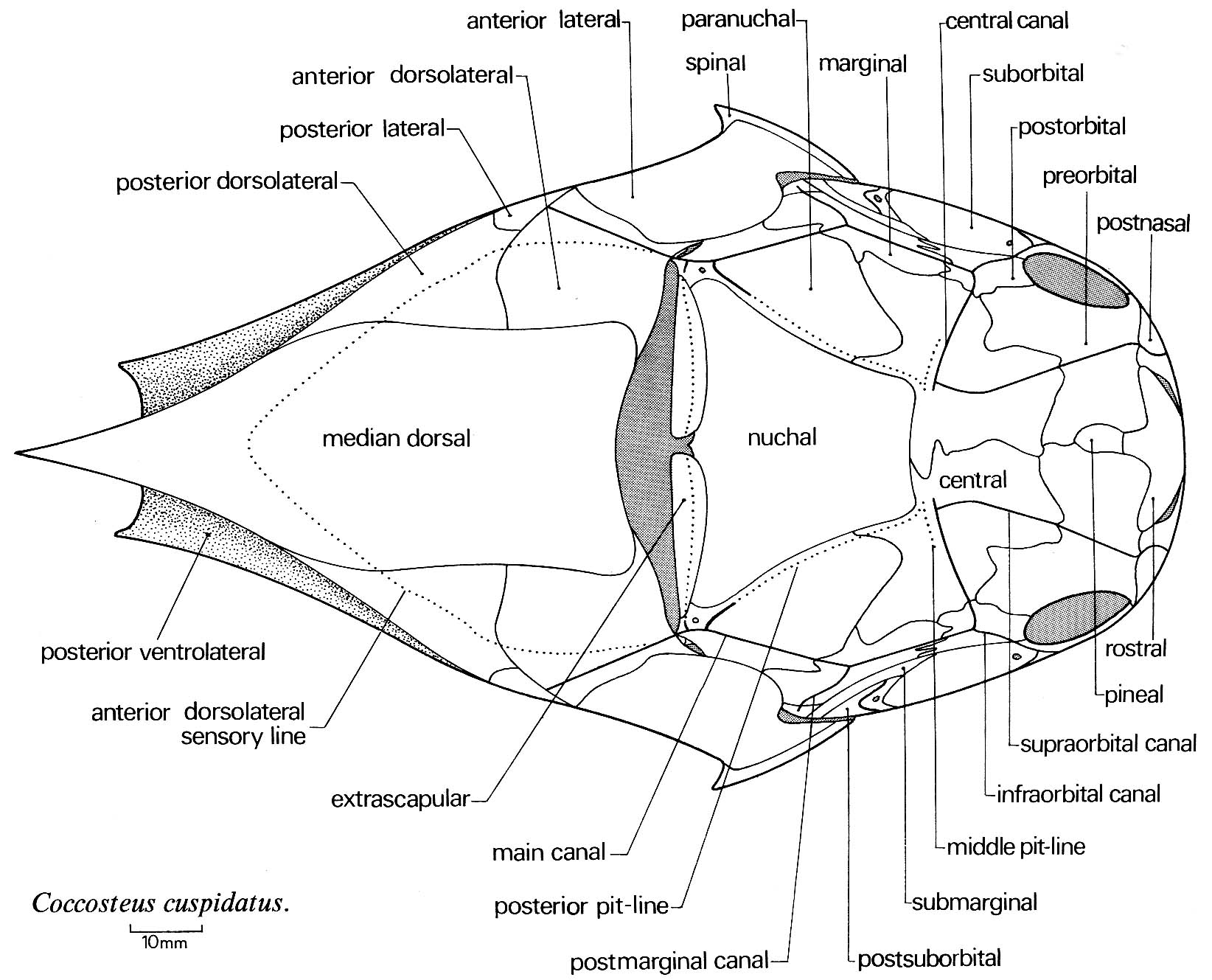
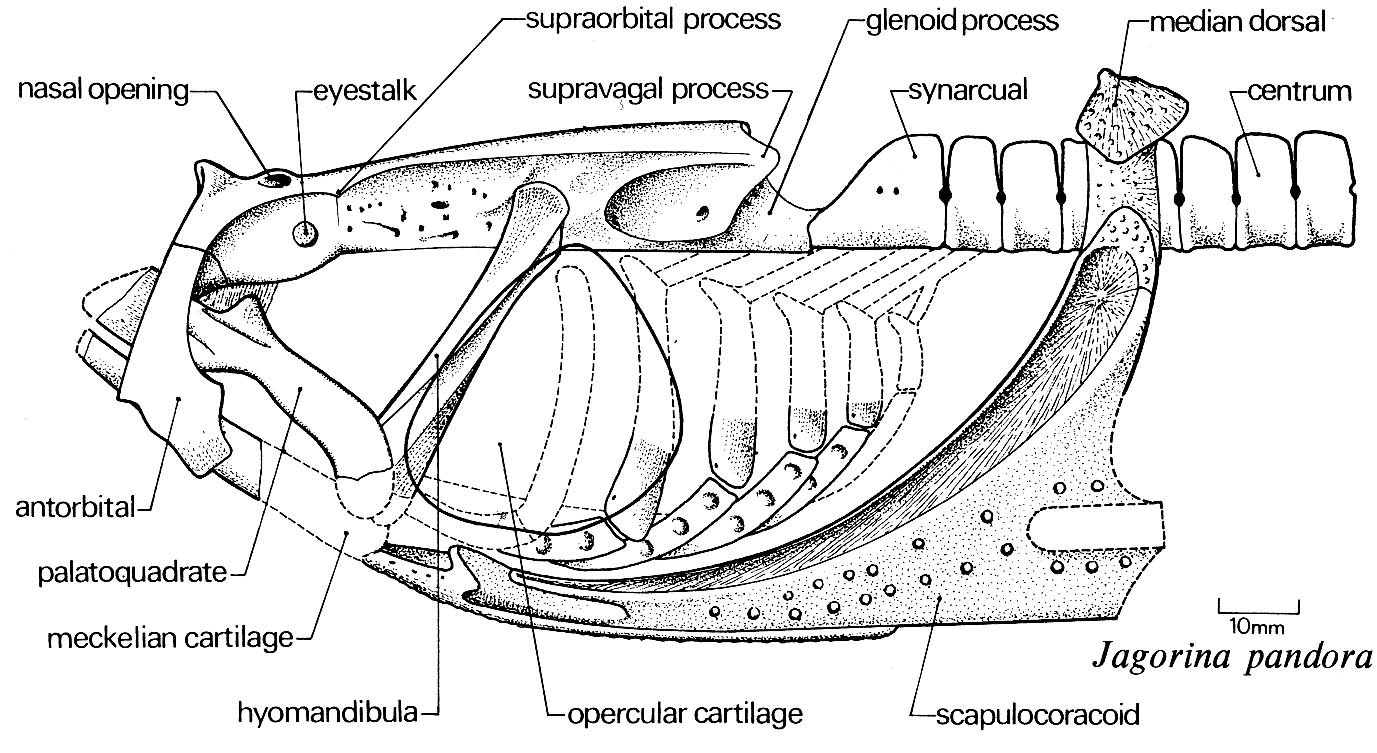
Jaw anatomy: Up until now, when we have spoken of a "skull" we have meant only a plating of dermal bone around the head, or, in the case of Galeaspids and Osteostraci, that plus in perichondrally ossified neurocranium or braincase. With the appearance of the gnathostome jaws and branchial skeleton, the skull becomes a complex composite structure. The following illustrations show its components using the fossil bony fish Eusthenopteron as an example.
- Start with the neurocranium:
- Add the branchial arches:
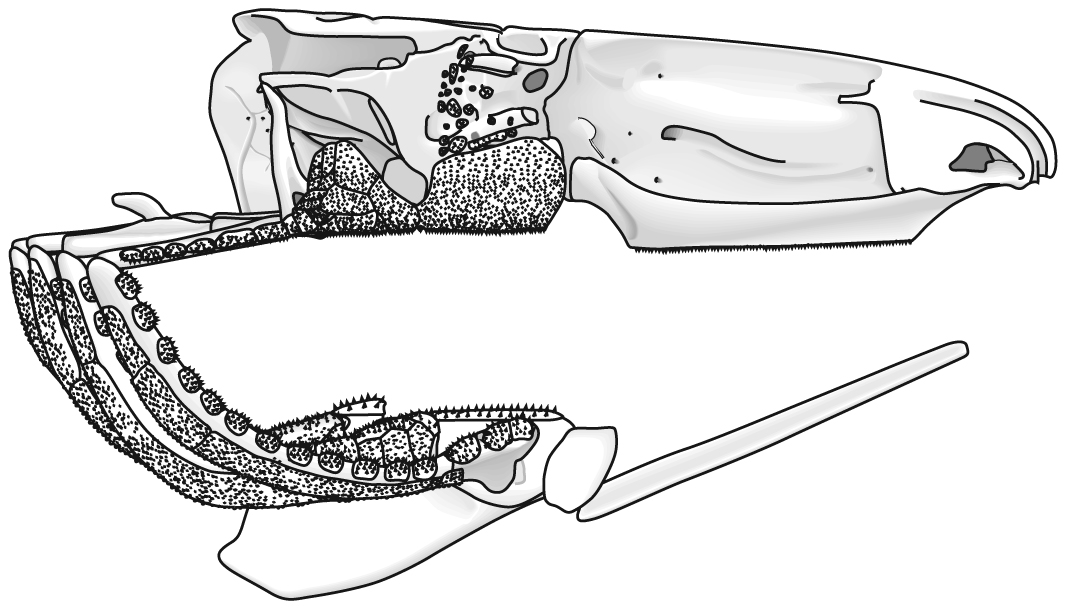
- Add the hyoid arch:
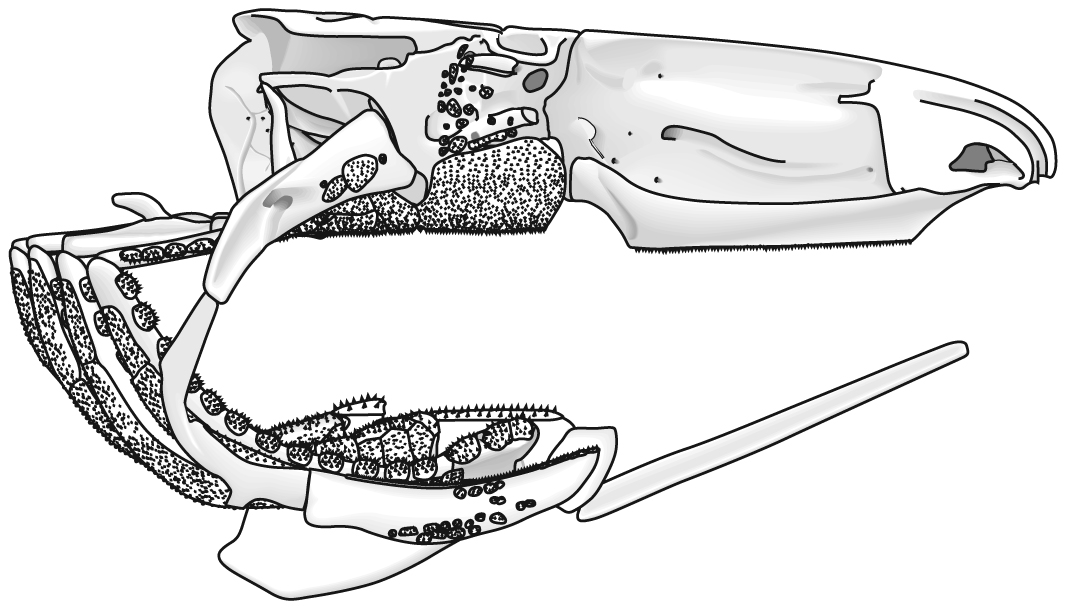
- Add the mandibular arch - palatoquadrate and Meckel's cartilage:
Note, whereas the branchial arches are separated from the hyoid arch and from one another by gill slits, the space between the hyoid and mandibular arches is either closed or perforated by the spiracle. (This is particularly conspicuous in rays, who breath in through it.) See also this link.
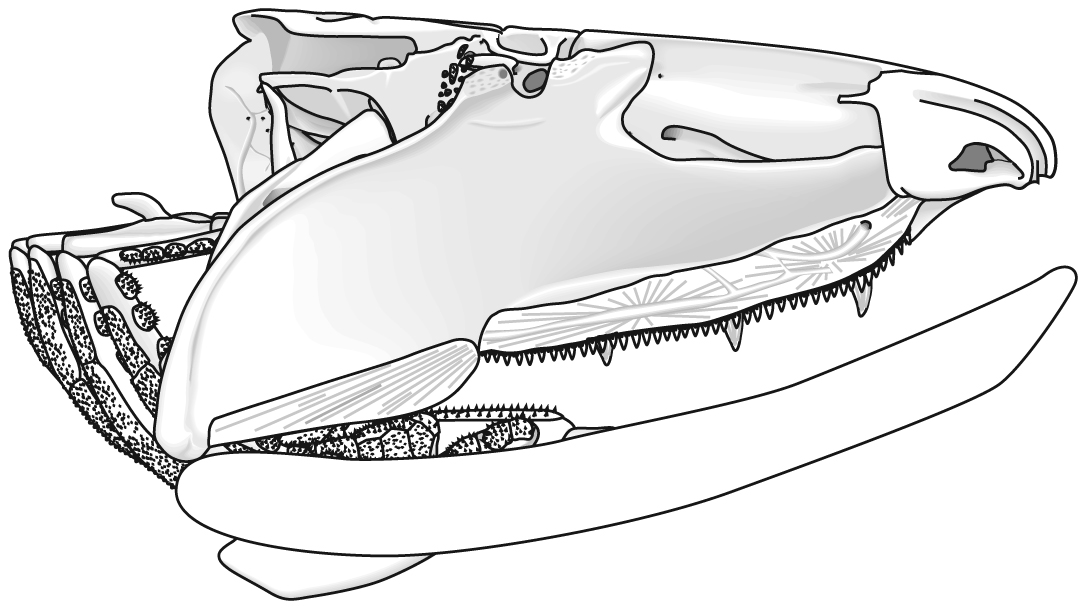
- Finally, add the dermal bone of the skull roof:
Six Gnathostome Issues:
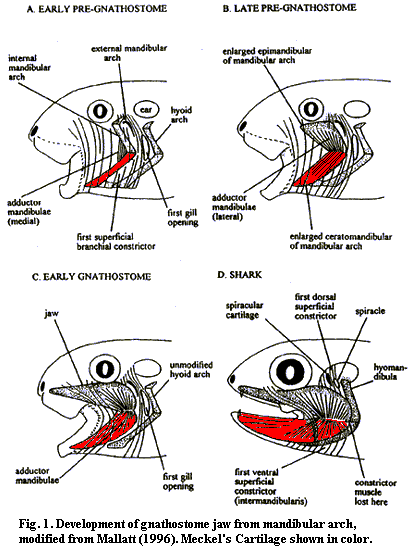
1. The Origin of Jaws: Many gnathostome synapomorphies are continuations of longstanding vertebrate trends toward increased skeletal ossification, brain enlargement, and improvement of swimming. Big exceptions are the changes to visceral arches.
Oddly, fossil intermediaries between jawed and jawless vertebrates just don't seem to exist. What's up?
Three historic hypotheses:
- In many modern gnathostomes, the mandibular and hyoid arches enclose an opening called the spiracle (=eustacean tube in land verts). In living aquatic vertebrates, the spiracle can be invested with pleated gill-like tissue called the pseudobranch. To Gegenbauer, this was the vestigial remnant of what was originally a full gill occupying a full gill slit between the mandibular and hyoid arches, prior to their becoming specialized. Gegenbaur predicted that fossil gnathostomes would be found with a complete gill slits where the spiracle sits.
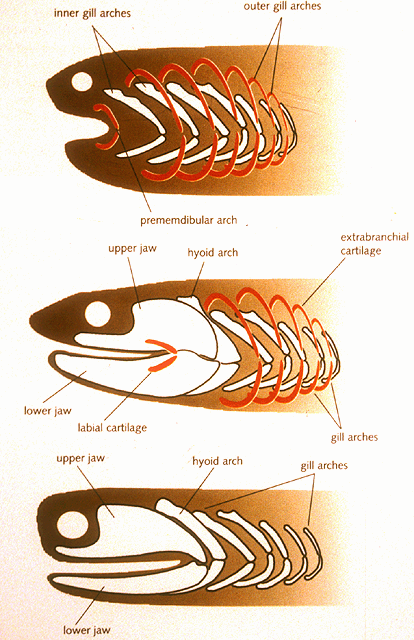
Serial homology of visceral arches: Carl Gegenbaur in 1872 proposed that the jaws and hyoid arch represented modified anterior branchial arches. The earliest versions would have come complete with respiratory gills. Gegenbaur responsibly suggested future fossil evidence that would support this in a specific way:
Alas, but after a century of careful scrutiny, no such transitional forms have been found. Ironically, Gegenbaur's appealing hypothesis was so widely accepted that in many modern textbooks it is still treated as revealed wisdom.
During the 20th century, two new hypotheses have arisen:

Janvier, Philippe. 1993. Early Vertebrates
Indeed, recent developmental studies show that anterior and posterior surfaces of the lamprey velum and anterior and posterior regions of the mandibular arch receive homologous neural crest cells from different regions of the neural tube, and show similar gene expression. (Yokoyama et al., 2020) Establishing this homology allows us to identify homologs of the hyoid arch in lamprey embryos by position and by innervation by the facial nerve, as in gnathostomes.
The 2023 Synthesis: Everyone is right! Finally, Gai et al., 2022 use Synchrotron X-ray Tomographic Microscopy to identify corresponding structures in the galeaspid Shuyu. In Shuyu, however, the hyoid arch and pouch resemble the branchial arches behind them. Patterns of blood vessels around it indicate that it contained functional gills, as well. So everyone is right:
- Fossil fish do occur with full gill slits between the mandibular and hyoid arches, just not jawed fish! Moreover, the mandibular arch never looked like a branchial arch.
- The original of the mandibular arch was at the rear of the original oral cavity
- The velar cartilage was the evolutionary basis for the mandibular arch
- Gai et al., 2022 point out that although no gnathostome has a fully open spiracular gill-slit the acanthodians Cheiracanthus and Mesacanthus appear to have retained both anterior and posterior pseudobranchs in their spiracles. Thus, the evolution of the gnathostome spiracle proceeded in two steps:
- Reduction of the full gill opening with functioning gills in front and in back to a narrow spiracular passage with paired pseudobranchs
- Elimination of one of the pseudobranchs.
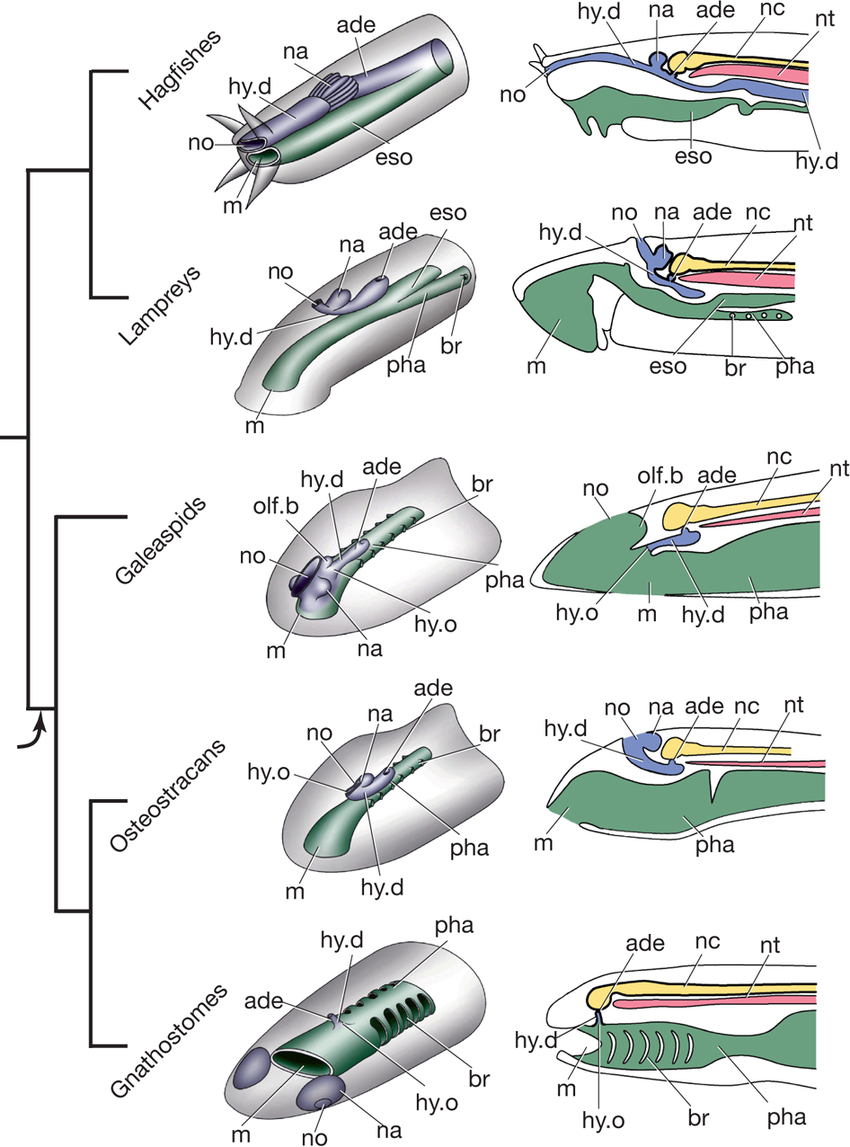
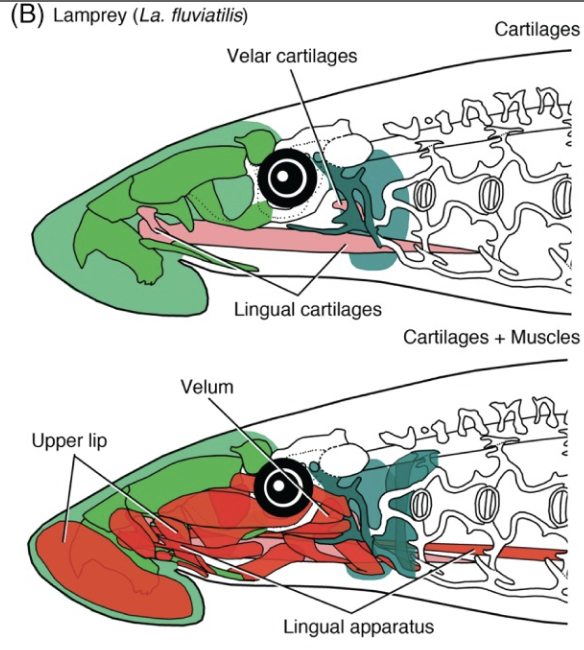
Schematics of mouth, nasal capsule, hypophyseal duct, and pharynx
across jawless taxa. Note: Region between mouth margin and opening
of hypophyseal duct is the cyclostome "upper lip."
But careful! As Miyashita 2015 points out, the cyclostome velum isn't the only neural crest derived structure that expresses genetic markers and innervation homologous to the mandibular arch of gnathostomes. Other homologous structures form many of the cartilages of the front of the face that support the "upper lip," the region between the mouth and the nasohypophyseal duct (Linked image shows neural crest derived structures in red, pink, dark green.) Miyashita argues that the elimination of the cyclostome upper lip by the separation of the nasal capsules and hypophyseal duct and the hypophyseal ducts migration to the roof of the mouth was a necessary prerequisite to their transformation into jaws, because they would confine these neural crest derivatives to the posterior part of the oral cavity. Given the lamprey-like face of osteostraci, this would have happened immediately prior to the actual appearance of jaws.
One has to be impressed by how many gnathostome synapomorphies this cascade of events encompasses:
- The ventral flexure of the forebrain causes the hypophyseal duct to move onto he roof of the mouth, eliminating the cyclostome "upper-lip."
- In its new position, the hypophyseal duct divides migrating neural crest into two streams that chondrify as trabeculae.
- All of which blocks anterior migration of neural crest that would become cyclostome anterior cartilages of face
2. New Branchial Arches:
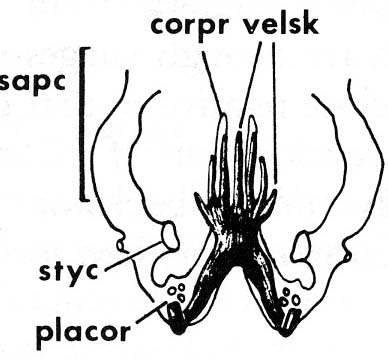
The lamprey velar skeleton bears on the issue of gnathostome branchial arches, too.Note:
- Lampreys' branchial arches are external to the gills
- Gnathostomes' are internal to them
- As gnathostomes, the principal arches of chondrichthyans (cartilaginous fish) are internal, however they also have thin extrabranchial cartilages lateral to their gills.
Lamprey velar skeleton in dorsal view
from Janvier, 1993 in Hanken and Hall. The Skull. Vol 2
As with jaws, Mallatt and Janvier square off:
- Mallatt proposes that the ancestors of vertebrates had both internal and external branchial arches. In lampreys the internal ones were lost and in gnathostomes, the external ones were lost.
- Janvier regards the internal gnathostome arches as evolutionary novelties of gnathostomes. However, he considers them to be based on the developmental pattern of the medial elements of the lamprey velar skeleton that has been developmentally shifted back into the branchial chambers.
Ontogeny: Developmentally, lampreys and gnathostomes are working with the same material: The gills form from blocks of endoderm that are surrounded top, bottom, medially, and laterally by a layer of migrating neural crest ectoderm. In gnathostomes, the neural crest cells develop into branchial arches medially because of an inductive relationship with adjacent gut-tube endoderm. In lampreys, neural crest apparently is triggered to form branchial arches by a similar relationship with regular ectoderm. Thus, the evolution of medial gill arches really reflects the evolution of a change in the way in which neural crest is signaled to make cartilage.
But note: Conventional wisdom holds that branchial arches are formed by neural crest and paired fins are formed from mesodermal mesenchyme. Sleight and Gillis, 2020, however, report that in the skate Leucoraja erinacea neural crest and mesenchyme collaborate in the formation of branchial arches, with neural crest predominating anteriorly and mesenchyme posteriorly. How taxonomically widespread this pattern is is a question for the future.
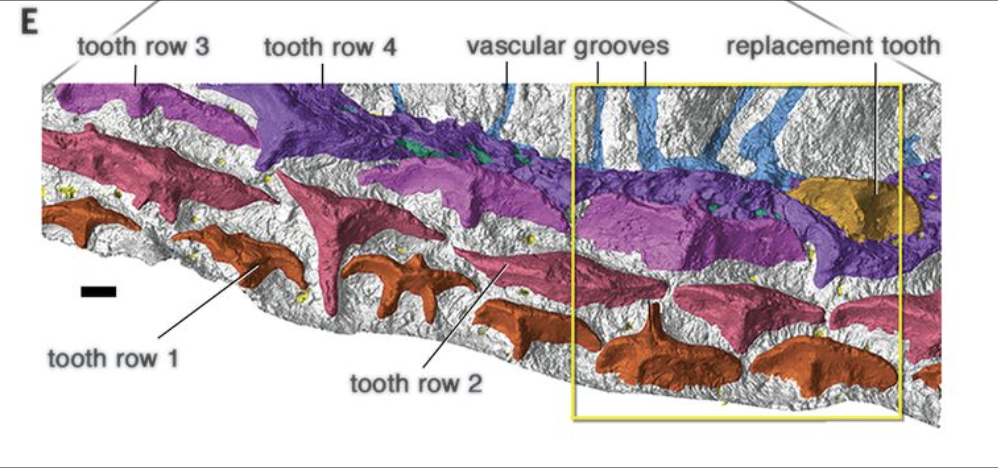
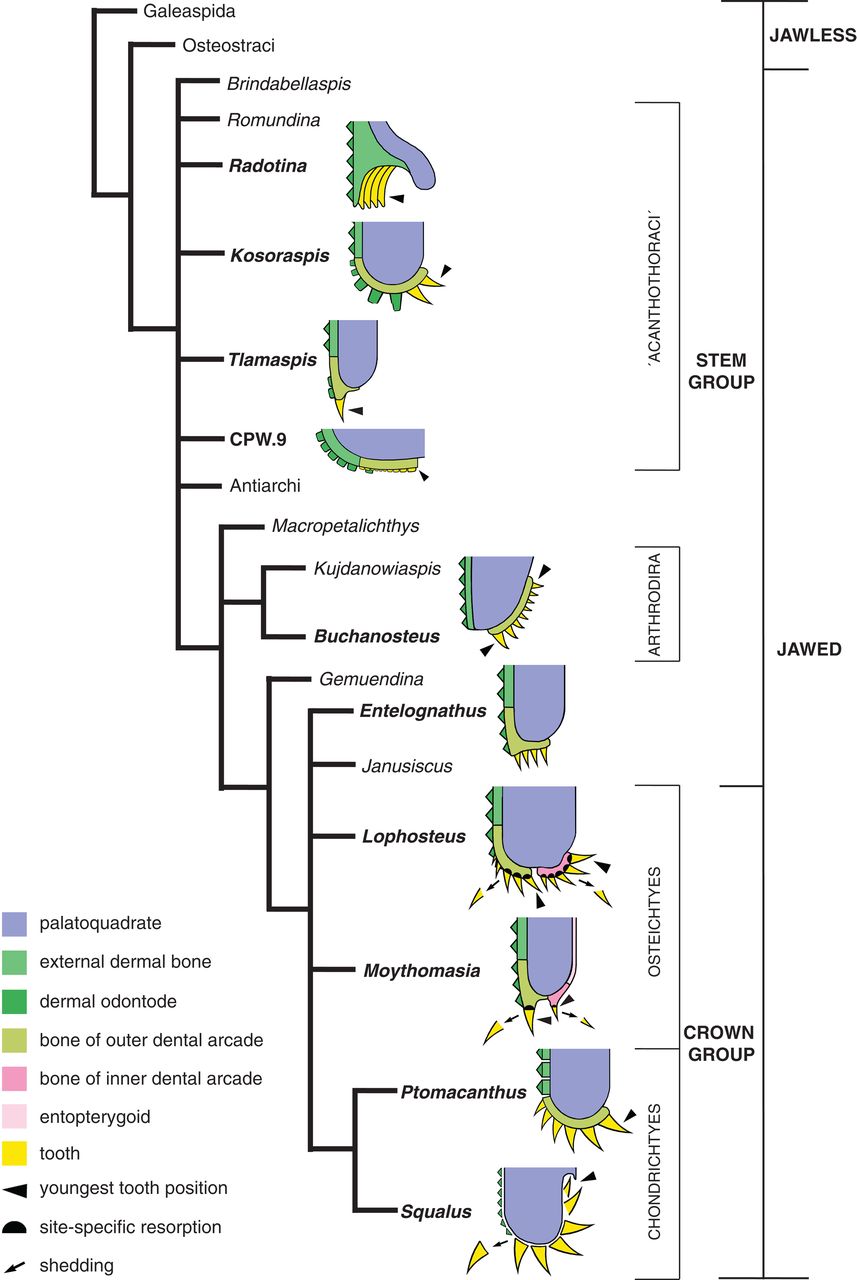
3. Teeth:
Until recently, proper teeth - odontodes lining the oral and pharyngeal cavities employed in feeding - were not recognized in any gnathostomes outside the crown Group of Gnathostomata. Recently, however, they have been recognized in several basal groups, especially members of Acanthothoracidae, enabling us to say:- They occur in tooth-families in which new teeth are added on the lingual (medial) side as the animal grows.
- Older teeth are not shed in the most basal gnathostomes
- Tooth families tend to be continuous with and grade into rows of dermal odontodes (placoid scales) on the outer surface (Vaškaninová et al., 2020)

4. The Pelvic Fins:
The evolution of distinct pectoral and pelvic fins is prefigured to some degree in the fin-folds of anaspids and thelodonts.- Lobe-shaped suprabranchial folds above the gill openings. (In osteostraci these have a proper skeleton.)
- Long uniform postbranchial folds stretching from posterior to the gill openings to near the anus.
- Pectoral fins originate higher on the body and attach by means of a small number of basal elements.
- Pelvic fins are often still elongate, and their skeleton consists of a long array of parallel fin rays.
- The fin-fold hypothesis, predicting that pectoral and pelvic fins arose simultaneously as expressions of the fin-fold.
- The archipterygium hypothesis, maintaining that the pectoral girdle and fin formed from an ancestral branchial arch.
- we know so little about the branchial arches of the most basal (placoderm-grade) gnathostomes, even though their heads and shoulder girdles are clearly distinct on the surface.
- in Osteostraci there is no distinct pectoral girdle, rather, pectoral fins simply hang off of a solid co-ossified head shield.
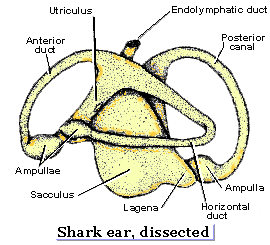
5. The Horizontal Semicircular Canal:
A true enigma. We see no intermediate forms. However:- In mice, mutations of the otx1 gene results in the failure of the canals to form.
- A stunning reversal: Maklad et al., 2014 have shown that two sets of horizontal semicircular canals are present in the lamprey Petromyzon marinus but that they form medially rather than laterally and are not homologous to the gnathostome canals.
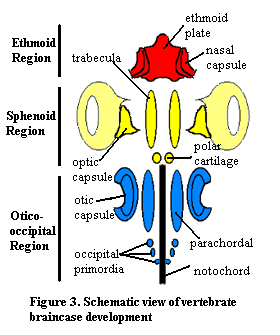
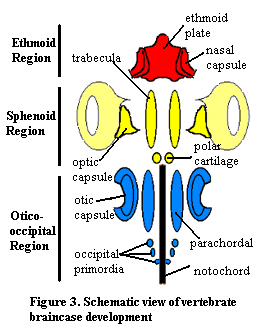
6. Braincase Development:
The braincase of adult gnathostomes ossifies in a variety of patterns, however the early development of the cartilages that give rise to it is relatively uniform. The first cartilages to appear give rise to the floor of the braincase or form the special sense capsules. From front to back we see:- Ethmoid region:
- Nasal capsules: Enclosing the olfactory organs.
- Ethmoid plate: (Neural crest ectoderm) Midline structure supporting the nasal capsules.
- Sphenoid region:
- Trabeculae: (Neural crest ectoderm) Neural crest ectodermal - Paired rods giving rise to the floor of the front of the braincase between the optic capsules
- Eyes
- Occipito-otic region:
- Otic capsules: (Mesoderm) Enclosing the membranous labyrinth.
- Parachordals: (Mesoderm) Medial to otic capsules and lying alongside the notochord.
- Occipital arch: An upside-down U forming the posterior end of the braincase enclosing the foramen magnum through which the spinal cord exits the braincase.
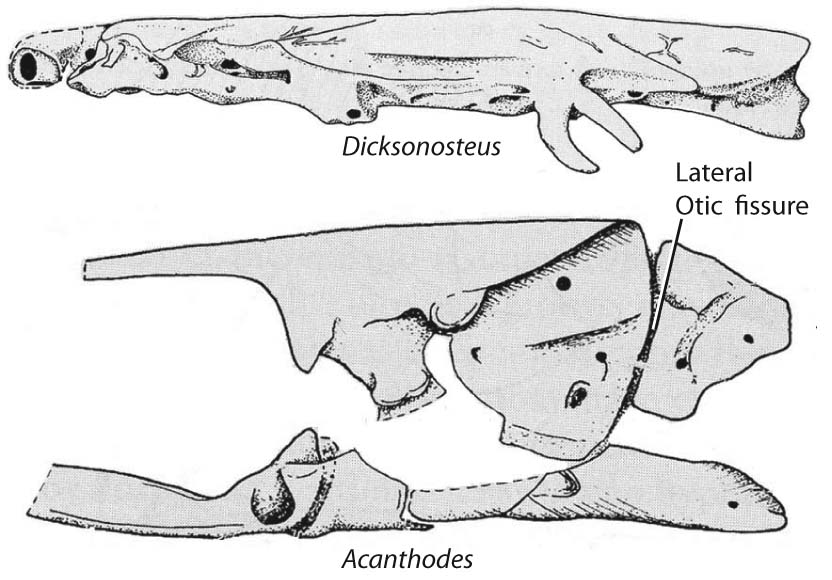
All vertebrates have the special sense capsules (albeit only one nasal capsule for some basal vertebrates) and parachordals. Only gnathostomes definitely have the remainder (maybe galeaspids and osteostracans). The gnathostome braincase encloses the exits of the twelve cranial nerves. The occipital arch is believed to have originated from a short series of arcualia or vertebrae that became incorporated into the rear of the skull. Note that the otic capsules and occipital arches do not fuse well, resulting in a persistent lateral otic fissure.
Gnathostome diversity:
Traditionally, three major groups of unknown relationships were recognized:
- Placodermi - Silurian - Devonian forms with extensive bony armor
- Acanthodii - Silurian - Permian forms with extensive bony armor
- Chondrichthyes - living cartilaginous fish and their fossil relatives
- Osteichthyes - living bony fish and their fossil relatives

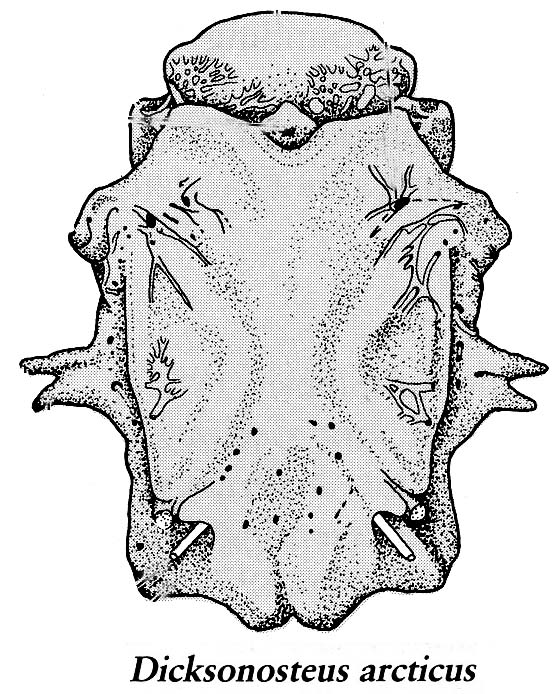
Placodermi:
(Silurian - Devonian) Lovely and entertaining armored gnathostomes and poster-children for the Devonian. Experienced a rapid worldwide diversification at the beginning of the Devonian and abrupt decline and extinction at its end.Identifying features:
- Distinct cranial and thoracic armor with unique ossification pattern. The cranial and thoracic shields often meet at a mobile joint.
- Jaws lined with gnathals, self-sharpening occluding bony plates.
- Teeth? Recent investigations have identified smaller odontodes in the gnathal plates of some placoderms:
- Acanthothoracans - (basal placoderms) (Rücklin and Donoghue, 2015; Vaškaninová et al., 2020), where odontodes are added during growth to the peripheral margin of broad gnathal plates.
- Arthrodires - (derived placoderms) (Rücklin et al., 2012), in which the odontodes form a linear array.
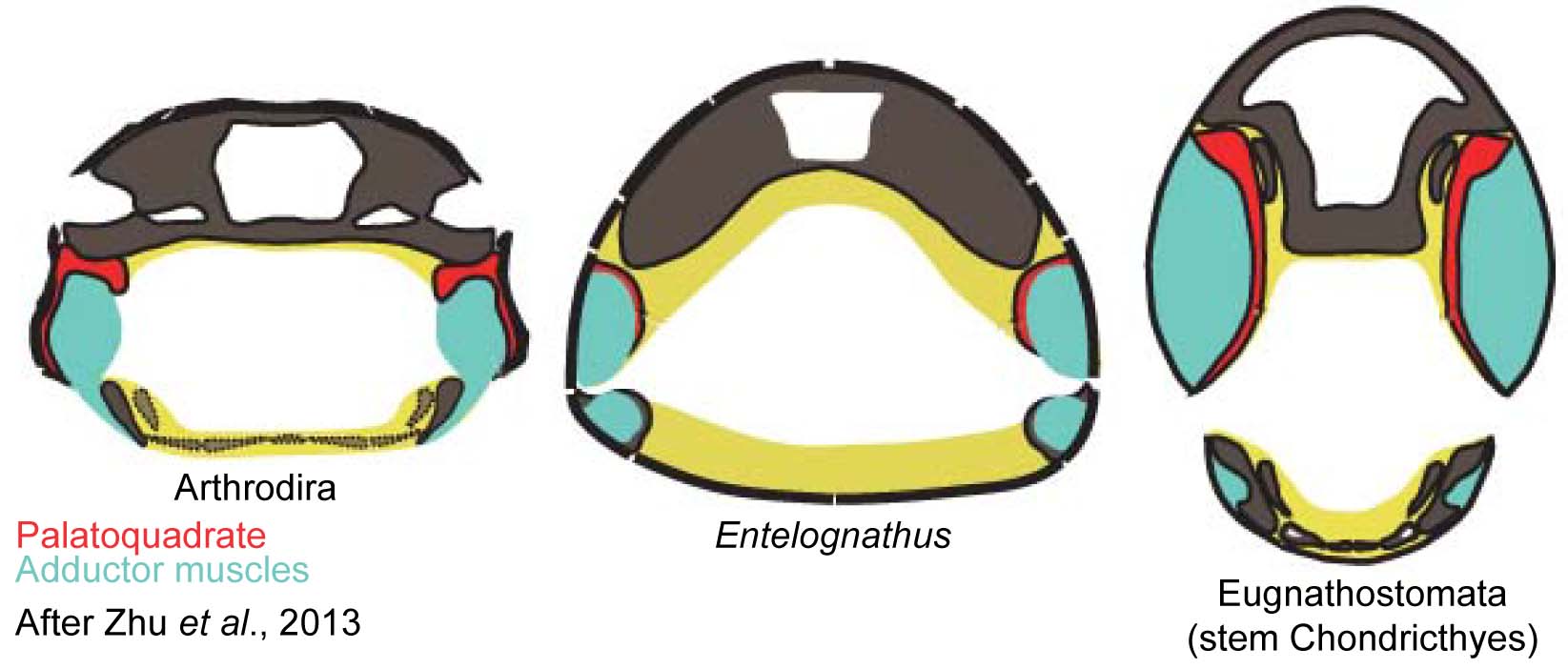
- Palatoquadrate attaches to the dermal bones of the skull roof, causing adductor muscles (that close the jaw) to pass medial to (inside) palatoquadrate, in stark contrast to other gnathostomes.
- The ethmoid region tends to ossify separately from the remainder of the braincase.
- Compared to other gnathostomes, the otico-occipital region takes up much more of the length of the braincase (a plesiomorphy!).
Phyllolepis by Dmitri Bogdanov
- Many have pectoral skeletons sporting corneal processes like those of galeaspids or osteostracans.
- Among Cyclostomi, Galeaspida, and Osteostraci the nasal capsules (or capsule) are crowded in front of a relatively short forebrain, while the upper lip is expansive. Basal placoderms including Acanthothoraci and Antiarchi show similar facial proportions whereas more derived gnathostomes have longer forebrains and nasal capsules that lie directly adjacent to the upper lip. (Dupret et al., 2014).

Janvier, Phillipe. 1993. Early Vertebrates - find the two placoderms
Placoderms were very diverse and occupied many ecological roles. During the Devonian, they greatly outnumbered other marine fish (right). Their specializations included:
- Shellfish feeding with crushing dental plates.
- Bottom feeding.
- Predation: Some of the largest predatory fish ever, such as Dunkleosteus. Many seem suited to resting on the bottom and engulfing prey by rapidly raising their head and opening jaws. Some localities, however, preserve more streamlined placoderms like: The latter two are especially interesting for displaying elongated upper and lower jaws respectively that prefigure later vertebrates using similar strategies.
- Deposit feeding.
- Ram suspension-feeding: Titanichthys, although the size of Dunkleosteus has a much more slender jaw. A morphometric analysis by Coatham et al., 2020 indicates that ecologically, it was specialized for suspension-feeding in the manner of a basking shark.
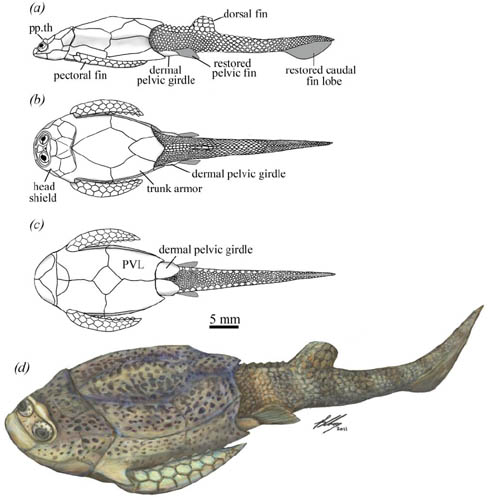
Reproduction:
Some placoderms are known to have practiced internal fertilization (the earliest known instance). Long suspected because some individuals had structures looking like "claspers" - the intromittant organs of chondrichthyans.
- In 2009, Long et al. described the fossil of a pregnant placoderm confirmed this.
- the claspers of sharks are definitely enhancements of their pelvic fins,
- those of placoderms are separate and distinct paired structures behind the pelvic fin.
Fossil record: Fragmentary record begins in the Early Silurian, followed by a rapid diversification. During the Devonian, placoderms were the dominant vertebrate group. Both marine and fresh water forms are recorded with a worldwide distribution except for absence in South America, which hosted south polar glaciers. Placoderm diversity was greatly reduced by the Late Devonian Frasnian extinction event, and they were completely extinguished by the Fammenian event at the end of the Devonian. Thus, entire radiation took up only about 77 million years, but while it lasted, it was spectacular. But maybe -
Placoderms look different and distinct, and for over a century, were assumed to be monophyletic, so it came as a surprise when Brazeau, 2009 and others began recovering them as paraphyletic. Today we regard them as basal gnathostomes. Recent phylogenies differ in detail but generally agree on which placoderm groups are closer to the base of Gnathostomata and which are closer to the crown. (Though King et al., 2016 find paraphyletic and monophyletic Placodermi to be equally parsimonious.) A sampling of some placoderm high-points:
Xiushanosteus mirabilis: (Early Silurian) The oldest known placoderm. Phylogenetically basal, but not the most basal. Abundant where present, this creature is similar to acanthothoracid placoderms, but is tiny.. Note 5 mm scale bar in image at right!
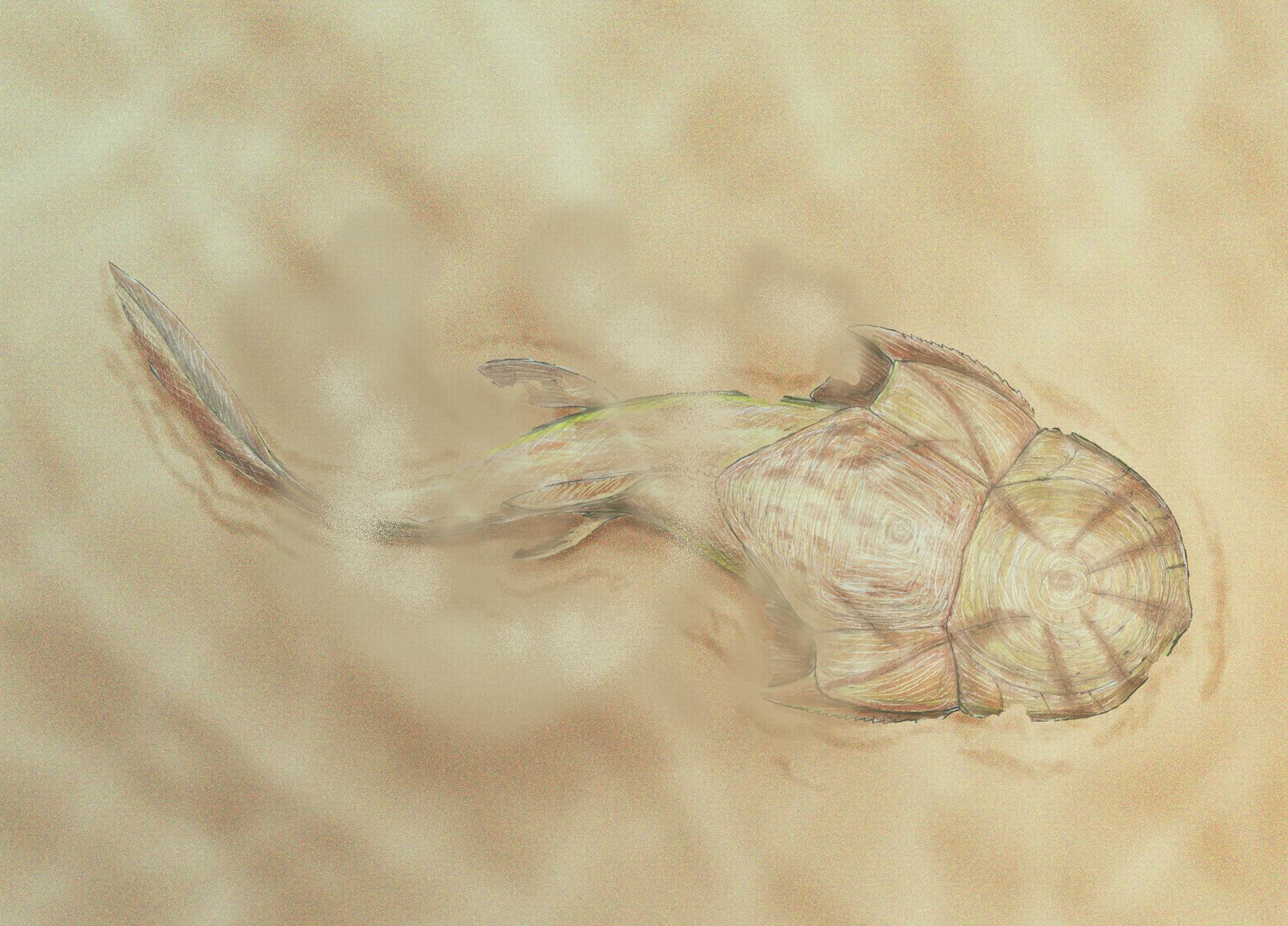
Stensioella: (Early Devonian) Among the most basal gnathostomes. Although poorly known, gnathostome synapomorphies including paired nares and pelvic fins are apparent. Features:

- The head and thoracic shields are seen to be an array of individual tesserae that have not firmly coalesced into characteristic placoderm head plates.
- Pectoral fins are large.
- Tail is long and whip-like.
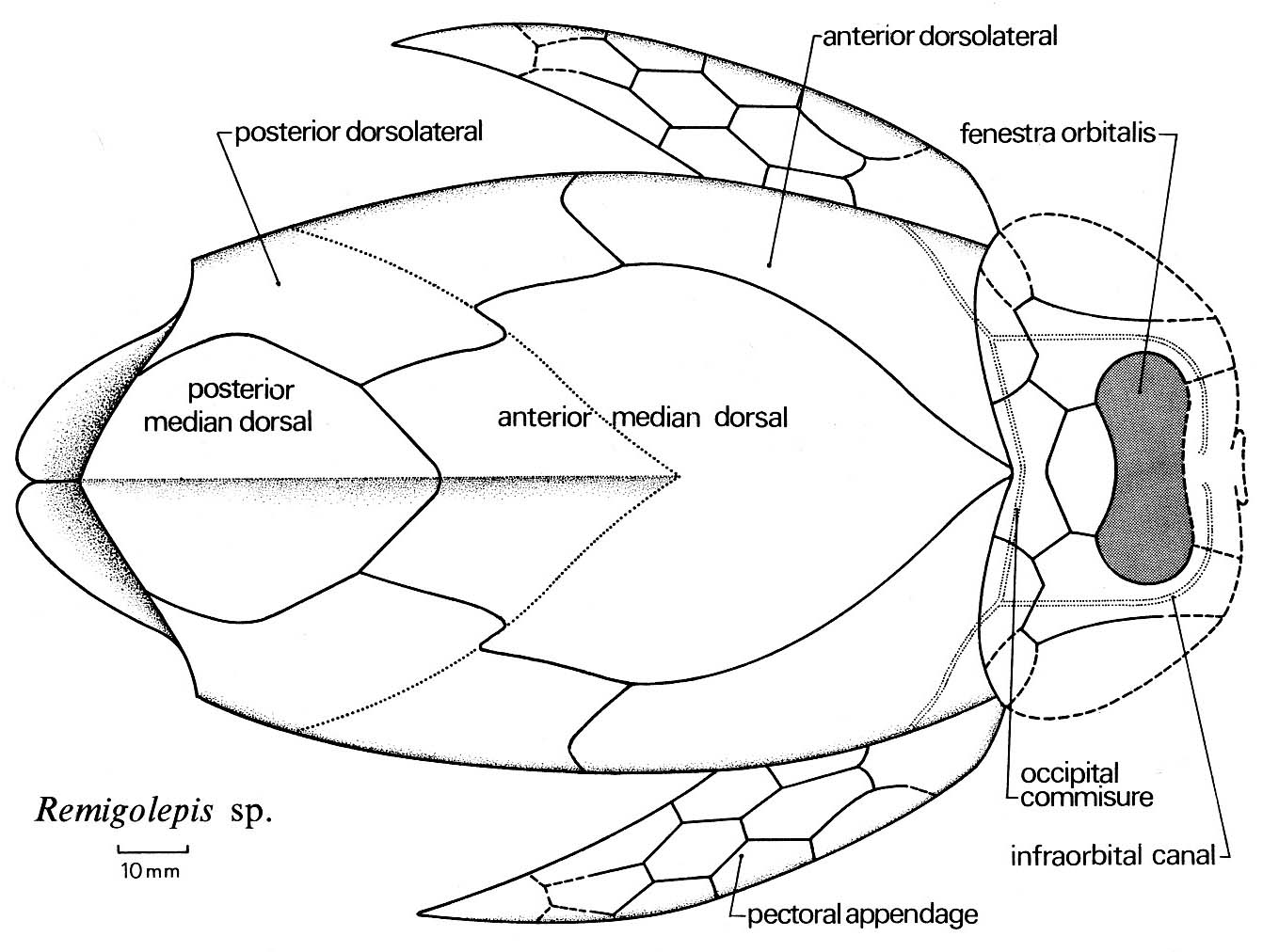
Antiarchi: (Silurian - Devonian) Diverse and successful weirdos (second only to Arthrodira in diversity) common both in sea and fresh-water environments. Key features:
- Extensive thoracic armor forming a box from which the tail (scaled or naked) protruded. The thoracic armor regularly contains elements including a posterior medial dorsal that other placoderms lack.
- Cranial armor articulates with thoracic by a condyle (knob) that fits into a thoracic socket.
- Orbits, nares, and pineal foramen are all confined to a small orbital fenestra in the top of the head.
- Strangest of all, the pectoral fins are armored with dermal bone plates. In derived taxa, these have an articulated joint at mid-length. Older and more basal antiarchs like Parayunnanolepis have shorter unjointed pectoral fins.
- Anal fin absent.
- The presence or absence of pelvic fins in antiarchs has long been debated. Zhu et al.'s 2012 examination of Parayunnanolepis indicate that pelvic fins, attached to a pelvic girdle with both dermal and endochondral elements are definitely present. Note: Zhu et al., in press cast some doubt on the presence of pelvic endochondral elements in this taxon.
- The clustering of orbits, openings to the nasal capsule(s) and pineal foramen on top of the head is similar in both groups. (See osteostracan)
- Patterns of vascularization (blood supply) and nerves in the thoracic armor where these articulate with pectoral fins are very similar to those of osteostraci.
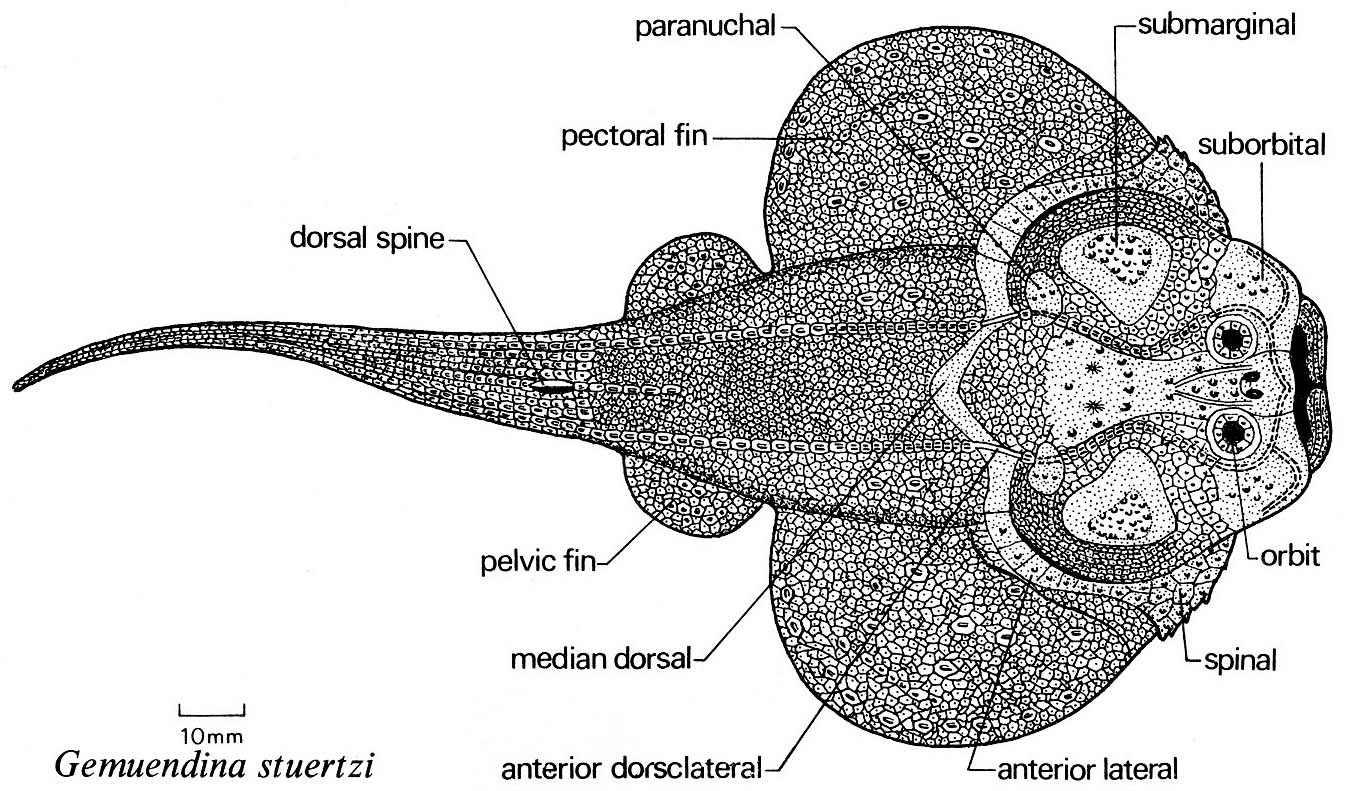
Rhenanida: (Devonian) Secondarily flattened bottom dwellers with large pectoral fins. We have seen the "bottom-dweller" ecomorph before and will see it again. Rhenanids are the first gnathostomes to take up that life style through the enlargement of their pectoral fins. Features:

- Enlarged pectoral fins
- Reduction of bony plates to small tesserae (obscuring the homologies of all but a few.)
- Jaw plates flattened and covered with small denticles
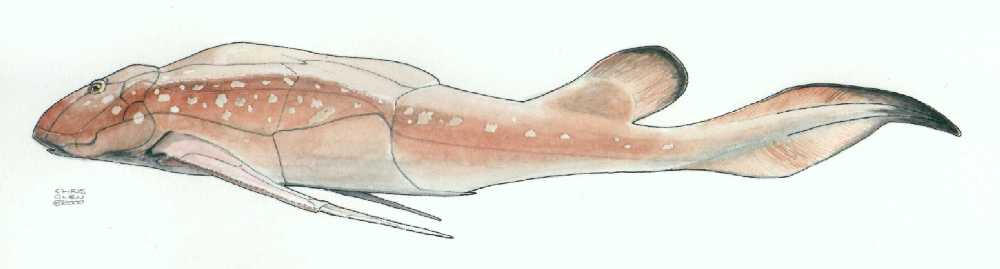
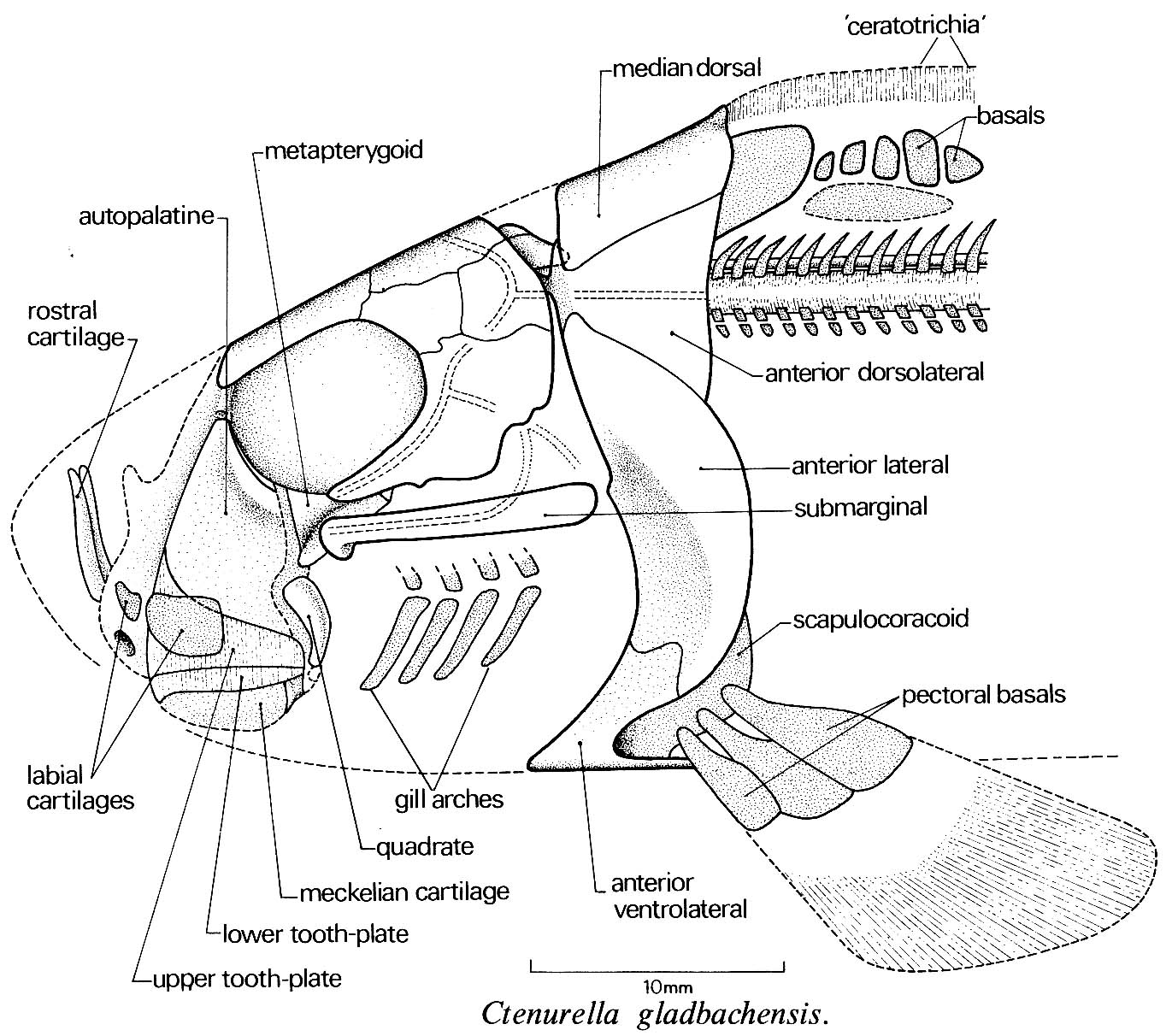
Ptyctodontida: (Devonian) Strongly similar to living ratfish. Features:

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
- Significantly reduced cranial and thoracic armor, in which the bone was mostly covered with skin.
- Tooth plates have a spongy microscopic texture
- These are the placoderms in which we see both the sexual dimorphism of the pelvic region mentioned above and evidence for viviparity.
{kind=link}

Eastmanosteus calliaspis
{kind=link}
- Mobile joint between socket on cranial armor and condyle on thoracic.
- A large nuchal gap between the two shields allows the head to be elevated to increase the gape.
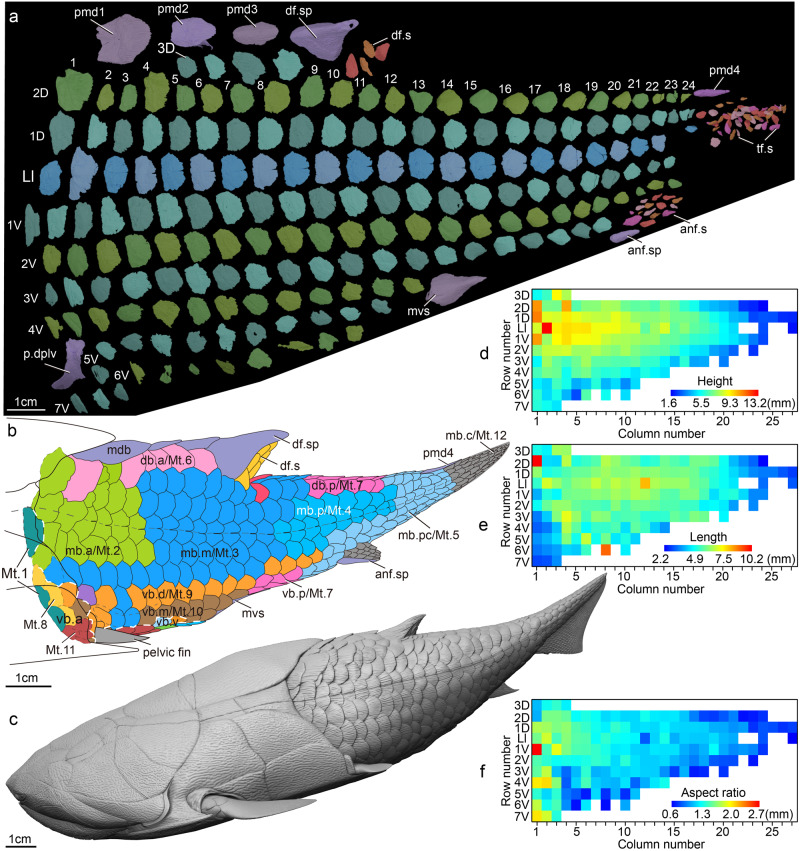
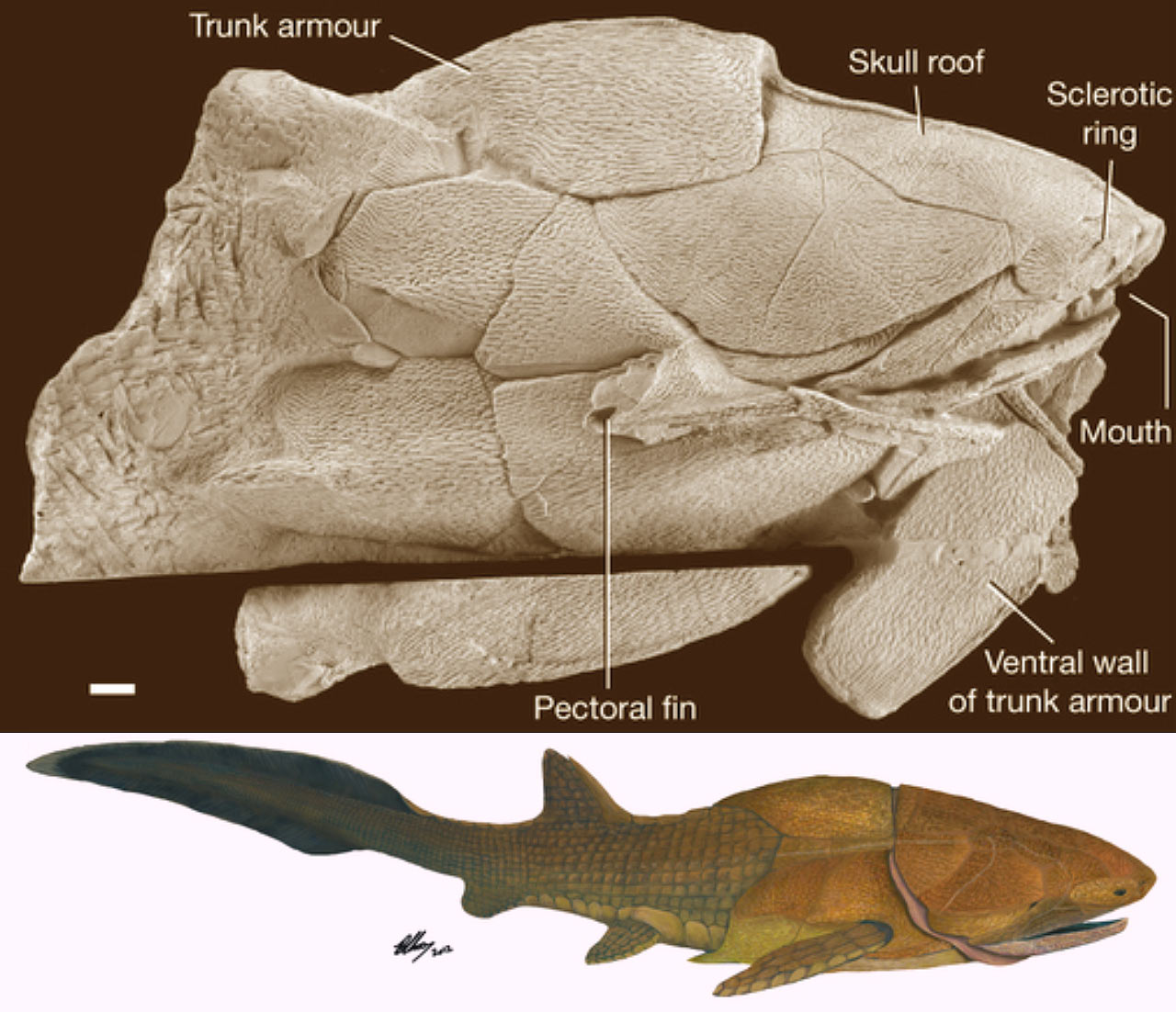
- The heads and thoracic girdles of placoderms have an excellent record. Not so much for the rest of the animal, however among Arthrodira, we begin to pick up a good sense of the body profile of derived placoderms, including Amazichthys (Jobbins et al., 2022) and Entelognathus (Cui et al., 2023). These are not dissimilar from basal crown gnathostomes.
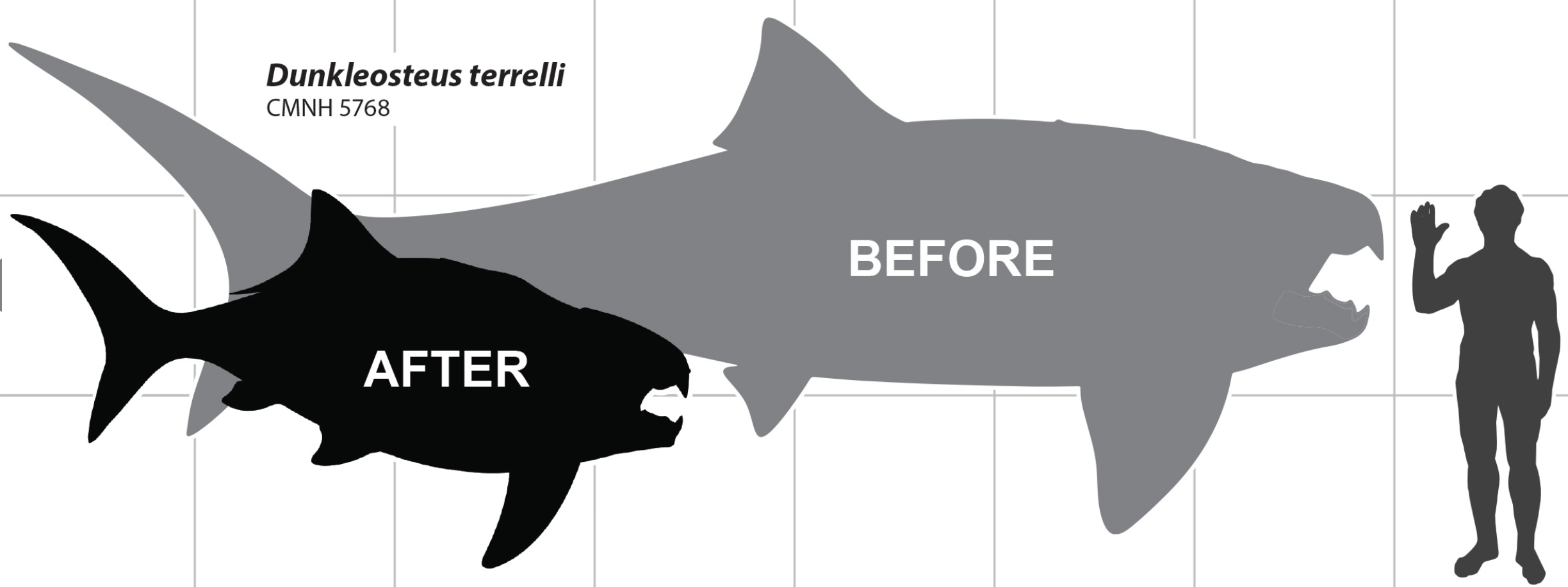
- And yet the profiles of even familiar taxa remain enigmatic. Engelman, 2023 notoriously "shrunk the dunk", employing an orbit to operculum metric to greatly reduce length estimates for Dunkleosteus. Interesting speculation awaiting physical evidence.
{kind=link}
{kind=link}
{kind=link}
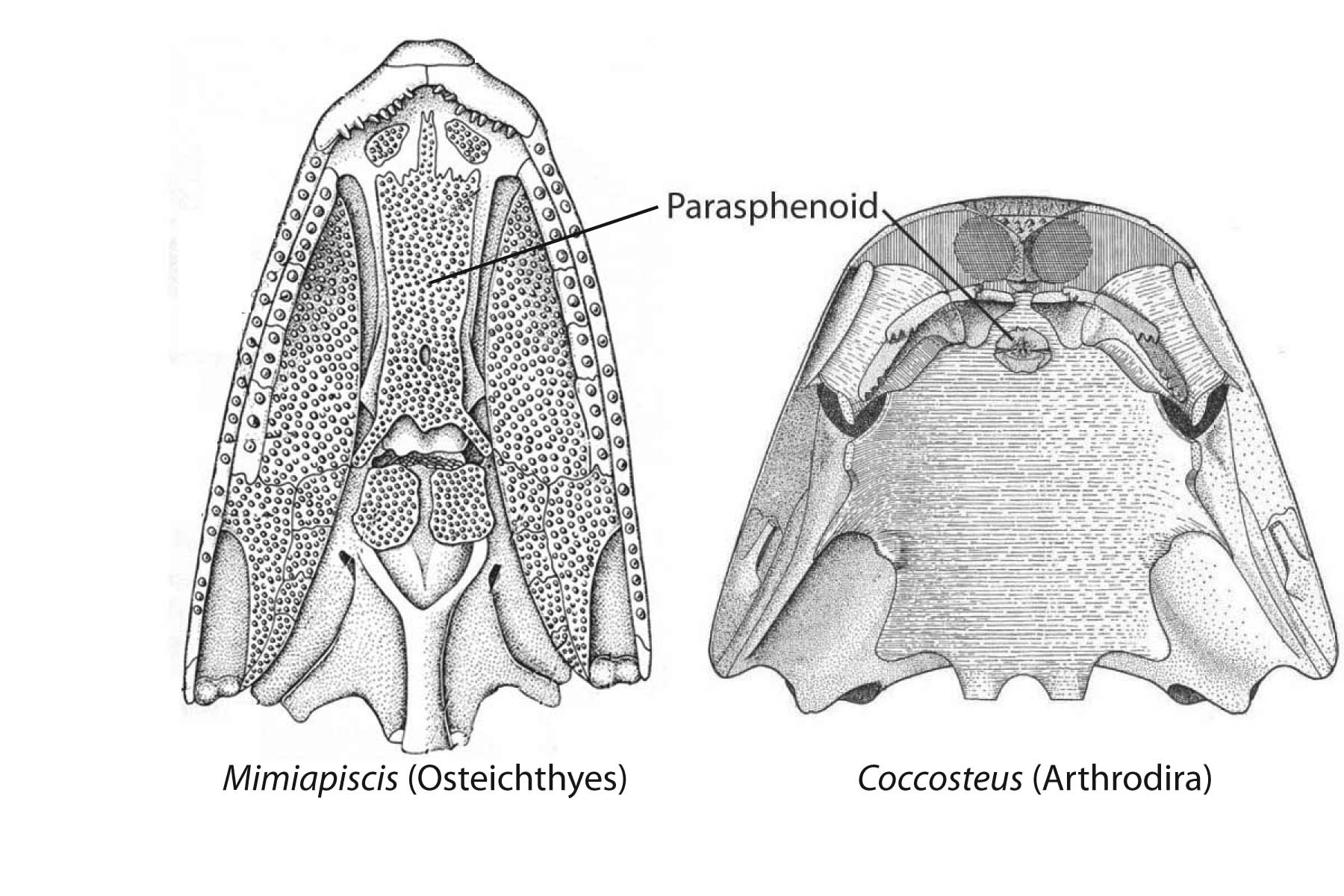
Potential synapomorphy of Arthrodira and Osteichthyes:
- The parasphenoid - a dermal element of the palate that forms beneath the sphenoid region of the neurocranium. (Which other placoderm-grade dermal bones are homologous with those of Ostiechthyes is an area of active research.)
{kind=link}

Entelognathus primordialis
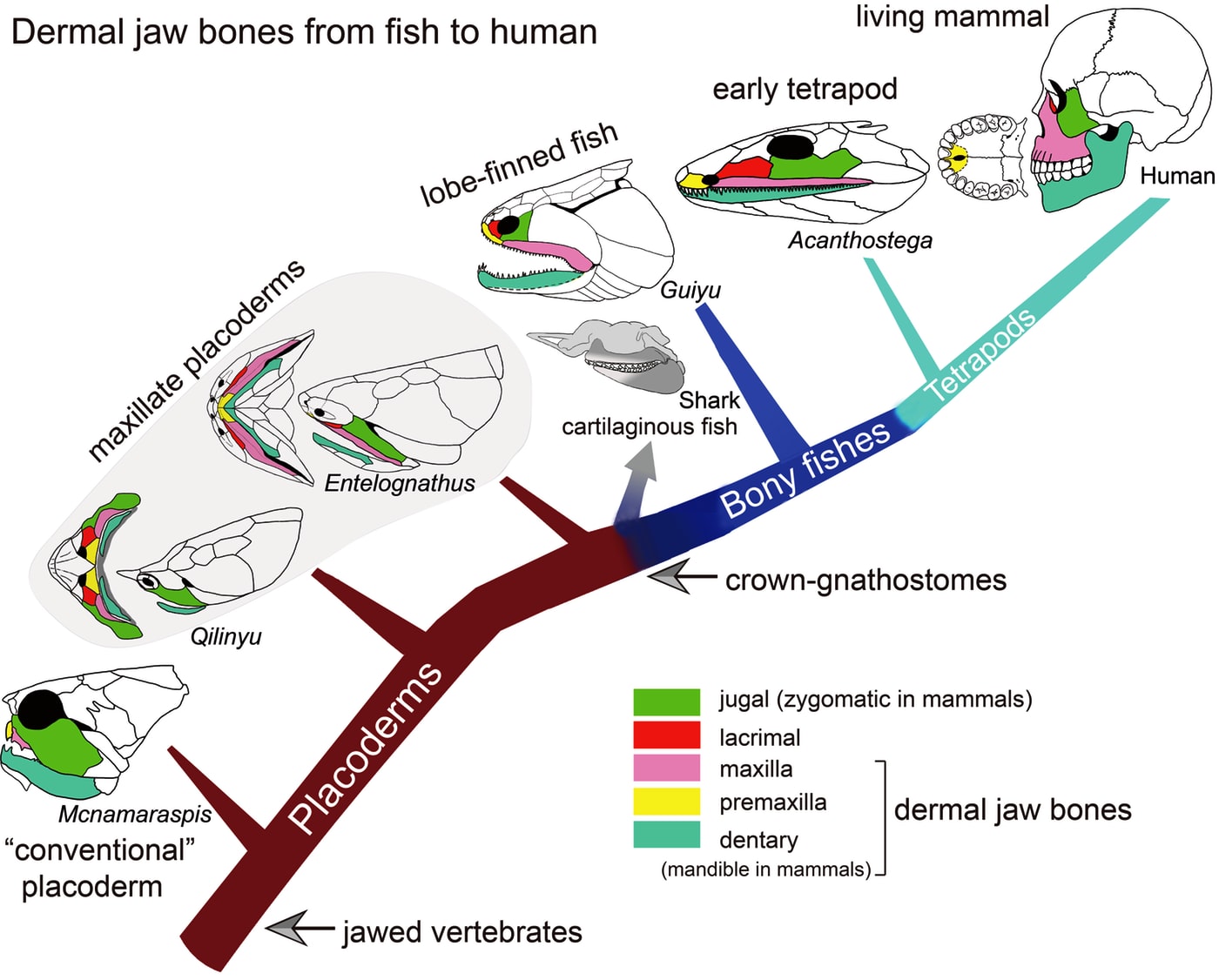
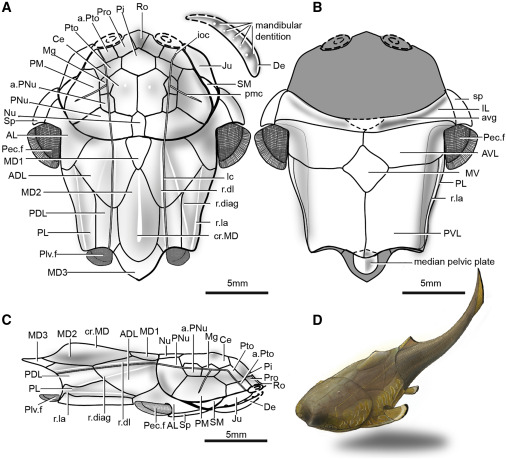
Entelognathus primordialis: (Silurian) Zhu et al., 2013. described this unremarkable creature with a extensive thoracic shield. Entelognathus looks like an arthrodire without the hinged articulation of head and thoracic armor or nuchal gap.
What makes Entelognathus interesting is its possession of potential synapomorphies with Osteichthyes:
- Three dermal bones of the face:
- Maxilla
- Premaxilla
- Dentary
- Much of the adductor muscle of the jaw is lateral to the palatoquadrate.
{kind=link}
{kind=link}
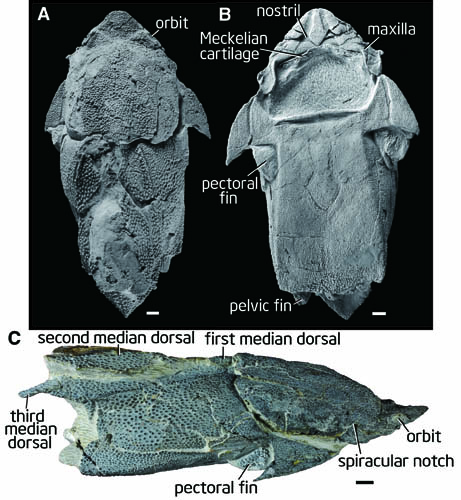
Qilinyu rostrata
- Qilinyu rostrata, (Silurian) a second "maxillate" placoderm-like fish. Based on Qilinyu, Zhu et al conclude that the premaxilla, maxilla, and dentary are homologous to placoderm gnathal plates. This identification facilitates the further identification of the jugal and lacrimal in typical placoderms. (Zhu et al, 2016)
- Bianchengichthys micros, (Silurian) a third. In this one, the premaxilla, maxilla, and dentary retain arthrodire-like teeth. (Li et al., 2021)
{kind=link}
Entelognathus and Qilinyu, with their extensive armor, were not what we expected of close relatives of living gnathostomes, whose ancestral body covering, we had long presumed, consisted of small scales and plates. Turns out that's wrong. In fact, Entelognathus and its friends have overturned much of our understanding of early gnathostome evolution. We take that up in the next lecture.
- Martin Brazeau, 2009 The braincase and jaws of a Devonian "acanthodian" and modern gnathostome origins Nature 457, 305-308.
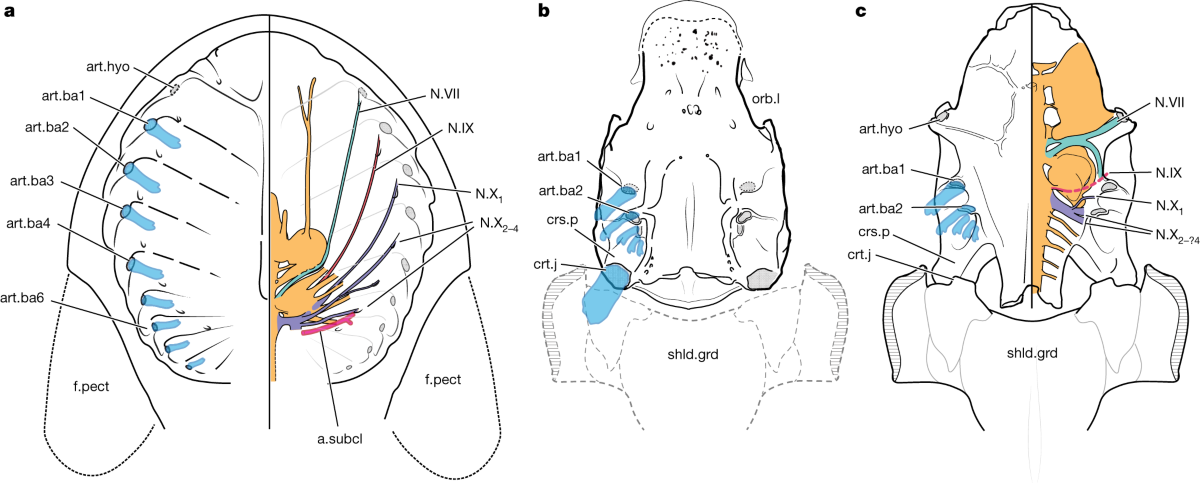
- Martin D. Brazeau, Marco Castiello, Amin El Fassi El Fehri, Louis Hamilton, Alexander O. Ivanov, Zerina Johanson and Matt Friedman, 2023. Fossil evidence for a pharyngeal origin of the vertebrate pectoral girdle Nature 623, 550-554.
- Samuel J. Coatham, Jakob Vinther, Emily J. Rayfield and Christian Klug. 2020. Was the Devonian placoderm Titanichthys a suspension feeder?. Royal Society Open Science 7200272.
- Simon Conway Morris and Jean-Bernard Caron. 2014. A primitive fish from the Cambrian of North America. Nature, 512, 419-422.
- Xindong Cui, Matt Friedman, Yilun Yu, You-an Zhu, Min Zhu. 2023. Bony-fish-like scales in a Silurian maxillate placoderm. Nature Communications, 2023 Nov 22;14(1):7622.
- J. Vincent Dupret, Sophie Sanchez, Daniel Goujet, Paul Tafforeau, and Per E. Ahlberg. 2014. A primitive placoderm sheds light on the origin of the jawed vertebrate face. 507, pages500–503 (2014).
- by Russell K. Engelman. 2023. A Devonian Fish Tale: A New Method of Body Length Estimation Suggests Much Smaller Sizes for Dunkleosteus terrelli (Placodermi: Arthrodira). Diversity 15(3), 381.
- J. Zhikun Gai, Min Zhu, Per Ahlberg, Philip Donoghue. 2022. The Evolution of the Spiracular Region From Jawless Fishes to Tetrapods. Frontiers in Ecology and Evolution, Sec. Paleontology Volume 10.
- J. Andrew Gillis, Olivia R.A. Tidswell. 2017. The Origin of Vertebrate Gills. Current Biology, February 9, 2017.
- Christine Hirschberger, Victoria A Sleight, Katharine E Criswell, Stephen J Clark, J Andrew Gillis. 2021. Conserved and unique transcriptional features of pharyngeal arches in the skate (Leucoraja erinacea) and evolution of the jaw. Molecular Biology and Evolution, 38(10): 4187–4204.
- Christine Hirschberger and J Andrew Gillis. 2022. The pseudobranch of jawed vertebrates is a mandibular arch-derived gill. Development 149(13).
- Philippe Janvier, 1993. Patterns of diversity in the skull of jawless fishes. In The Skull: Patterns of Structural and Systematic Diversity (Volume II, eds J. Hanken and B. K. Hall), pp. 131–188. The University of Chicago Press: Chicago.
- Philippe Janvier, 1996. Early Vertebrates. Oxford University Press.
- Melina Jobbins, Martin Rücklin, Humberto G. Ferron, and Christian Klug, 2022. A new selenosteid placoderm from the Late Devonian of the eastern Anti-Atlas (Morocco) with preserved body outline and its ecomorphology. Frontiers in Ecology and Evolution 10-2022.
- Melina Jobbins, Martin Rücklin, Marcelo R. Sánchez Villagra, Hervé Lelièvre, Eileen Grogan, Piotr Szrek and Christian Klug, 2024. Extreme lower jaw elongation in a placoderm reflects high disparity and modularity in early vertebrate evolution. Royal Society Open Science 11(1).
- Benedict King, Tuo Qiao, Michael Lee, Min Zhu, and John Long. 2016. Bayesian morphological clock methods resurrect placoderm monophyly and reveal rapid early evolution in jawed vertebrates. Systematic Biology 0(0) 1-18.
- Shigeru Kuratani and Per. E. Ahlberg. 2018. Evolution of the vertebrate neurocranium: problems of the premandibular domain and the origin of the trabecula. Zoological Letters 4(1).
- Qiang Li, You-an Zhu, Jing Lu, Lijian Peng, Guangbiao Wei, Min Zhu. 2021. A new Silurian fish close to the common ancestor of modern gnathostomes. Current Biology, 31(18), 3613-3620
- John Long, Kate Trinajstic, and Zerina Johanson. 2009. Devonian arthrodire embryos and the origin of internal fertilization in vertebrates. Nature, 457, 1124-1127
- A. Maklad, C. Reed, N Johnson, B Fritsch, 2009. Anatomy of the lamprey ear: morphological evidence for occurrence of horizontal semicircular ducts in the labyrinth of Petromyzon marinus. Journal of Anatomy, 224(4):432-46.
- Jon Mallatt. 1984. Early vertebrate evolution: pharyngeal structure and the origin of gnathostomes. Journal of Zoology. 204: 169-183.
- Tetsuo Miyashita. 2015. Fishing for jaws in early vertebrate evolution: a new hypothesis of mandibular confinement. Biological Reviews. 91(3): 611-657.
- Martin Rücklin and Philip Donoghue. 2015. Romundina and the evolutionary origin of teeth. Biology Letters, 11: 20150326. http://dx.doi.org/10.1098/rsbl.2015.0326.
- Martin Rücklin, Philip Donoghue, Zerina Johanson, Kate Trinajstic, Federica Marone, andMarco Stampanoni. 2012. Development of teeth and jaws in the earliest jawed vertebrates. Nature 491, 748-751.
- Victoria A Sleight and J Andrew Gillis. 2020. Embryonic origin and serial homology of gill arches and paired fins in the skate, Leucoraja erinacea. eLife 2020;9:e60635
- Kate Trinajstic, Catherine Boisvert, John Long, Anton Maksimenko and Zerina Johanson. 2015. Pelvic and reproductive structures in placoderms (stem gnathostomes). Biological Reviews, 90(2): 467-501
- Valeria Vaškaninová, Donglei Chen, Paul Tafforeau, Zerina Johanson, Boris Ekrt, Henning Blom, and Per Erik Ahlberg. 2020. Marginal dentition and multiple dermal jawbones as the ancestral condition of jawed vertebrates. Science, 369(6500): 211-216
- Hiromasa Yokoyama, Miho Yoshimura, Daichi G. Suzuki, Hiroki Higashiyama, Hiroshi Wada. 2020. Development of the lamprey velum and implications for the evolution of the vertebrate jaw. Developmental Dynamics, 250(1): 88-98
- Min Zhu, Xiaobo Yu, Brian Choo, Junqing Wang, and Liantao Jia. 2012. An antiarch placoderm shows that pelvic girdles arose at the root of jawed vertebrates. Biology Letters. 8(3): 453-456..
- Min Zhu, Xiaobo Yu, Per Erik Ahlberg, Brian Choo, Jing Lu, Tuo Qiao, Qingming Qu, Wenjin Zhao, Liantao Jia, Henning Blom & You'an Zhu, 2013 The braincase and jaws of a Devonian "acanthodian" and modern gnathostome origins. Nature 502, 188-193.
- Min Zhu, Per Erik Ahlberg, Zhaohui Pan, Youan Zhu, Tuo Qiao, Wenjin Zhao, Liantao Jia, Jing Lu, 2016. A Silurian maxillate placoderm illuminates jaw evolution. Science 354(6310) 334-336.
- You-an Zhu, Qiang Li, Jing Lu, Yang Chen, Jianhua Wang, Zhikun Gai, Wenjin Zhao, Guangbiao Wei, Yilun Yu, Per E. Ahlberg and Min Zhu. The oldest complete jawed vertebrates from the early Silurian of China. Nature 609, 954–958.
- You-An Zhu, Ya-Jing Wang, Qing-Ming Qu, Jing Lu, Min Zhu, in press. The pelvic morphology of Parayunnanolepis (Placodermi, Antiarcha) revealed by tomographic data. Vertebrata PalAsiatica DOI: 10.19615/j.cnki.2096-9899.221126.
- Martin D. Brazeau, Marco Castiello, Amin El Fassi El Fehri, Louis Hamilton, Alexander O. Ivanov, Zerina Johanson and Matt Friedman, 2023. Fossil evidence for a pharyngeal origin of the vertebrate pectoral girdle Nature 623, 550-554.