Euryapsida and kin - the reptilian invasion of the seas
Eventually we will move to the Archosauromorph side of the reptilian tree, but first, a problem.....

We risk neglecting a reptilian radiation that was both:
- Ecologically significant: The reptilian conquest of the open oceans
- Historically significant as it contained the first fossil reptiles to be scientifically studied, starting in the early 19th century before anyone had heard of a dinosaur,
- and are well-known to any child with a book of prehistoric animals:
- Ichthyosaurs
- Plesiosaurs
- Placodonts
- Thalattosaurs
- Saurosphargids
- other odd-balls
- but NOT:
- mosasaurs and dolichosaurs, which were squamates
- pleurosaurs, which were rhynchocephalians
- sea turtles, which are (surprise) turtles.
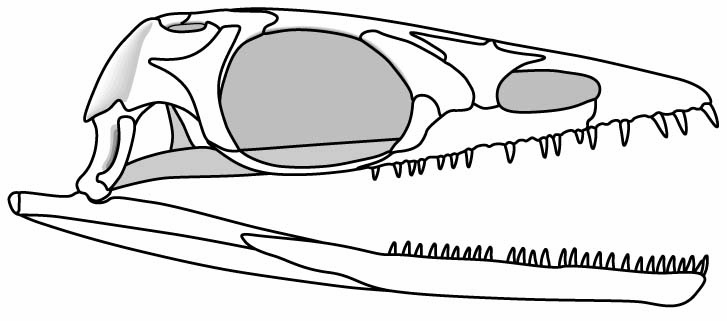
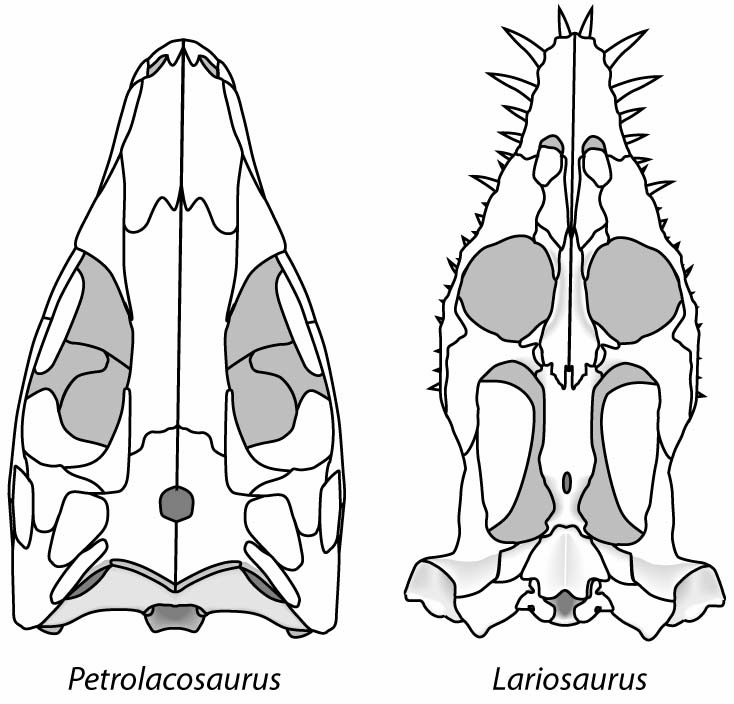
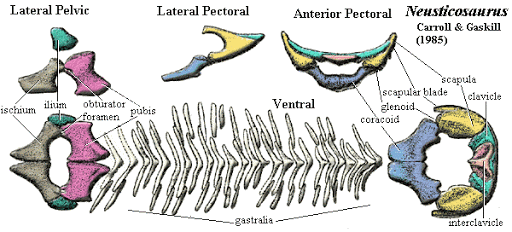
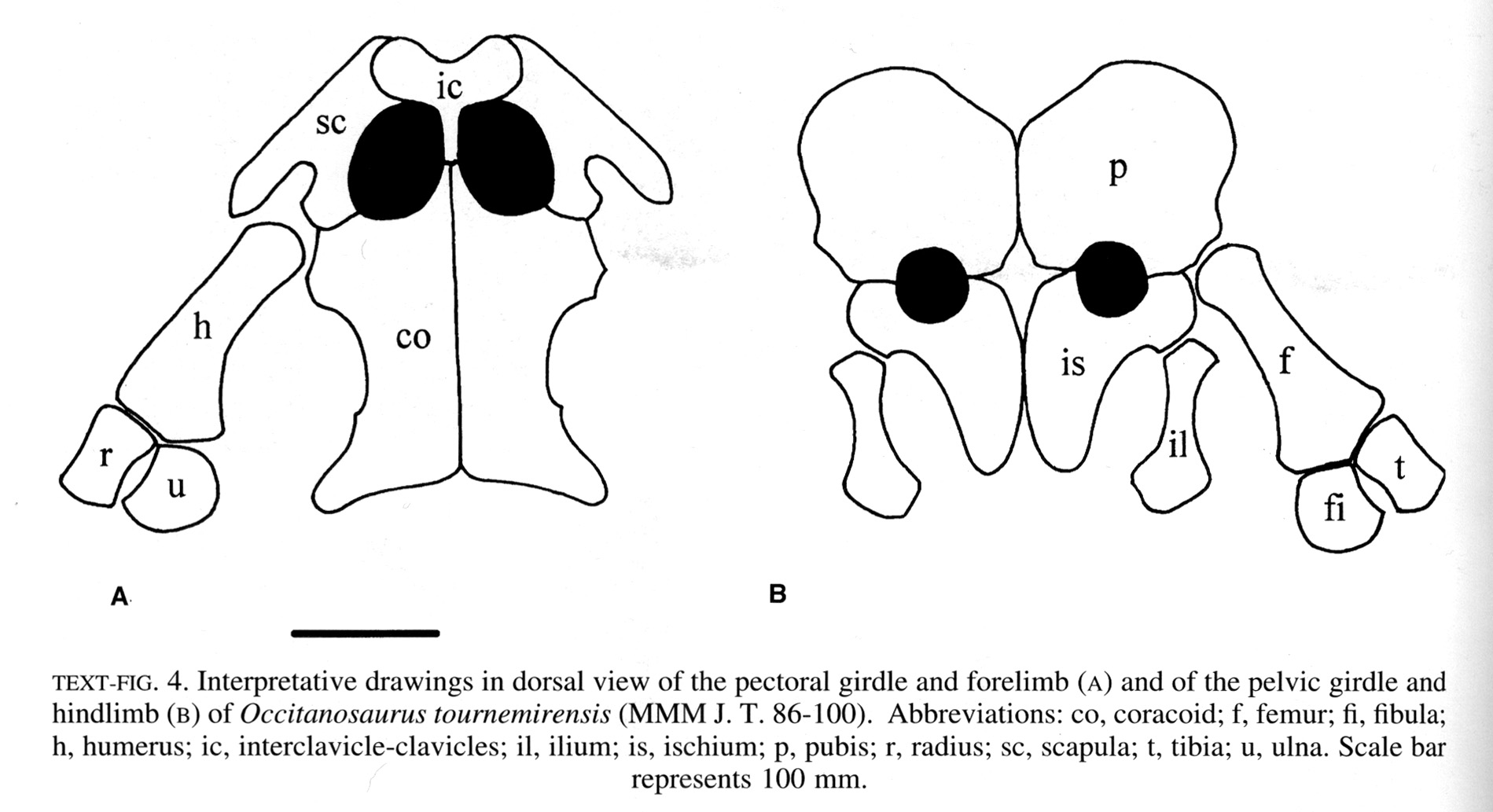
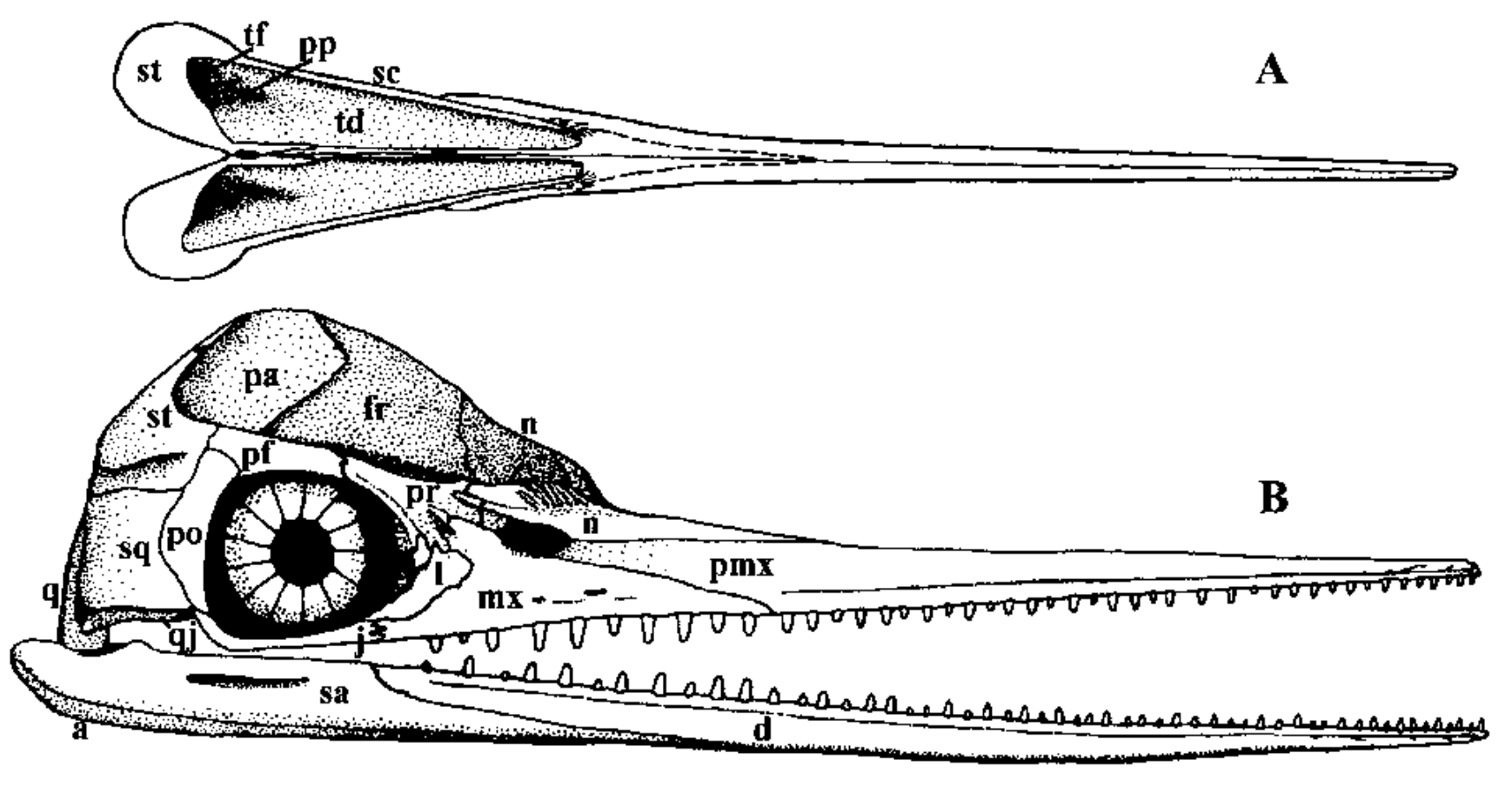
The euryapsid condition in Neusticosaurus
- A distinct supratemporal fenestra
- A very broad upper temporal arch separating the supratemporal fenestra from a vanishingly shallow, indistinct infratemporal embayment.
First, we describe the well-known groups whom everyone agrees to be monophyletic:
Eosauropterygia:
Eosauropterygia: (Early Triassic - terminal Cretaceous)
Min ∇ (Pachypleurosauria, Nothosauria, Plesiosauria)
These encompass an outstanding size range from the 20 cm. Triassic pachypleurosaurs to plesiosaurs the size of sperm-whales. Their general characteristics:
- Visually oriented predators, based on cranial endocasts Allemand et al., 2022.
- Limb-propelled swimmers with relatively stiff torsos
- Typically with necks elongated by the multiplication of cervical vertebrae
- It's been suggested that they had horizontal tail-flukes (Sennikov, 2019) or maybe vertical - Maybe.
- Premaxillae that extend far posteriorly between...
- Greatly reduced nasals
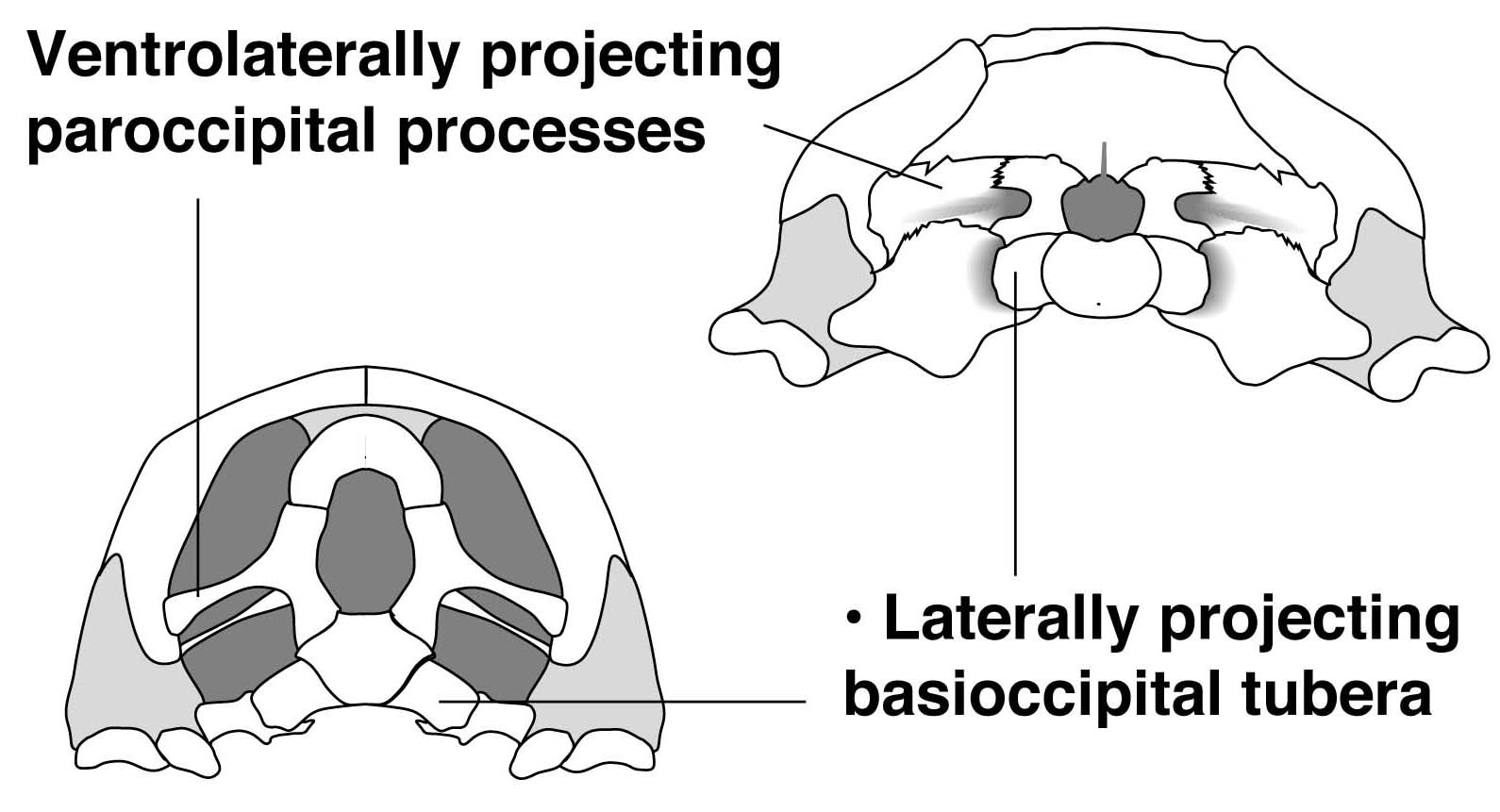
- Paroccipital processes that are directed ventrolaterally
- Derived palates in which the pterygoids meet on the midline, closing the interpterygoid vacuity and forming a ventral floor to the neurocranium (right) enclosing a distinct cranioquadrate canal.
- The suborbital fenestrae are secondarily closed
- The ectopterygoids are very long
- At least fifteen cervical vertebrae,
- Dorsal neural arches with accessory intervertebral articulations (dissimilar to those of squamates.)
- Transformed pectoral girdles in which the (anterior) coracoids meet in a broad symphysis that extends far posteriorly.
- Robust pubes and ischia that enclose a large thyroid fenestra.
Eosauropterygian diversity:
Pachypleurosauria: (Early - Middle Triassic) Small (20 - 100 cm) near-shore creatures of the Triassic Tethys Sea. Recent analyses either recover these as a paraphyletic grade-group of basal eosauropterygians (Shang et al., 2020, Wang et al., 2022) or as a monophyletic group (Xu et al., 2023; Hu et al., 2024). GORK, but the monophyletic hypothesis seems to have momentum.
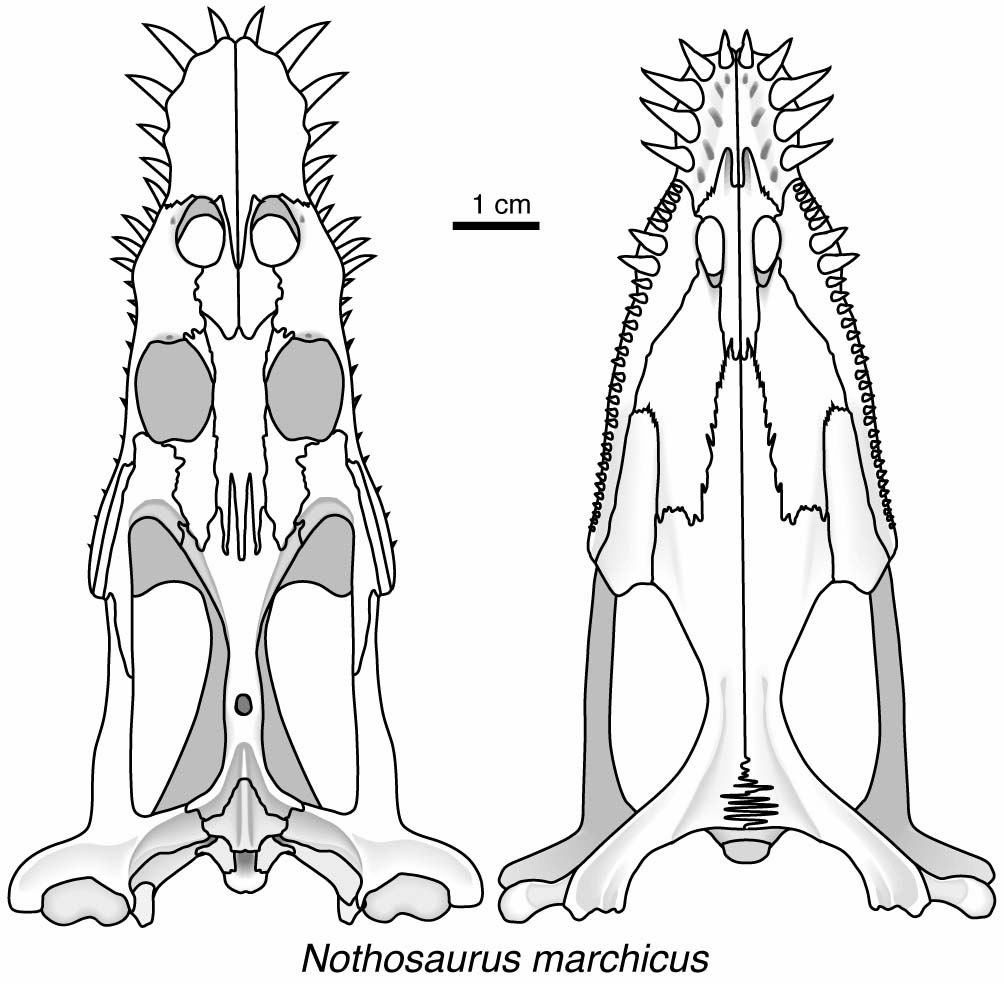
Inhabited shallow marine environments and shoreline. Ballast provided by pachyostotic ribs and vertebrae. Skulls characterized by:
- Very small supratemporal fenestrae
- Characteristic jaw articulation.
Could they walk on land? Probably. In many we see sexual dimorphism in which one gender has robust forelimbs and other has slender ones. Robust morph could be:
- Females coming ashore to lay eggs
- Males grappling onto females during mating
- The digestive system, including gut contents revealing that Keichousaurus was piscivorous, swallowing its prey whole (Li et al, 2023).
- Large embryos, suggesting live birth of young (Cheng et al, 2004).
"Nothosauria": (Middle - Late Triassic) Monophyletic or paraphyletic? See Wang et al., 2022. Small to large predators, inhabiting near-shore waters and shorelines. Their skulls display adaptations to the capture of large prey, captured by movement of the entire head on the end of the neck (head hunter). As in other predators of this sort:

- the temporal region is elongated
- the tooth row extends far posteriorly
Derived features include:
- fusion of the parietals
- extreme narrowing of the skull table
- reinforcement of the braincase in the temporal region by the widening of the epipterygoid. (Compare with alternate strategies in Aistopoda and Serpentes.)
- Occipital elements (supratemporal, opisthotic) strongly sutured to adjacent skull roof.
Current nothosaurian poster-child: Brevicaudosaurus jiyangshanensis with its improbably short fat tail.

M'mmm! Mud! Paludidraco multidentatus by N. Tamura does its suspension/deposit-feeding thing
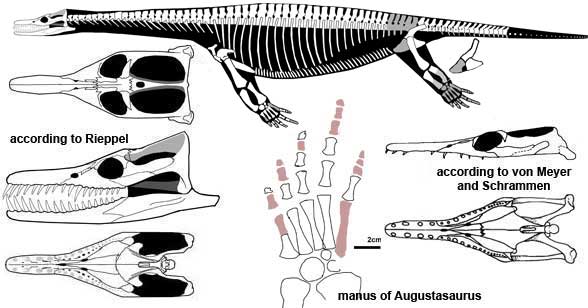
Pistosauroidea: (Early Triassic - terminal Cretaceous)
Max ∇ (Plesiosauria ~ Nothosauria, Pachypleurosauria)
Pistosauroids, including the classic plesiosaurs were the only Eosauropterygians to invade the open ocean, but they did it in stages: Earliest known members of the plesiosaur lineage were nothosaur-like in general form, but by the Middle Triassic, forms like Pistosaurus were finally invading the open oceans.
- Neck elongation achieved by increase in number of vertebrae (Pistosaurus had 24).
- Seal-like flippers retain distinct elbow, wrist, and fingers (enclosed in mitten of flesh). Maybe it could have come onto the beach.
Osteological trends: Among pistosauroids, we see:
- Extreme reduction of the nasals
- Nares retracted to being closer to orbit than to tip os snout.
- Braincase exposed ventrally and not concealed by pterygoids. (Contrast to Nothosauria.)
- In occipital view, the braincase is loosely connected to the skull roof (in contrast to Nothosauria.)
Plesiosauria: (Latest Triassic - terminal Cretaceous)
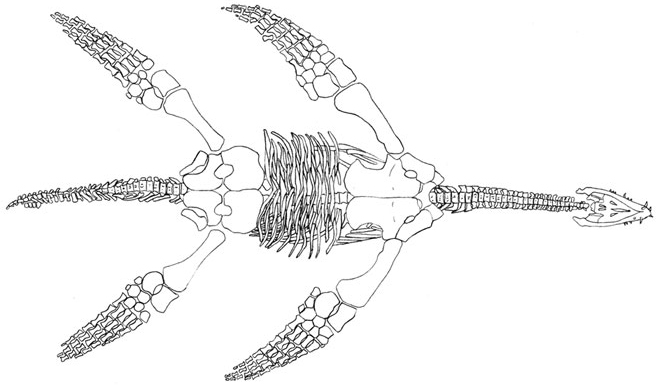
Max ∇ (Plesiosaurus ~ Augustasaurus)
The classic eosauropterygians of the mid Mesozoic. These experienced their primary radiation at the beginning of the Jurassic, and represent the second amniote radiation of the open oceans. Characterized by:
- limbs fully transformed into flippers (no functional elbow or wrist)
- compact torsos incapable of side to side undulation
- medium to large size
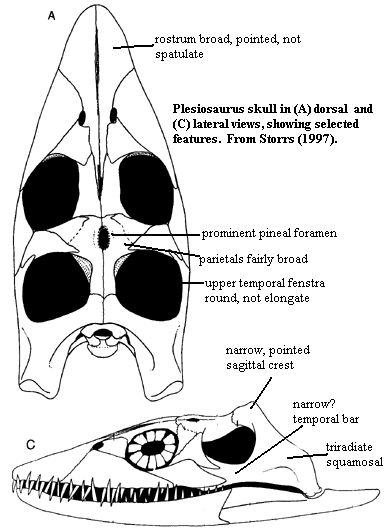
Synapomorphies include (Rieppel, 1997):
- Loss of nasals (E.G. Rhomaleosaurus)
- Pectoral and pelvic girdles with ventral elements greatly elongated.
- Pubis does not contact ilium
- Hyperphalangy
- Extremely robust gastralia
The popular image of plesiosaurs is of the derived long-necked, small-headed forms, however the ancestral state of body proportions is more like in the stem pistosauroid Pistosaurus, with a moderately long head on a moderately long neck (E.G. Rhomaleosaurus - right). From this beginning, plesiosaurs diversified into forms that were:

- very long-necked
- short necked with large heads - the "pliosaur" morph.
Plesiosaur diversity:
Rhomaleosauridae: (Jurassic) Relatively unspecialized with robust skulls. Synapomorphies include:
- Minimal contact bewtwen premaxillae and nares.
Neoplesiosauria: (Jurassic - Cretaceous)
Min ∇ (Plesiosaurus, Pliosaurus)
All other plesiosaurs, including Pliosauridae and Plesiosauroidea. These encompass short-necked and long-necked forms respectively:
Pliosauridae: (Jurassic - Cretaceous)
Max ∇ (Pliosaurus ~ Plesiosaurus)
Short-necked forms with large heads, including the largest plesiosaurs, with skulls up to 3 m. (E.G. Pliosaurus kevani). Note, though, that the earliest known plesiosaur - Rhaeticosaurus martens from the Late Triassic:
- is a basal pliosaur
- has a moderately long neck (Wintrich et al., 2017)
Plesiosauroidea: (Jurassic - Cretaceous)
Max ∇ (Plesiosaurus ~ Pliosaurus)
Long-necked forms with small heads. Among them, Plesiosaurus itself is relatively primitive. Osteological trends include:
- Elongation of the neck with over 28 cervicals
- Forelimbs larger than hindlimbs
- Premaxillae extend posteriorly separating prefrontals
Plesiosauroid include superlatives such as:
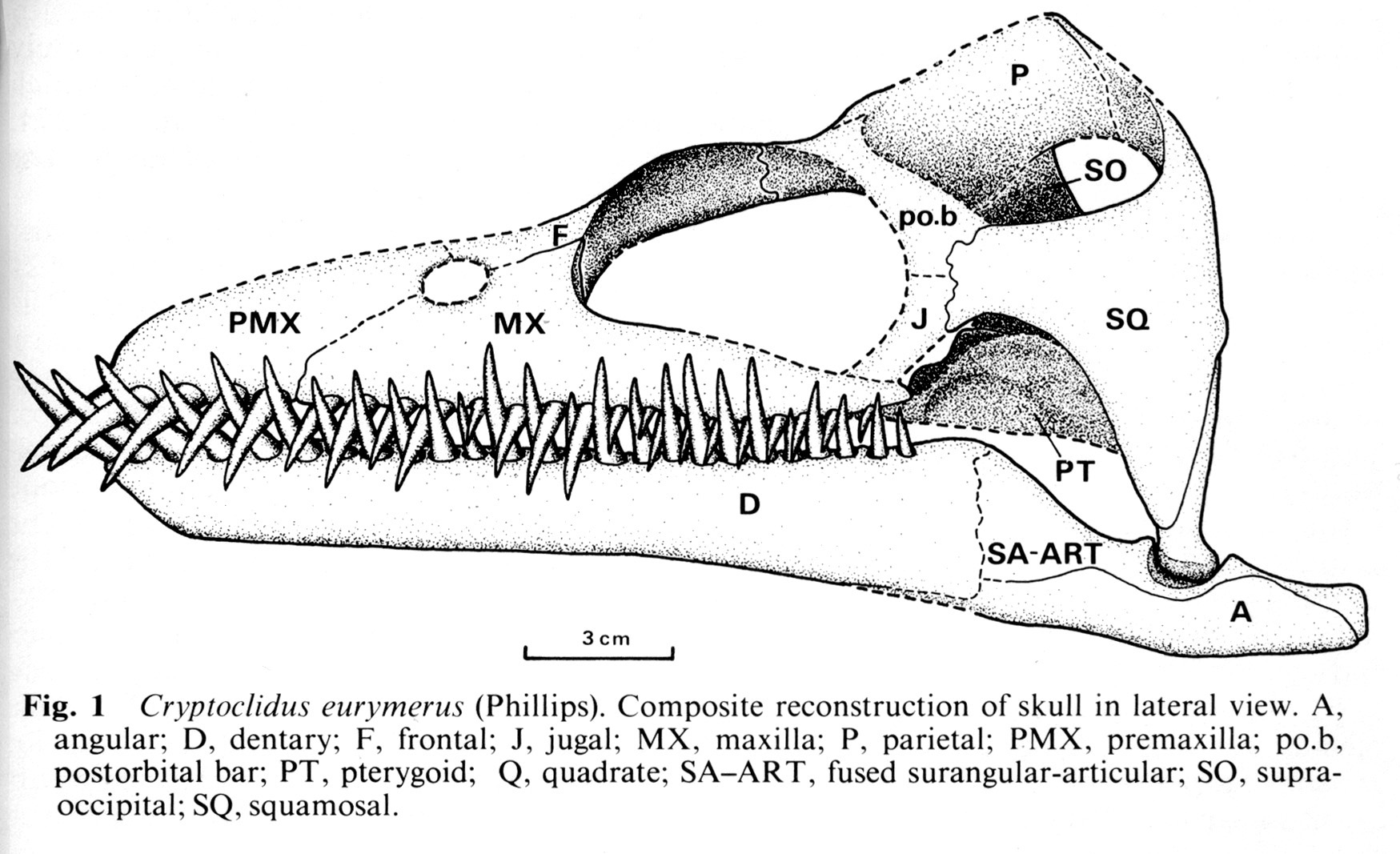
- Cryptoclididae: (Jurassic - Cretaceous(?)) With moderately long necks. Skulls display:
- Rod-shaped jugal
- Deep infratemporal embayment
- Elasmosauridae: (Cretaceous) who took the elongation of the neck to the greatest extreme (E.G. 72 cervical vertebrae in Elasmosaurus)
- Leptoclididae: (Early Cretaceous) Small with short-moderately long necks and small heads (like Brancasaurus), notable for tendency to invade brackish and fresh-water habitats. (Bunker et al., 2022)
- Polycotylidae: (Cretaceous) An independent evolution of the "pliosaur morph" among medium-sized Cretaceous plesiosauroids. (E.G. Dolichorhynchops). Interesting in part because of documented viviparity (O'Keefe and Chiappe, 2011). In this case, however, a single large fetus was observed.
Plesiosaur Ecology: Long-necked plesiosauroids seem to have exploited a hybrid predation strategy combining elements of pursuit and ambush predation, however there were variations. O'Keefe et al., 2017 describe the elasmosaurid Morturneria saymourensis as a suspension or deposit feeder, on the basis of its dentition.
Endothermy: Criteria of bone histology similar to those applied to terrestrial sauropsids, suggest that plesiosaurs may have had elevated body temperature and metabolic rate:
- Krahl et al., 2013 found fibrolamellar bone (more typical of endotherms) to be present in Pistosaurus but absent in Nothosaurus, suggesting that the shift from near-shore to pelagic ecology was accompanied by a metabolic shift.
- Fleischle et al., 2018 builds on this result with a survey of plesiosaur taxa, revealing bone histology indicative of endothermy and rapid growth throughout the group.
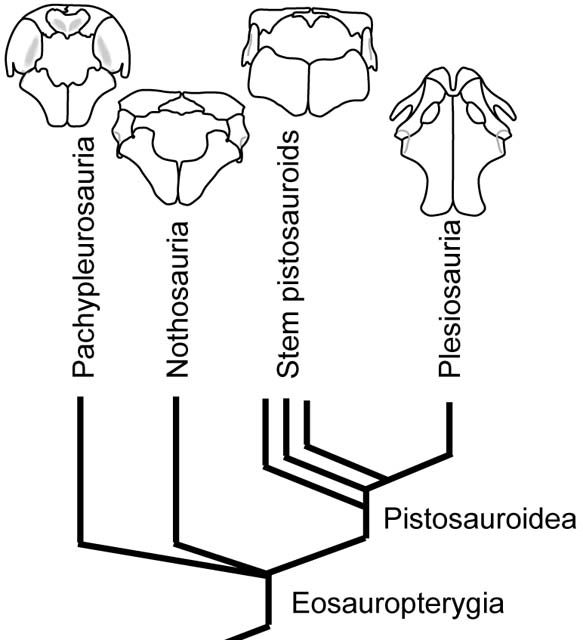
Eosauropterygian Phylogeny: But we skipped a step! Eosauropterygia consists of:
- Pachypleurosauria (Early - Middle Triassic) Small shallow marine predators.
- Nothosauria (Middle - Late Triassic) Medium to large shallow marine predators.
- Pistosauroidea (Early Triassic - Late Cretaceous) Medium to large marine predators that invade the open ocean.
- A handful of basal eosauropterygians to be addressed later.
Currently there is no consensus, but historically two hypotheses emerge:
- The Eusauropterygia hypothesis, where Nothosauria and Pistosauroidea form a clade, to the exclusion of the pachypleurosaurs. (E.G. Storrs, 1991; Ripple et al., 2002)
- The Basal Pistosauroidea hypothesis, where Nothosauria and Pachypleurosauria form a clade, to the exclusion of Pistosauroidea. (E.G. Merck, 1997; Hu et al., 2024).

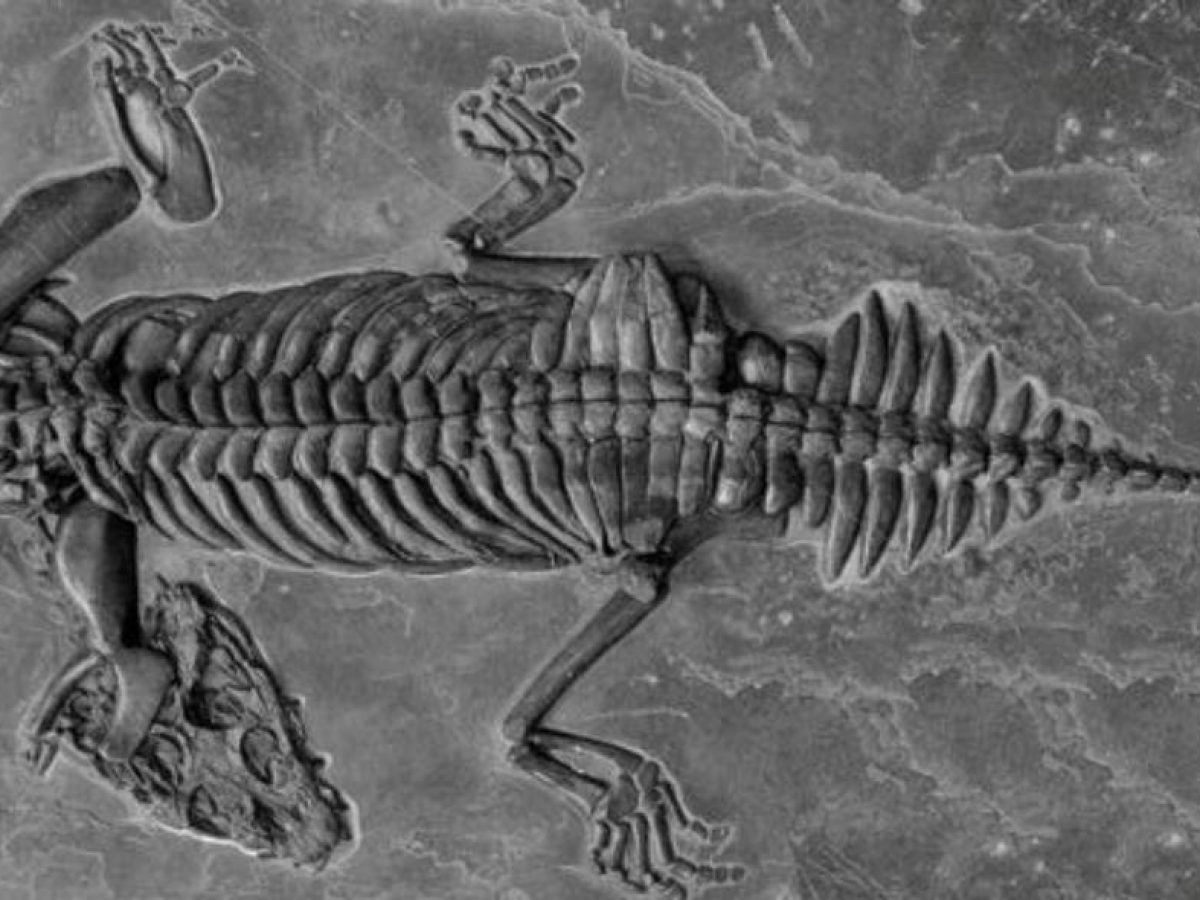
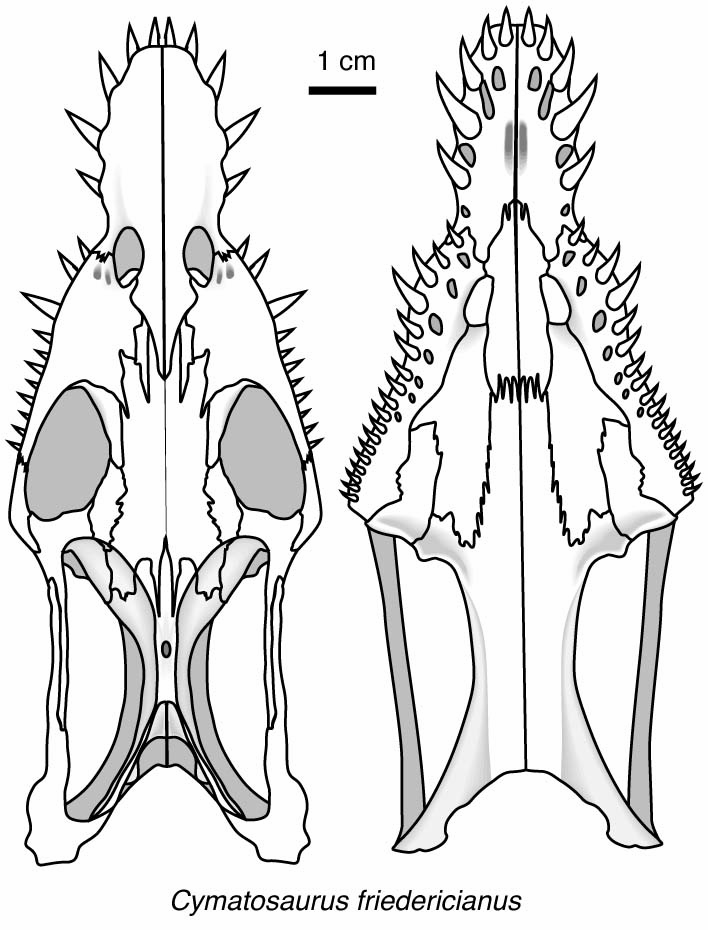
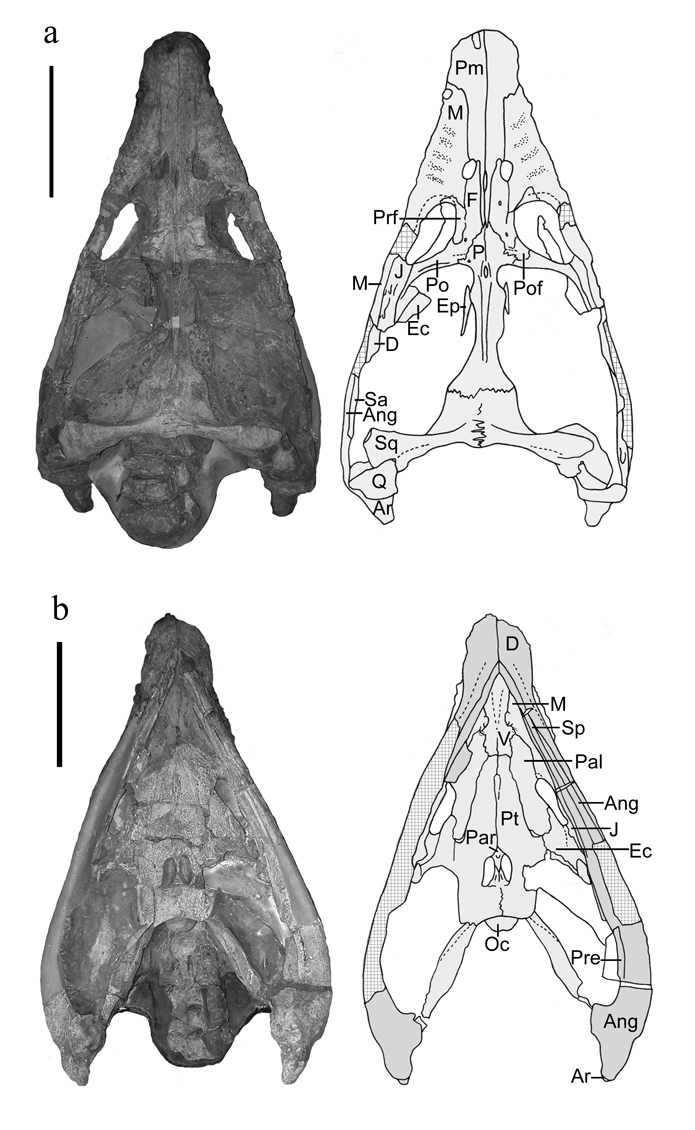
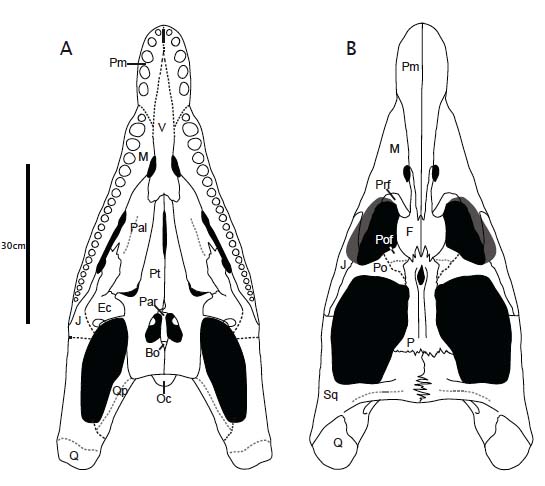
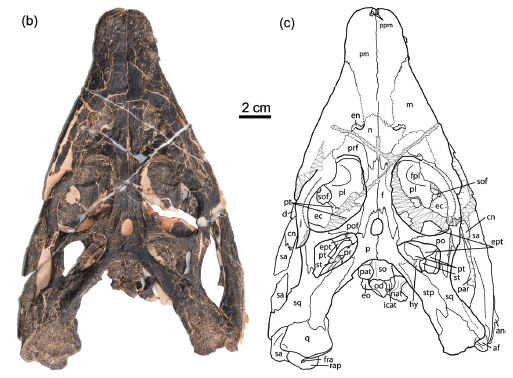
Placodontia:
(Middle - Late Triassic)
Placodontiformes: Max ∇ (Paraplacodus ~ Eosauropterygia)
Placodontia: Min ∇ (Paraplacodus, Psephoderma)
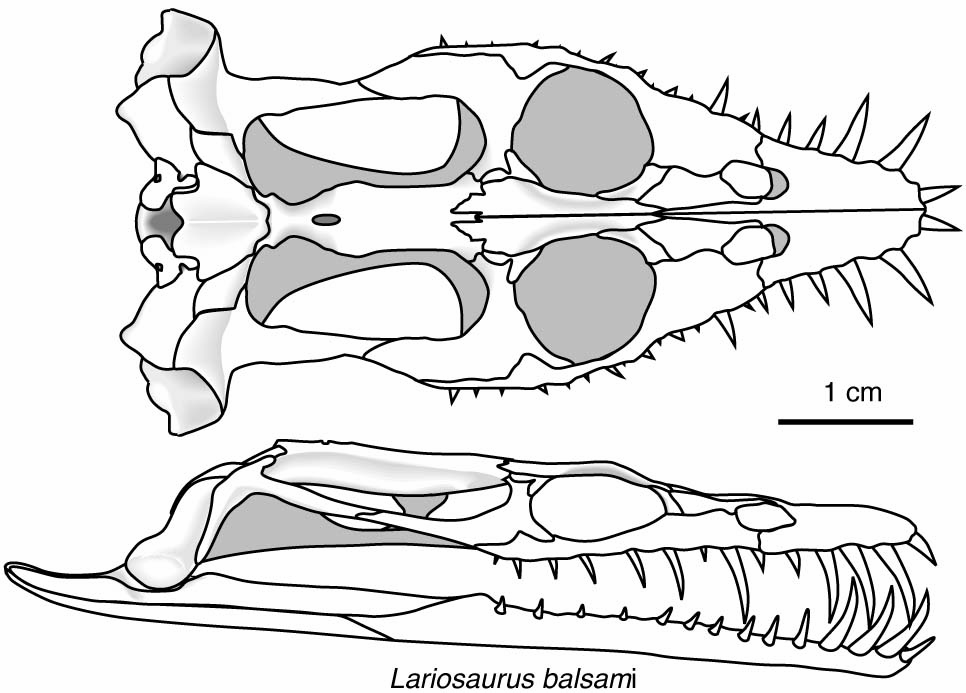
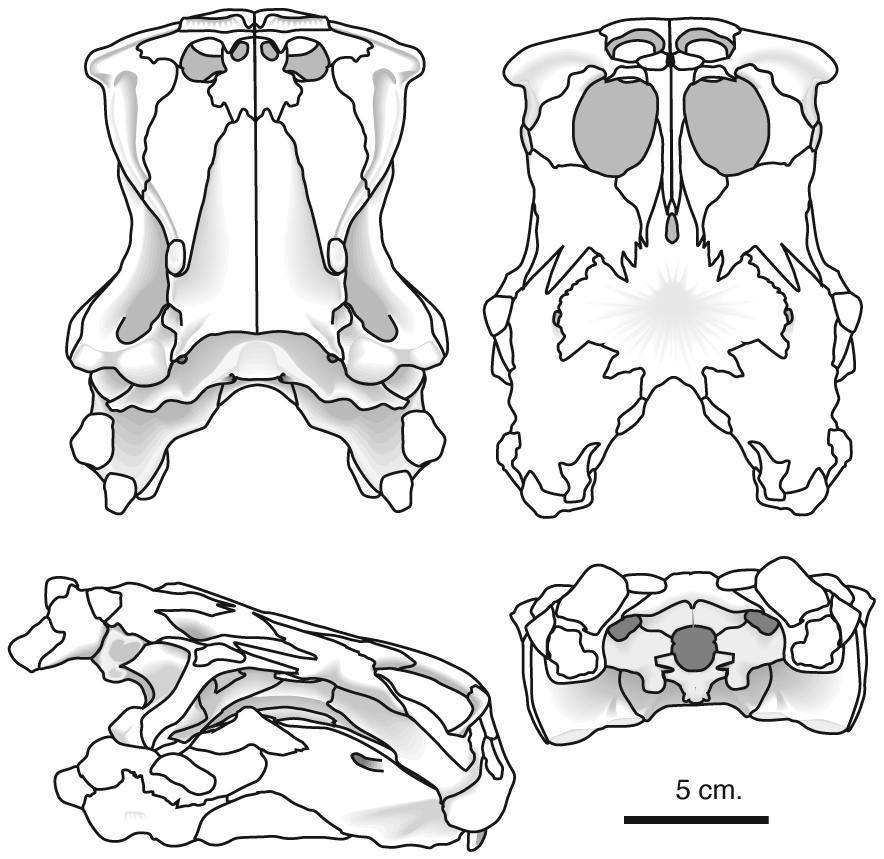
Includes Placodus pictured here was restricted to shallow marine environments of the middle and late Triassic. They probably swam clumsily or walked on the bottom in the manner of a snapping turtle. In contrast to their locomotor apparatus, their skulls were intensely modified for withstanding terrific biting forces. The dorsal view at right shows the great size of the muscles mass that closed the jaw. The palatal view shows the transformation of the teeth (marginal and palatal) into a pavement of button-like teeth with which they presumably crushed hard-bodied invertebrates.
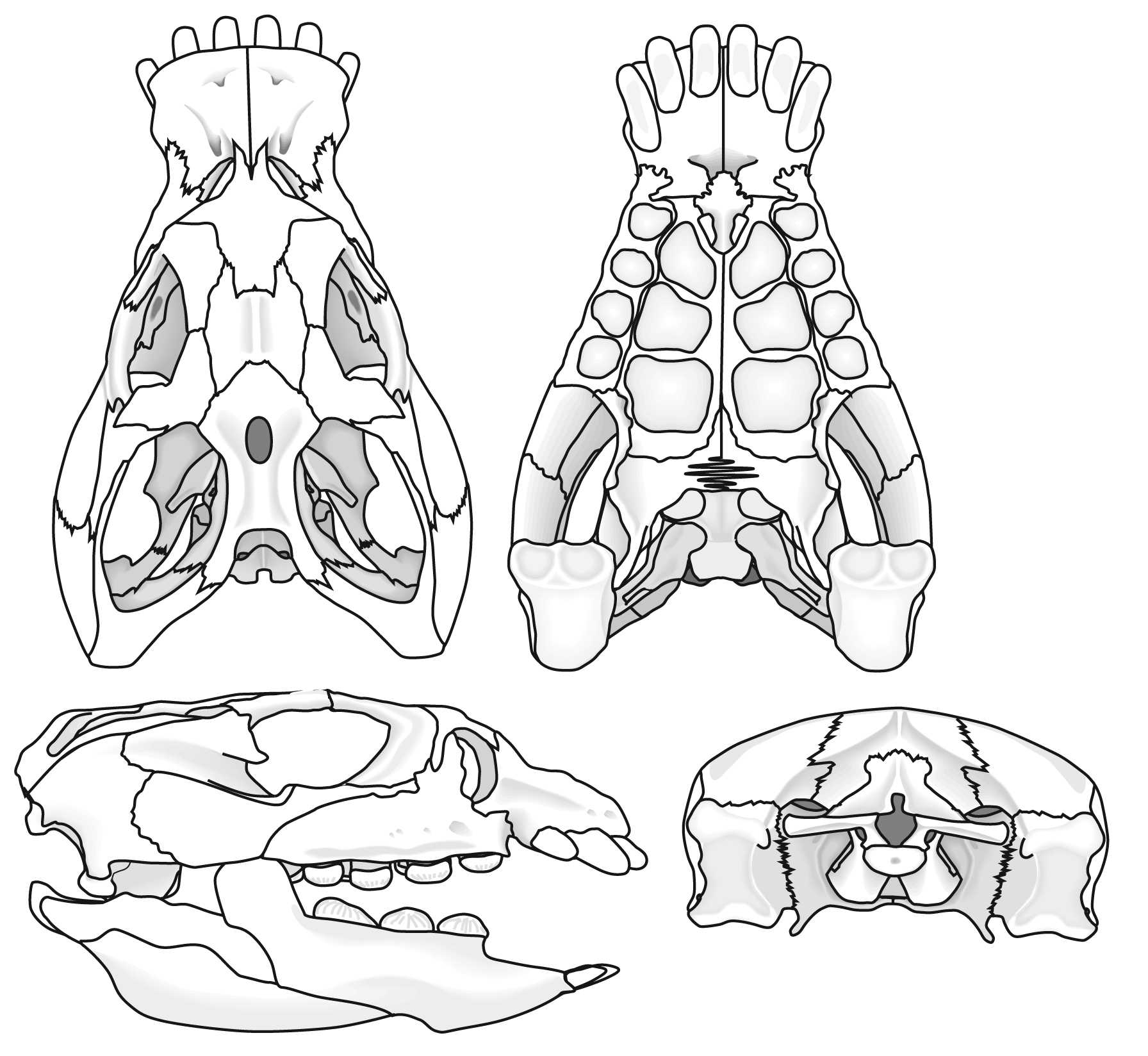
Placodus gigas
- The presence of button-shaped marginal and palatal teeth
- Differentiation between premaxillary and maxillary teeth
- Pterygoids that meet, obliterating the interpterygoid vacuity and forming a floor to the braincase.
- Significant reinforcement of the occipital region by suturing of elements
- Presence of an anterior element of the braincase similar to the laterosphenoid of snakes
- Pachostotic ribs and gastralia give trunk a boxy look.
More derived placodonts form the Cyamodontoidea Many developed extensive dermal armor reminiscent of that of turtles. At least one member, Henodus chelyops has been recovered from lagoonal/estuarine environments. How it fed is enigmatic, but it does not seem to have been crushing hard shelled prey as the others did. Grazing and suspension feeding have been proposed.

At the other end, the stem placodont, Palatodonta bleekeri (Middle Triassic), lacks crushing teeth altogether but shares procumbent incisors with placodus. (Neenan et al., 2013)

Ichthyopterygia:
(Early Triassic - Early Late Cretaceous)
Included many ecologically shark or dolphin-like pelagic predators. Ichthyosaurs and their close relatives were the first marine reptiles, appearing in the Early Triassic. Initially near-shore creatures, Late Triassic ichthyosaurs became the first marine reptiles to invade the open oceans. Remarkably, Kear et al., 2022 report derived ichthyosaur remains from only 2 my after the End Permian Extinction Event and conclude that ichthyosaurs must have been present in the Late Permian. This extraordinary claim requires verification.
Ichthyosaurs appear in the fossil record highly modified for marine life, with:
- limbs modified as flippers
- a distinct tail bend supporting a caudal fin
- greatly enlarged eyes
- no obvious means of coming ashore to lay eggs. For decades we have known that in derived forms, ichthyosaurs clearly gave live birth. Recently it has come to light that the very primitive ichthyosaur Chaohusaurus also did so.
Chaohusaurus: (Early Triassic) Among the first ichthyosaurs and phylogenetically most primitive. Roughly the size of an otter, it nevertheless was unable to come onto land, and shows the general synapomorphies of Ichthyopterygia, including::

- Consolidation of fore and hindlimb into flippers.
- A peak in height of the caudal neural spines with spines on either side inclined toward one another (the precursor of a proper tail-bend)
Grippioidea (Early Triassic) Represented by the well-known Utatsusaurus hataii shows the general synapomorphies of Ichthyopterygia, including:
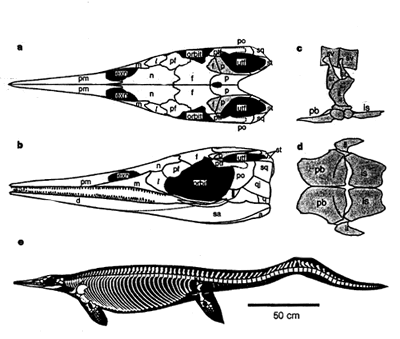
- Postfrontal with posterior process
- Anterior terrace of supratemporal fenestra. This seems to allow the jaw adductor muscles to extend onto the skull roof, strengthen the bite of these otherwise very gracile heads.
- Closure of interpterygoid vacuity
- Loss of ectopterygoid
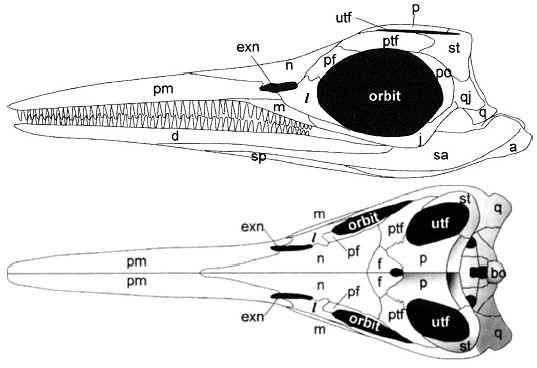
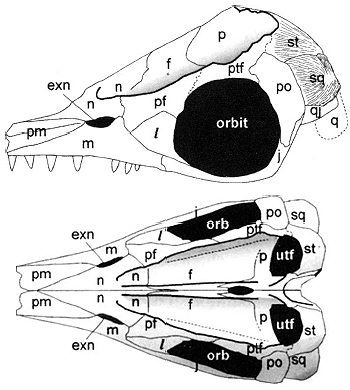
Ichthyopterygian irritant: The interpretation of the temporal region as having a large three-dimensional supratemporal occupying the posterior border of the supra temporal fenestra, and posterolateral corner of the skull, and a small plate-like squamosal on the side of the skull is universally accepted by ichthyosaur specialists. This creates the odd situation where:
- the supratemporal looks like a normal squamosal
- the squamosal looks sort of like a normal quadratojugal.
Ecology of basal ichthyopterygians: Despite their long snouts, Chaohusaurus and Utatsusaurus don't seem to have been simple "snout-hunters," as their posterior teeth were bulbous, as if for crushing hard prey. Did they, perhaps, probe into soft sediment for shelled prey then crush it in the backs of their jaws?
Ichthyosauria: (Middle Triassic - Early Late Cretaceous)
The synapomorphies:
- Lateral orientation of the naris
- Exclusion of the postorbital from the supratemporal fenestra
- Prefrontals and postfrontals meet, excluding frontal from orbit margin
- Pineal foramen on the fronto-parietal suture.
- Ulna lacks a post-axial shaft (i.e. perichondral ossification does not occur there.)
Cymbospondylidae: (Middle - Late Triassic) Medium to large animals. From such beginnings we get, by the Late Triassic, s whale-sized ichthyosaurs like Cymbospondylus. This was, nevertheless, an elongate eel-shaped creature. In Cymbospondylidae, we finally see ichthyopterygians with uniformly sharp pointed teeth.
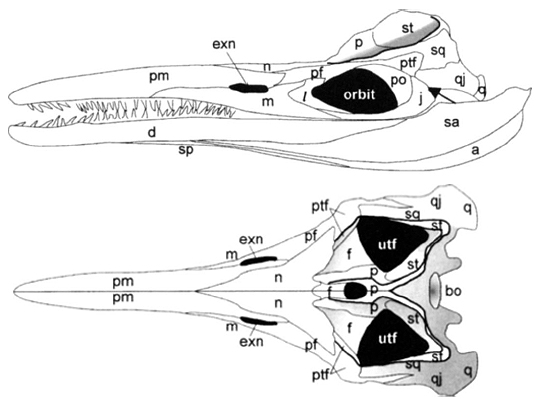
The image of Cymbospondylus at right allows us to track a trend in ichthyosaur evolution: The nasals become increasingly large, overlapping adjacent elements like the frontals. This parallels other long snouted marine amniotes, who use overlap of adjacent dermal elements to strengthen the region between the orbits - a zone of weakness. Parallel developments include:
- Enlargement of the frontals in mosasaurs
- Enlargement of maxillae and premaxillae in toothed whales

PC1 and PC2 of morphometric comparison of marine reptiles by Gutarra et al., 2023.
Ichthyopterygians in red, mosasaurs in salmon.
If, at this point, you feel like you have heard the story of ichthyopterygian evolution before, only in a squamate context, you are not alone. Until this point, the evolutionary trajectory of Triassic ichthyopterygians toward life as open ocean predators has largely prefigured that of Cretaceous mosasaurs in features like:
- Evolution of hydropedal limbs and hydropelvic pelves
- Elongation of the body
- Appearance of a tail-bend and caudal fin
- Elongation of the rostrum
But all of that is about to change, as ichthyosaurs adopt ecological strategies unmatched by mosasaurs.
Hueneosauria: (Middle Triassic - Early Late Cretaceous)
Min ∇ (Mixosaurus, Parvipelvia)
Named for early 20th century ichthyosaur luminary Friedrich von Huene.
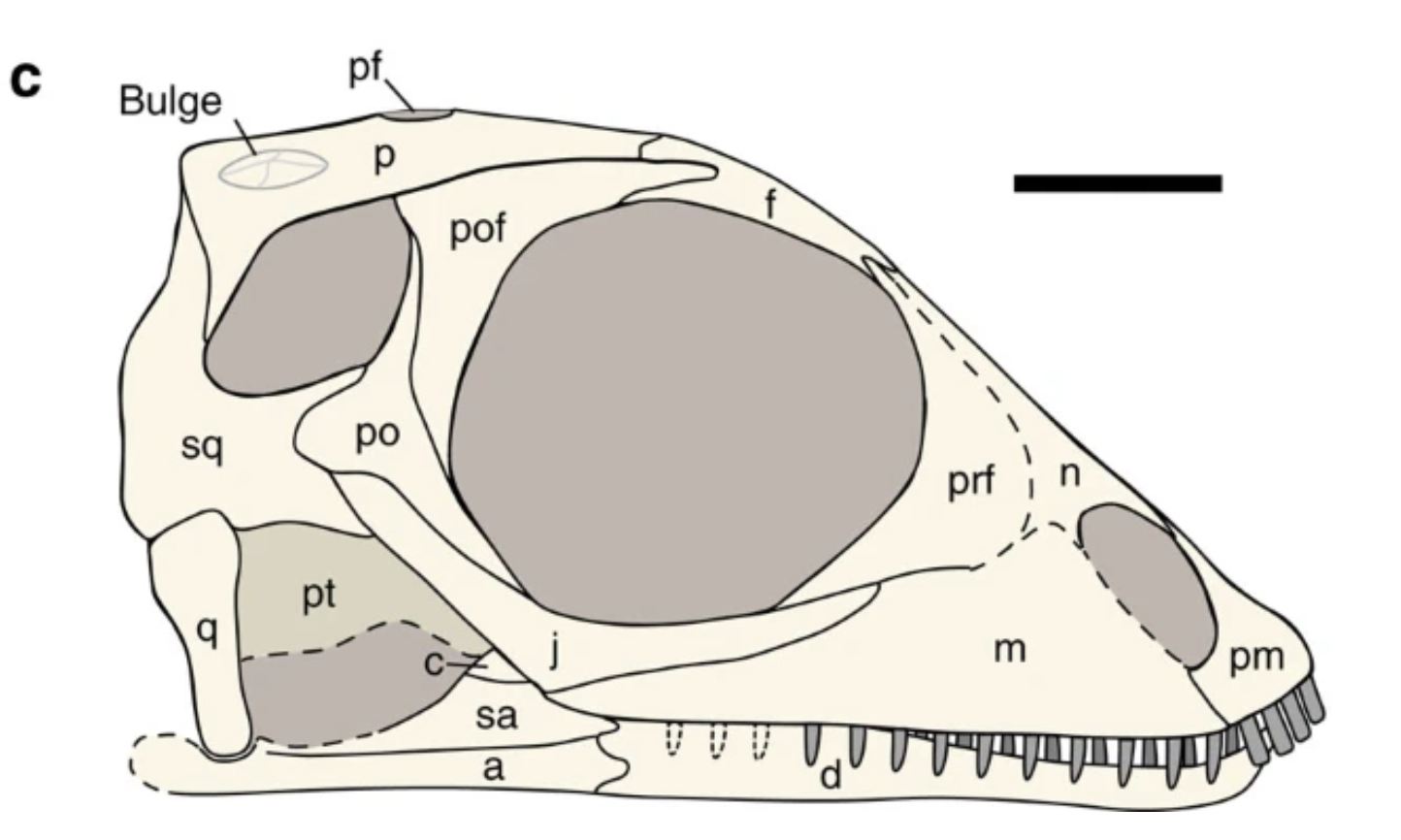
Mixosauridae: (Middle Triassic) small ichthyosaurs representing the first appearance of a more compact body form and distinctly larger forelimbs. A strange autapomorphy: the anterior terrace of the supratemporal fenestra extends far anteriorly past the orbits. Some superlatives:
- Mixosaurus preserves the oldest evidence for a proper dorsal fin. (Renesto et al., 2020)
- Contectopalatus is exceptionally large for a mixosaurid, with individuals possibly up to 5 m in length. Its skull shows an especially robust form of the supra temporal anterior terrace which, coupled with a tall sagittal crest, provides extensive origin points for jaw muscles. (Maisch and Matzke, 2000)
"Shastasaurs": (Late Triassic). The Late Triassic saw the radiation of ichthyosaurs that clearly invaded the open ocean realm - the first amniotes to do so unambiguously. This "shastasaurid" grade included the largest marine reptiles on record and possibly the largest animals ever. (See Shonisaurus sikanniensis from Nicholls and Manabe, 2004.)


Min ∇ (Temnodontosaurus, Ichthyosaurus)
More primitive ichthyosaurs were extinguished by the Triassic/Jurassic extinction event, however ichthyosaurs roared back in the early Jurassic in the form of Parviplevia, the ichthyosaurs with reduced pelves and hindlimbs. This is also the time of the rise of plesiosaurs. Plesiosauria and Parvipelvia have in common their invasion of the open oceans. This is also the interval in which we have soft-tissue impressions that reveal the dorsal and caudal fins of these animals. (Note: a few of these seem completely genuine but many were "improved" by 19th century preparators.)
Of special interest:
- Diet: Most preyed on small small items, although some are thought to have specialized on shelled prey like cephalopods while others specialized in softer prey. (Jamison-Todd et al., 2022) Some, like Temnodontosaurus were orca-sized macropredators.
- Elevated metabolism: The body profile of parvipelvians reveals dorsal fins and tall semilunate (half-moon shaped) tails with narrow caudal peduncles. Among living taxa these characterize active fast-swimming pursuit predators like dolphins, tuna, billfish, and mako sharks. Soft tissue outlines also indicate the presence of layers of blubber - thermal insulation integument. Indications of tachymetabolic (warm-blooded) metabolism are found in:
- Fibrolamellar bone histology typical of warm-blooded animals. Surprise! Even basal ichthyopterygians like Utatsusaurus have this bone type.
- Oxygen isotope ratios in hard tissue consistent with significantly elevated body temperatures. (Bernard et al., 2010.
- Visual hunting: Parvipelvians include animals with the largest eyes of any vertebrate, both proportionally and in absolute size. Creatures like Ophthalmosaurus of the Late Jurassic represent the biomechanics limit for vision-driven hunters and are estimated to have been able to see at depths of 1.6 km. Oddly, Allemand et al., 2022 report that ichthyosaur endocasts suggest have surprisingly robust olfactory systems. How they used their sense of smell is enigmatic.
- Integument: Parvipelvian ichthyosaurs are famous for the preservation of body outlines in lagersätten like Holtzmaden (Early Jurassic) and Solnhofen (Late Jurassic). In exceptional cases, evidence of skin texture and pigment are preserved, indicating smooth skin, a layer of blubber, and countershaded coloration - dark above and lighter underneath. (Eriksson et al., 2022.)
Parvipelvian trends:
- Lacrimal contacts margin of nares (reversal to ancestral sauropod condition!)
- Large supratemporal extends forward to the postfrontal
- Nasals enlarge, strongly overlapping frontals and prefrontals
- Figure-eight shaped nares. In later forms like Ophthalmosaurus, the nares are completely divided, apparently to divide the functions of breathing and elimination of salt-gland brines. (Massare, 2021)
- Pineal foramen occupies border of parietals and frontals or is enclosed by frontals.
- The scapular blade is rectangular
- Pelvis significantly reduced with weak or absent sacrum
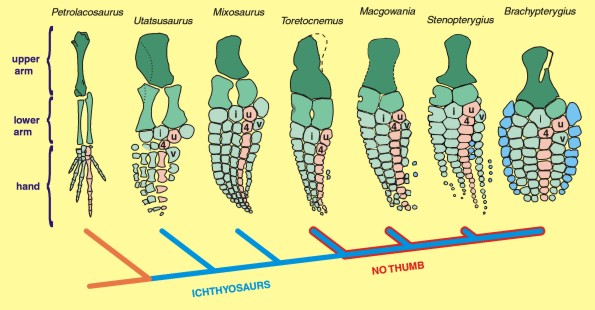
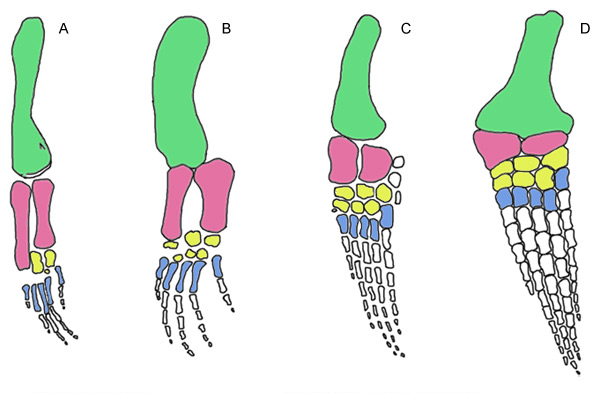
Limb elements: With eosauropterygians, the homology of limb elements, even in highly derived taxa, is reasonably clear. The only issue is hyperphalangy, the their tendency to add phalanges to each finger. With ichthyosaurs, however, the identity of limb elements is difficult to assess, in part, because of hyperdactyly, their tendency to add extra fingers, as well. No surprise that a volume on homology would use an ichthyosaur paddle in its cover design.

Edennasaurus acutirostris.
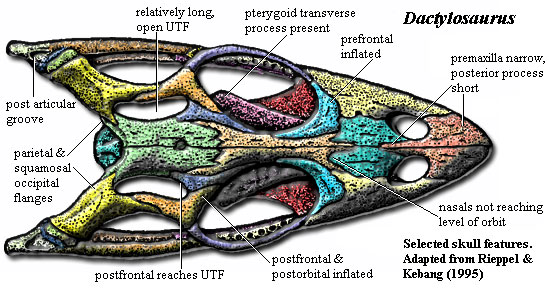
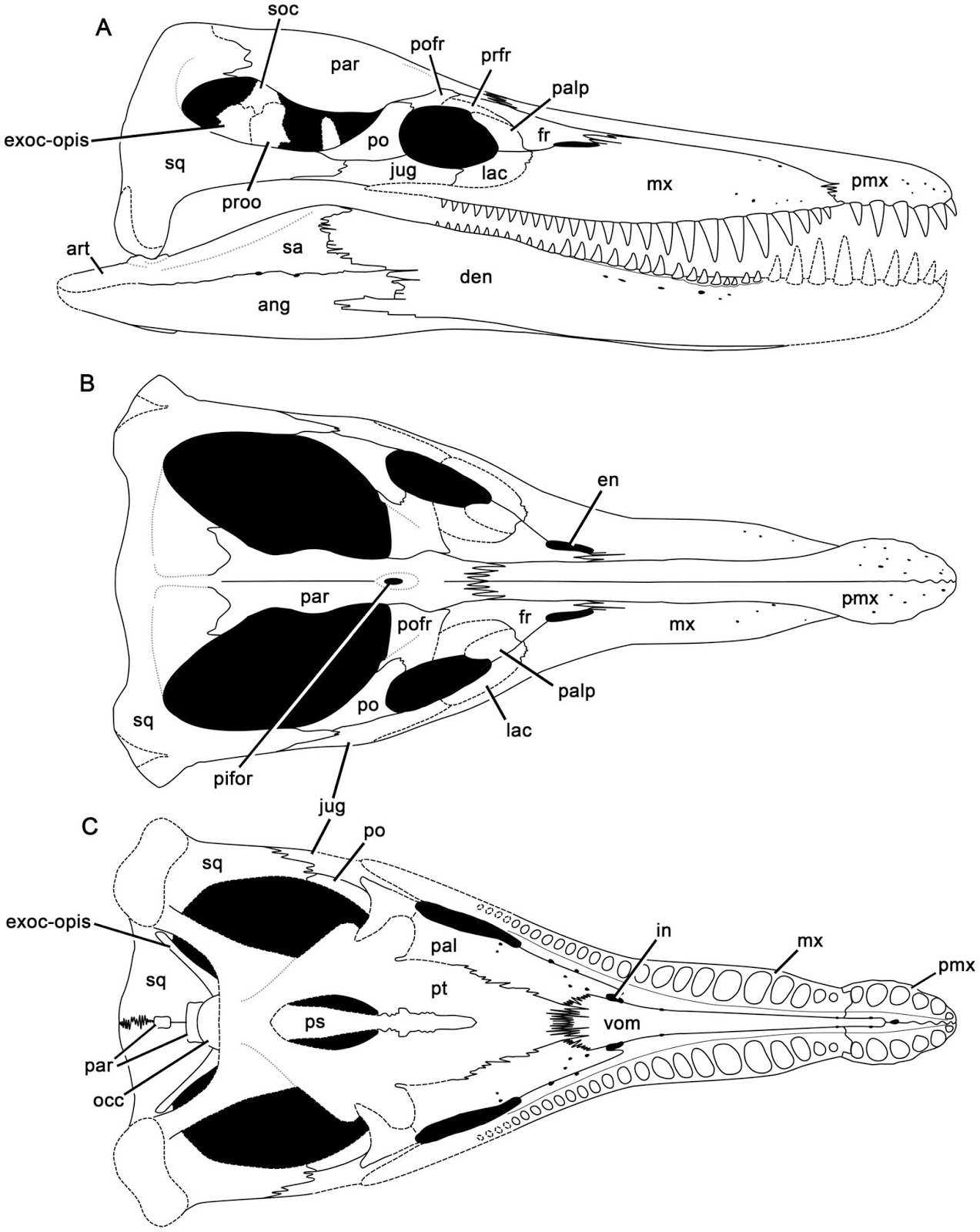
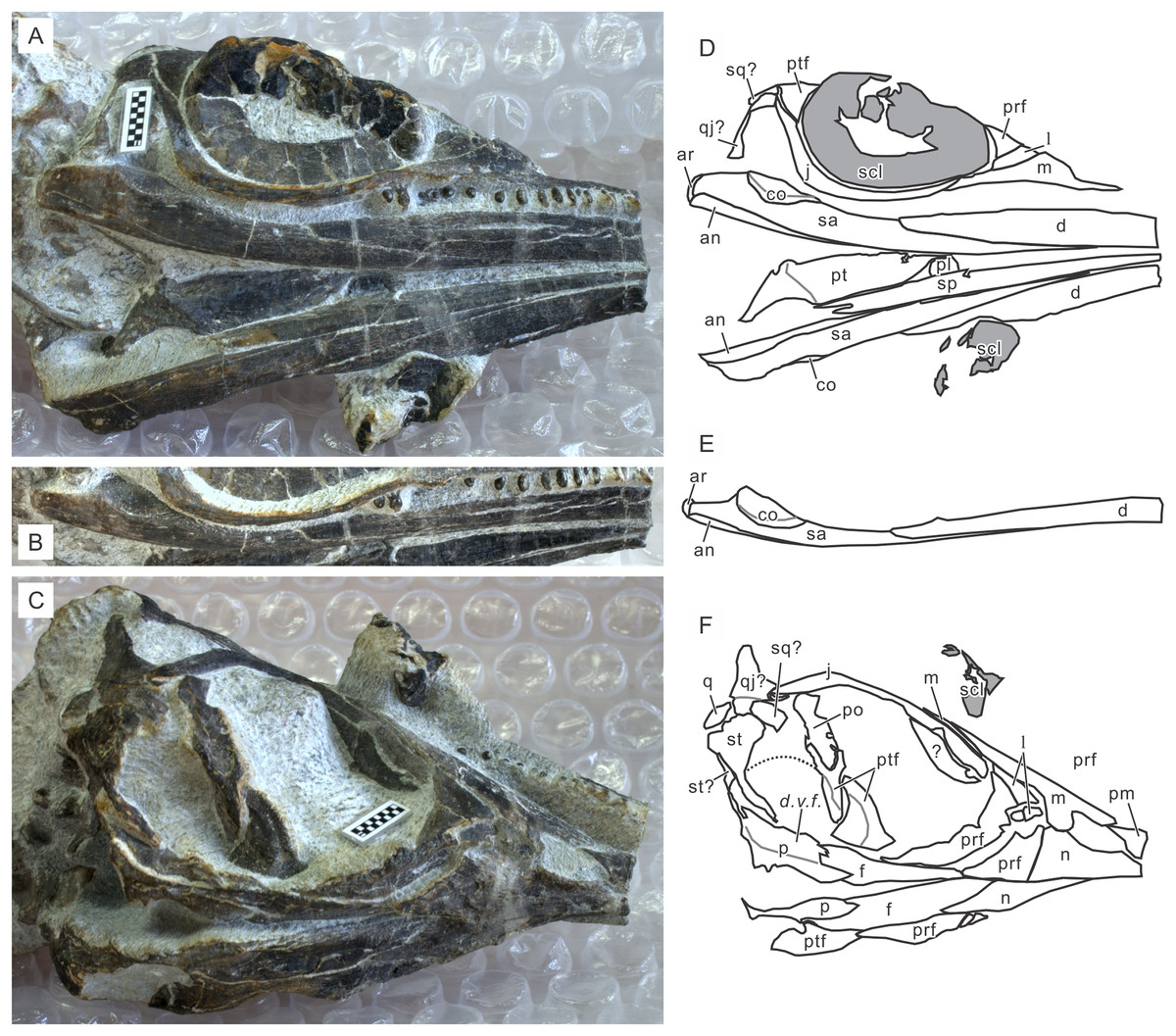
Thalattosauriformes/Thalattosauria
(Middle - Late Triassic)
Thalattosauriformes: Min ∇ (Thalattosaurus, Edennosaurus, Askeptosaurus)
Thalattosauria: Max ∇ (Nectosaurus, Hescheleria ~ Edennosaurus, Askeptosaurus)(Nicholls, 1999)
A final group of well-known marine saurians. Thalattosaurs were medium - medium-large animals capable of movement on land or in water. Edennasaurus (right), a more basal form is roughly the size and proportions of the Early Permian Mesosaurus (remember?) Ancestrally, however, thalattosaurs were neither head-hunters nor snout-hunters but a little of each. The thalattosaurian radiation encompassed larger predators like Askeptosaurus and durophagous forms like Thalattosaurus and Hescheleria.
Thalattosaurian synapomrphies:

- Elongate rostrum formed by premaxillae
- Posterior median process of premaxillae extends posteriorly past nares
- Nasals reduced
- Elongate posterior processes of jugals nearly close infratemporal embayment
- Supratemporal fenestrae reduced to narrow slits or closed
- Squamosals reduced
- Supratemporals large
- Distinct epiphyses of long limb elements.
Thalattosaur systematics: Are poorly resolved except for the presence of two major clades:
- Askeptosauroidea: Plesiomorphic skulls. Trunk skeleton emphasizes robust gastralia that limit undulations, indicating a greater reliance on appendicular swimming. (As in Edennasaurus, above)
- Thalattosauroidea: Plesiomorphic trunks. Skulls are variously modified for eating hard-shelled prey, with blunt teeth and (Thalattosaurus, right) down-turned premaxillae.
Indeed, what do these animals have to do with one another phylogenetically? Only one answer emerged clearly from the early days of cladistics:

Sauropterygia
(Early Triassic to Late Cretaceous) Storrs, 1991 clearly found Placodontia and Eosauropterygia to be sister taxa. Their synapomorphies include:- Closure of pterygoids beneath neurocranium
- The clavicles rest against the medial surfaces of the coracoids and scapulae.
Recent Arrivals:

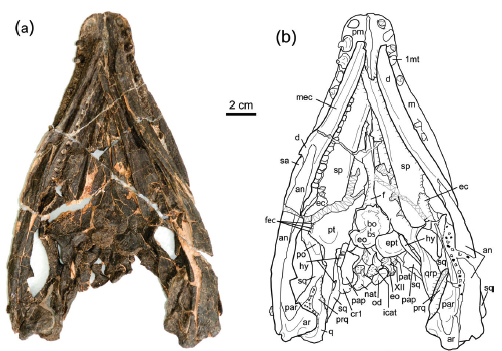
Saurosphargidae
(Middle Triassic) Burst onto the scene as well-described taxon only in the last decade. Characteristics include:- Elongate dorsal transverse processes
- Trunk skeleton in which transversely expanded ribs form a flattened "basket" covered with small osteoderms, forming a superficially (?) turtle-like shell, closed ventrally by robust lateral gastralia. (Note: this pattern differs in detail from that of armored placodonts or turtles.)
- Vaguely sauropterygian-like skulls in which the nares are retracted and the supratemporal fenestra is small or closed
- Palates in which the interpterygoid vacuity remains open.
- Pectoral girdles in which the clavicle is applied to the medial surface of the scapula and coracoid.
Points of interest:
- Middle Triassic saurosphargids tend to have flattened and laterally expanded trunk profiles in dorsal view, however the Early Triassic saurosphargiform Pomolispondylus baini is elongate. (Cheng et al., 2022.)
- Eusaurosphargis dalsassoi, which may (Wang et al., 2022) or may not (Scheyer et al., 2017) belong to Saurosphargidae, was apparently a terrestrial animal.

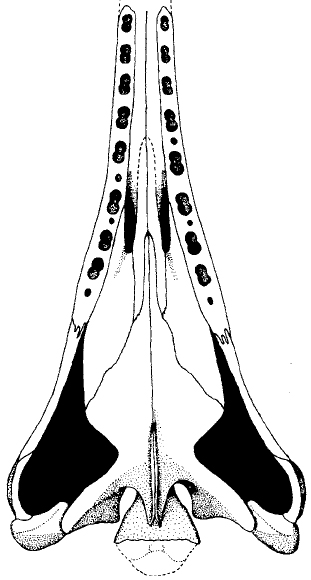
Atopodentatus unicus
Atopodentatus
(Early Triassic) New from Cheng at al., 2014, a 3 m. grazing herbivore (?):- Originally described based on badly deformed skull. Chun et al., 2016 illuminated true head shape as resembling floor sweeper with cropping teeth anteriorly and, possibly, suspension feeding teeth posteriorly.
- Emerges in most recent analyses as basal placodontiform. (E.g. Wolniewicz et al., 2022)
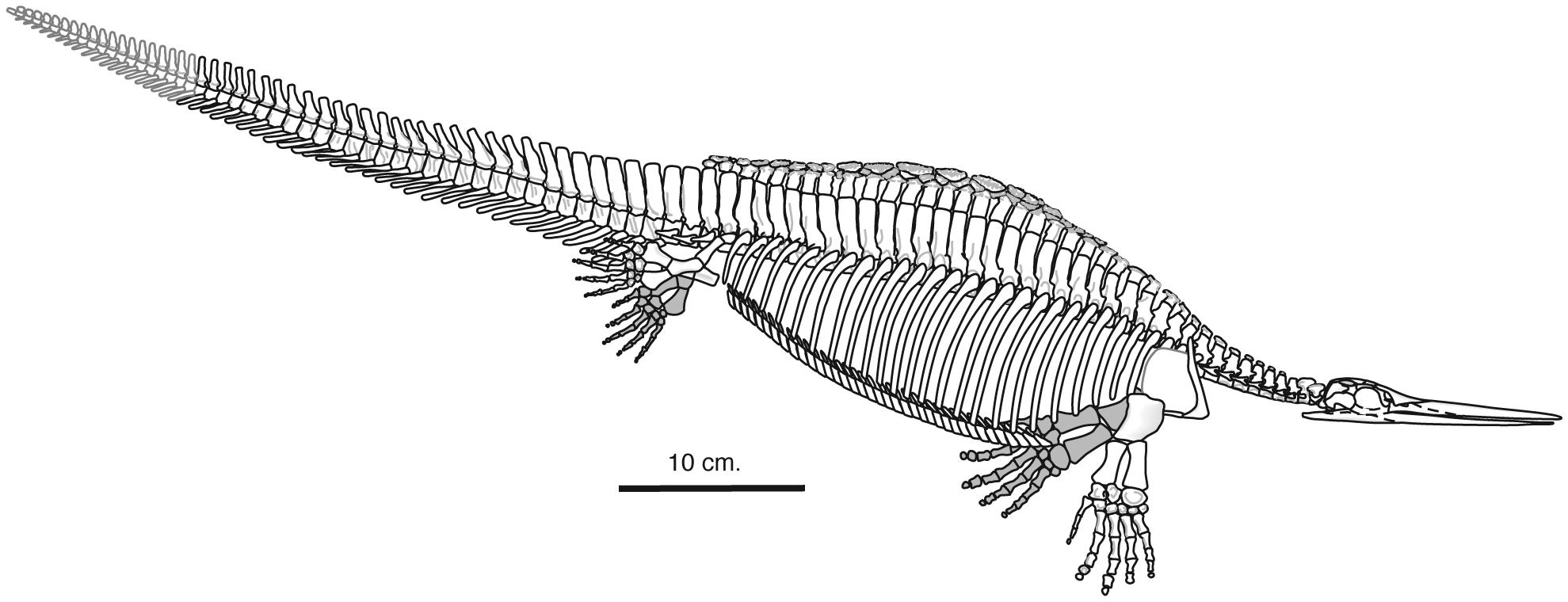
Hupehsuchia
(Early Triassic) A small group from the Early Triassic of China, a mash-up of ichthyosaurs in general proportions, eosauropterygians in stiffening of the trunk, and pladoconts in dermal armor. The first described was Hupehsuchus, impishly employed by Carroll and Dong, 1991 to demonstrate the perils of cladistic analysis. Hupehsuchians are characterized by:- spatula-shaped toothless rostrum
- trunk skeleton characterized by:
- Mid-line accessory elements articulating with neural spines
- Broad pachyostotic ribs articulating with
- Robust gastralia
- a tendency of the digital arch toward developing extra digits.
- A rostrum that is convergent on the morphology of baleen whales, strongly indicating lunge suspension-feeding as a strategy.
Eretmorhipis carrolldongi: (Early Triassic) If hupehsuchians aren't strange enough for you already, consider this animal with its:

- Five and a half fingers
- Expanded dorsal osteoderms
- Platypus-shaped skull
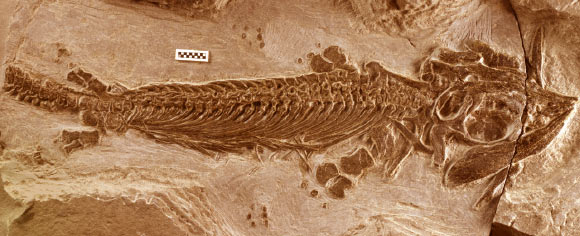
Cartorhynchus
From Motani et al., 2015, Cartorhynchus, a basal Early Triassic ichthyosauriform from southern China with:- a short snout invested with blunt durophagous teeth.
- flexible flippers with wrists and ankles
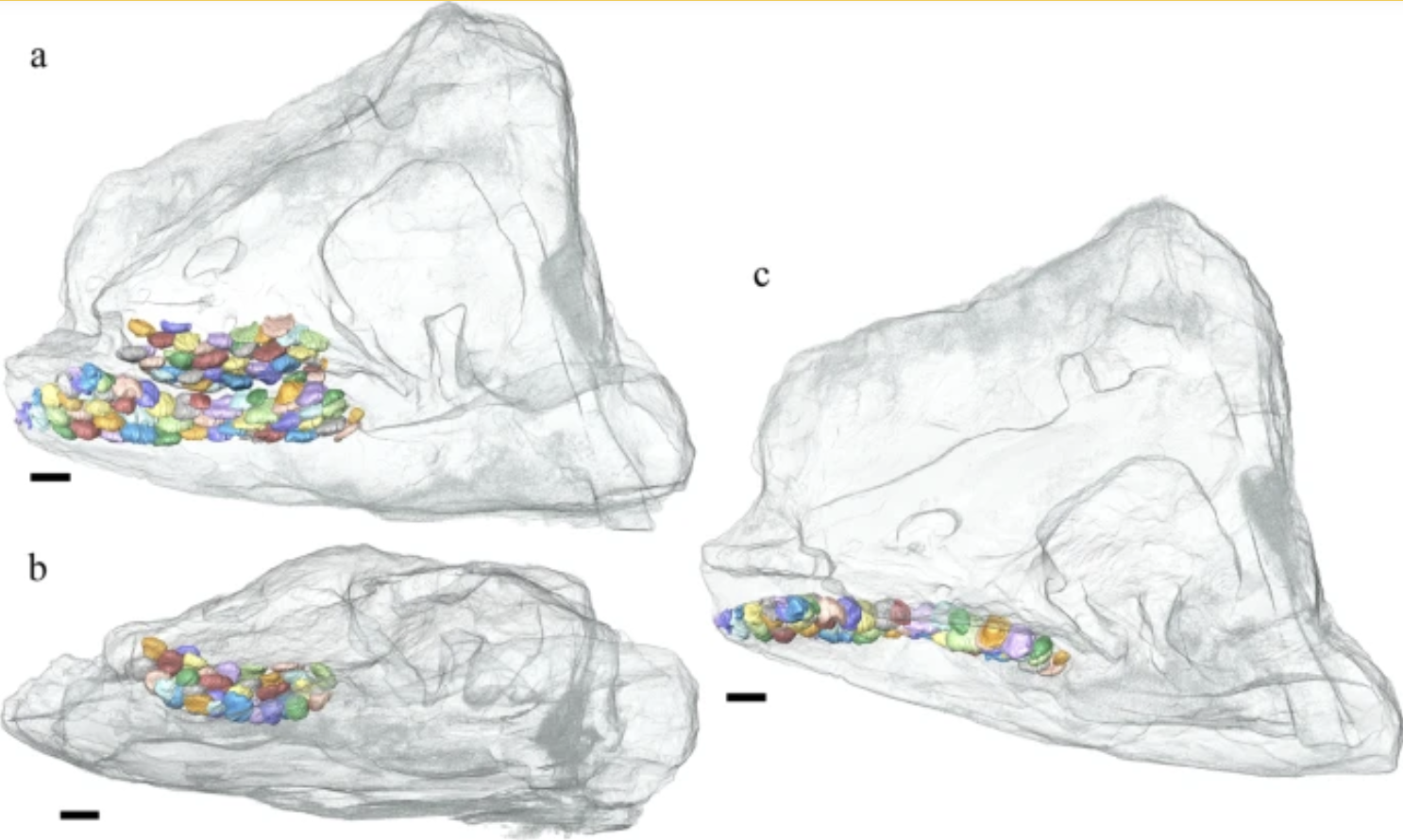
Sclerocormus
From the Early Triassic of Southern China: an ichthysauriform with:- button-like crushing teeth
- an extremely long tail
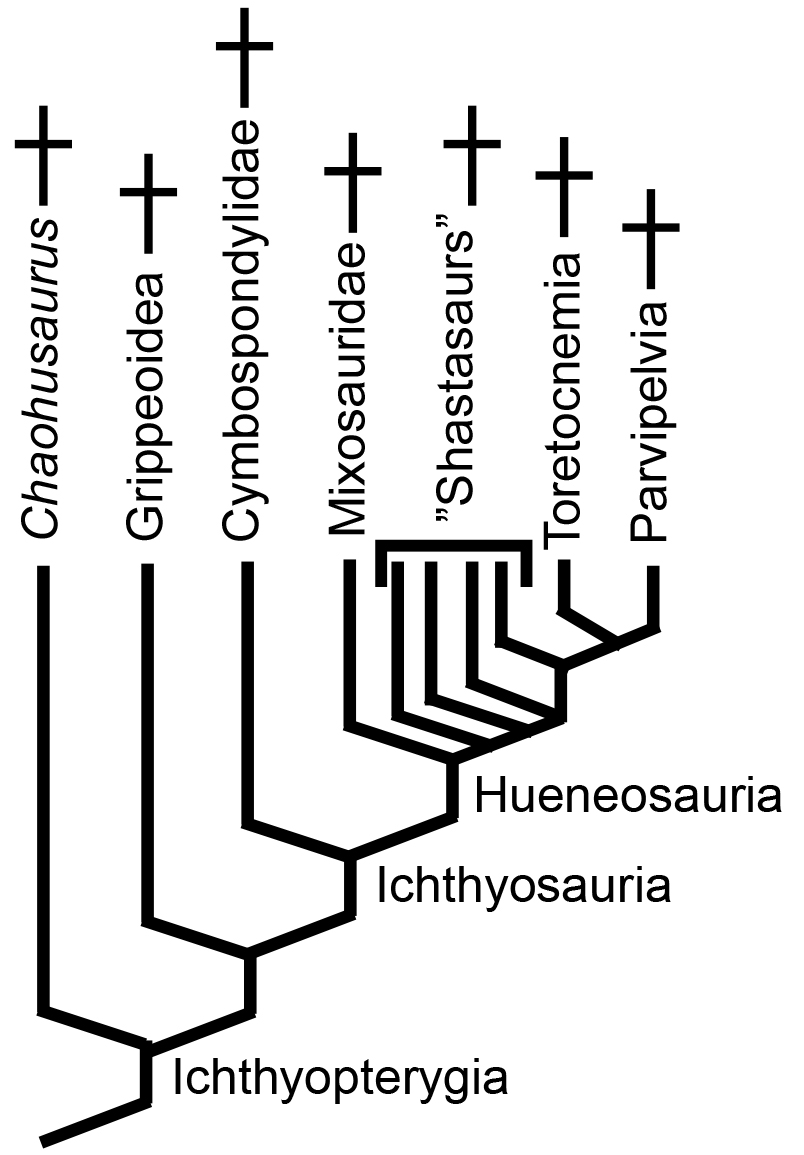
The real fun: Starting with Motani et al., 2015. phylogenetically analyzed representatives of the above taxa to obtain the result at right, finding an aquatic clade containing all of them (Corresponding to the traditional "Euryapsida." Important taxon names defined in the literature:
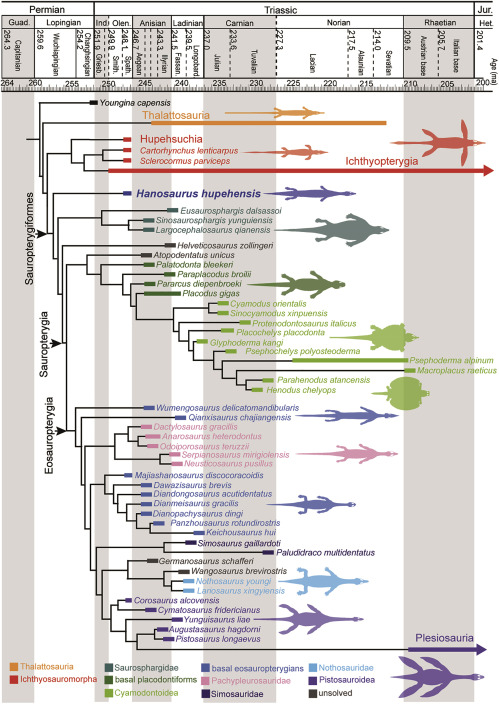
- Ichthyosauromorpha: ∇ (Hupehsuchia, Ichthyopterygia)
- Ichthyosauriformes: (Ichthyopterygia ~ Hupehsuchia)
- Omphalosauridae: ∇ (Cartorhynchus, Omphalosaurus)
- Sauropterygiformes: ∇ (Hanosaurus, Placodus, Plesiosaurus)
- In Ichthyopterygia, the pterygoids lap beneath the braincase in a manner suggestive of sauropterygians.
- At least one specimen of the parvipelvian ichthyosaur Stenopterygius has cartilagenous accessory supraspinous elements suggestive of hupehsuchians.
- Fragile:
- Minor changes in analytic assumptions reported by Motani et al., 2015 altered results.
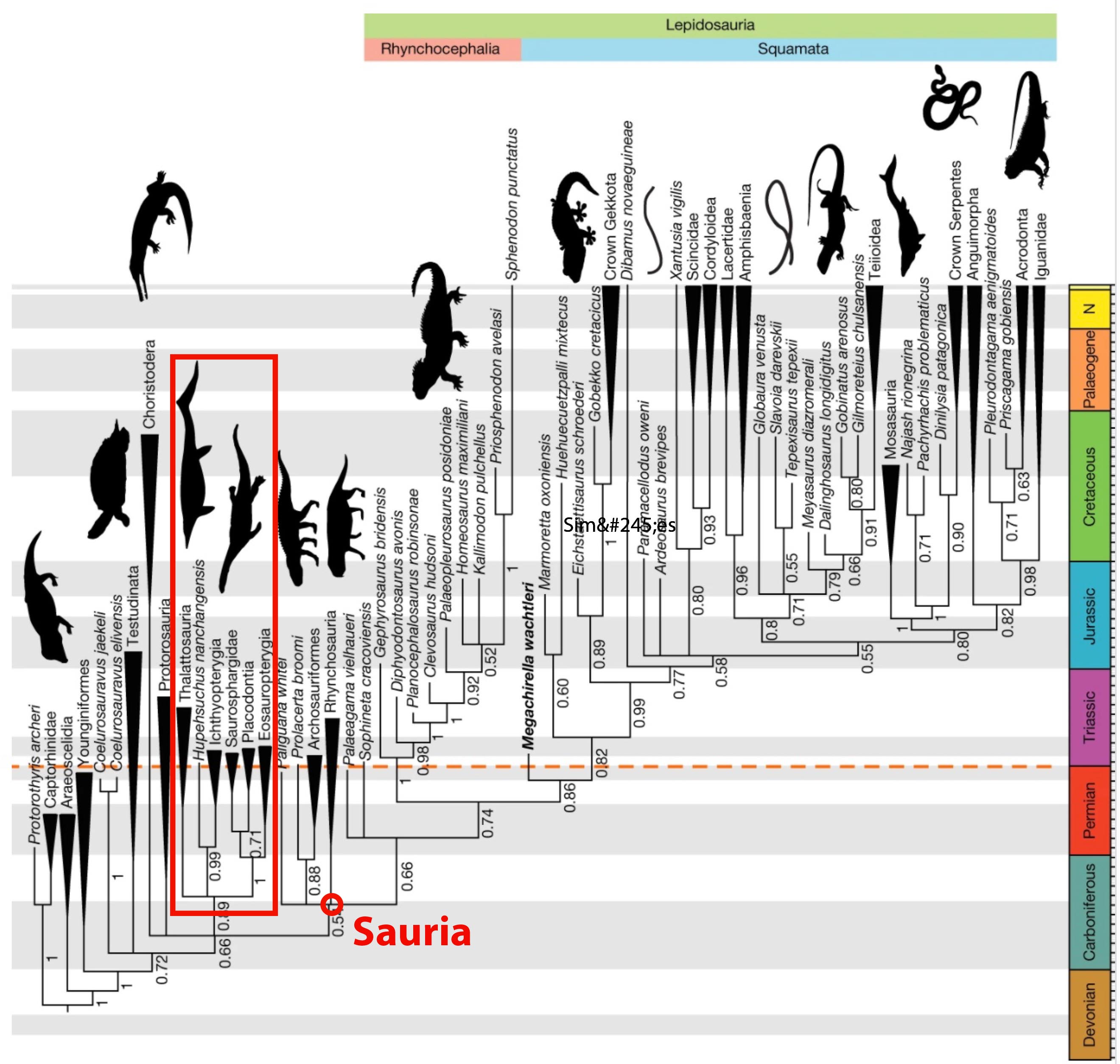
- Simões et al., 2018 recover the marine reptile clade just outside of Sauria.
- But then with minor modifications, Simões et al., 2020 recover it at the base of Archosauromorpha.
- The archosauromorph affinities of the marine clade is vindicated by Liu et al., 2024.
- Robust:
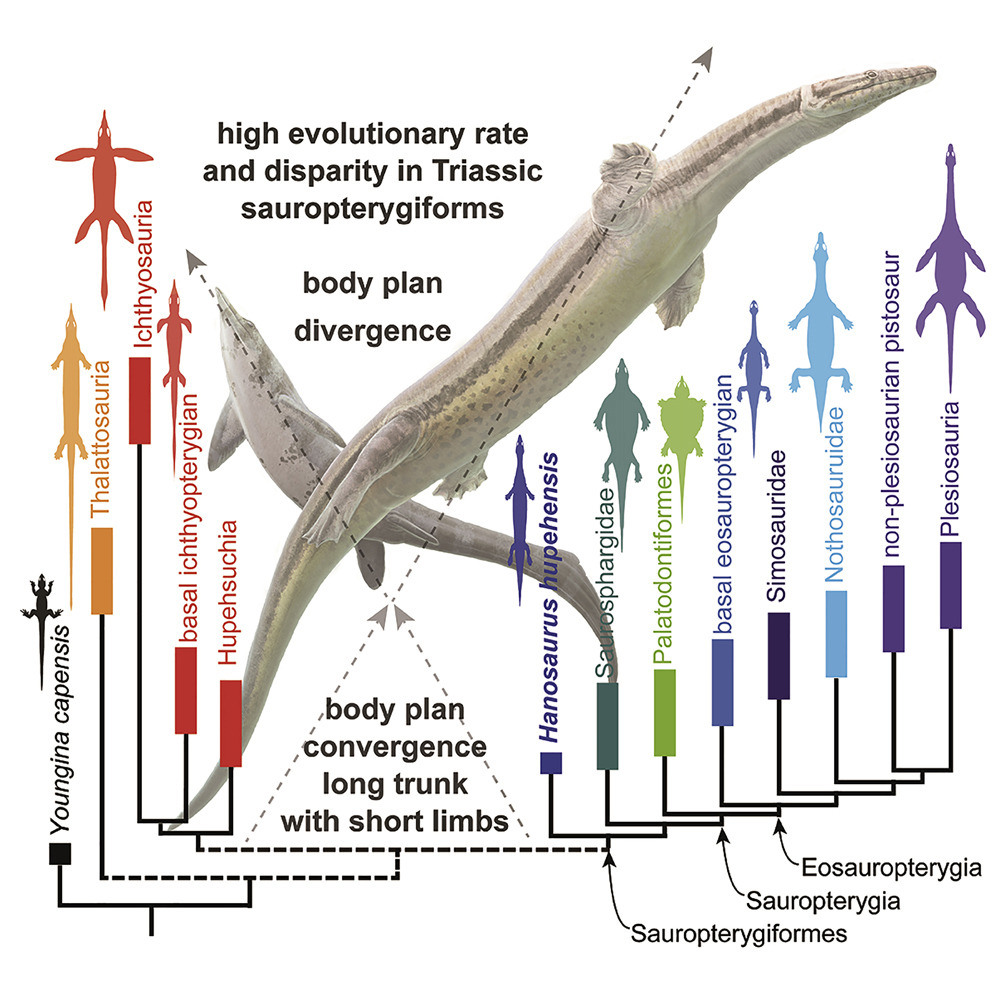
- The Triassic marine groups:
- Ichthyosaurs and their relatives: Ichthyopterygia, Hupehsuchia, and
- Sauropterygiformes: Sauropterygia, Saurosphargidae, Hanosaurus, and kin
- Thalattosauria
- Ichthyosaurs and their relatives: Ichthyopterygia, Hupehsuchia, and
- That clade branches from very near the common ancestor of Sauria.
- Concerns have been voiced that convergent adaptations to marine life might bias phylogenetic analyses, resulting in the recovery of a false marine clade, however the basic structure presented here appears even when these characters are omitted (Scheyer et al., 2017).
- The Triassic marine groups:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
The Archaeopteryx of euryapsids
What we would most love to see is a creature close to the ancestry of Euryapsida when it was just beginning to transition to an aquatic life-style - the Archaeopteryx of euryapsids. We don't have it, but can infer that it lived in the Late Permian. But we are getting close. Wang et al., 2022 describe Hanosaurus hupehensis, the sister taxon of sauropterygians and saurosphargids and basal member of Sauropterygiformes. The morphological gap between this long, slender, short-limbed creature, basal ichthyopterygians, and thalattosaurs is not so great.{kind=link}
And there is hope: The Nanzhang-Yuan'an marine reptile fauna of Hubei Province, China records conditions 4-5 my after the End Permian Extinction Event, and has recently been the source of many interesting taxa (Cartorhynchus, Sclerocormus, Pomolispondylus, Atopodentatus, Hanosaurus). God willing, it has more yet to offer. And if its stratigraphy can be solidified, Svalbard may offer key insights. Watch this space.
- Remi Allemand, Benjamin C. Moon & Dennis F. A. E. Voeten. 2022. The Paleoneurology of Ichthyopterygia and Sauropterygia: Diverse Endocranial Anatomies of Secondarily Aquatic Diapsids. In In: Dozo, M.T., Paulina-Carabajal, A., Macrini, T.E., Walsh, S. (eds) Paleoneurology of Amniotes . Springer, Cham. https://doi-org.proxy-um.researchport.umd.edu/10.1007/978-3-031-13983-3_3
- Bastiaans, D. 2024. Thalattosauria in time and space: a review of thalattosaur spatiotemporal occurrences, presumed evolutionary relationships and current ecological hypotheses. Swiss J Palaeontol 143, 36 (2024). https://doi.org/10.1186/s13358-024-00333-6
- Roger Benson, Mark Evans, Patrick Druckenmiller. 2012. High Diversity, Low Disparity and Small Body Size in Plesiosaurs (Reptilia, Sauropterygia) from the TriassicĐJurassic Boundary. PLosOne 7(3).
- Aurelien Bernard, Christophe Lecuyer, Peggy Vincent, Romain Amiot, Nathalie Bardet, Eric Buffataut, Gille Cuny, Francois Martineau, Jaen-Michel Malin, and Abel Prieur. 2010. Regulation of Body Temperature by Some Mesozoic Marine Reptiles. Science 328(5984) 1379-1382.
- Gabriele Bindelini, Andrzey Wolniewicz, Feiko Miedema, Torsten schemer, and Cristiano dal Sasso. 2021. Cranial anatomy of Besanosaurus leptorhynchus Dal Sasso & Pinna, 1996 (Reptilia: Ichthyosauria) from the Middle Triassic Besano Formation of Monte San Giorgio, Italy/Switzerland: taxonomic and palaeobiological implications. PeerJ Paleontology and Evolutionary Science (May 6, 2021).
- Roger Benson, Mark Evans, Patrick Druckenmiller. 2012. High Diversity, Low Disparity and Small Body Size in Plesiosaurs (Reptilia, Sauropterygia) from the TriassicĐJurassic Boundary. PLosOne 7(3).
- Magdalena Borsuk-Bialynicka and Susan Evans. 2009. A long-necked archosauromorph from the Early Triassic of Poland. Paleontologica Polonica 65: 203Đ234.
- Georgina Bunker, David M. Martill, Roy E. Smith, Samir Zouhri, Nick Longrich. 2022. Plesiosaurs from the fluvial Kem Kem Group (mid-Cretaceous) of eastern Morocco and a review of non-marine plesiosaurs. Cretaceous Research 140, 2022.
- Robert Carroll, 1985. Plesiosaur ancestors from the Upper Permian of Madagascar. Philosophical Transactions of the Royal Society of London B 293: 315-383
- Robert Carroll and Z-M Dong, 1991. Hupehsuchus, an enigmatic reptile from the Triassic of China, and the problem of establishing relationships. Philosophical Transactions of the Royal Society of London B 331:131-153.
- Yen-nien Cheng, Xiao-chun Wu, and Qiang Ji, 2004. Triassic marine reptiles gave birth to live young. Nature 432, 383-386.
- Cheng, L.; Chen, X. H.; Shang, Q. H.; Wu, X. C. 2014. A new marine reptile from the Triassic of China, with a highly specialized feeding adaptation. Naturwissenschaften. 101, 3.
- Cheng, Long.; Benjamin C. Moon; Shang, Chunbo Yan, Ryosuke Motani, Dayong Jiang, Zhihui An, Zichen Fang. 2022. The oldest record of Saurosphargiformes (Diapsida) from South China could fill an ecological gap in the Early Triassic biotic recovery. PeerJ Life and Environment, July 2022
- Chun Li, Da-Young Jiang, Long Cheng, Xiao-Chun Wu, Olivier Rieppel, 2014. A new species of Largocephalosaurus (Diapsida: Saurosphargidae), with implications for the morphological diversity and phylogeny of the group. Geological Magazine 151(1) 100-120.
- Chun, Li; Rieppel, Olivier; Long, Cheng; Fraser, Nicholas C, 2016. The earliest herbivorous marine reptile and its remarkable jaw apparatus. Science Advances. 2, 5.
- Mats E. Eriksson, Randolph De La Garza, Esben Horn, Johan Lindgren. 2022. A review of ichthyosaur (Reptilia, Ichthyopterygia) soft tissues with implications for life reconstructions. Earth-Science Reviews, (226), 103965, ISSN 0012-8252,
- Fang, ZC., Li, JL., Yan, CB. et al. 2023. First filter feeding in the Early Triassic: cranial morphological convergence between Hupehsuchus and baleen whales. BMC Ecol Evo 23, 36 (2023). https://doi.org/10.1186/s12862-023-02143-9
- C. V. Fleischle, T. Wintrich, P. M. Sander, 2018. Quantitative histological models suggest endothermy in plesiosaurs. BMC Evol Biol. 2013; 13: 123
- Susana Gutarra, Thomas L. Stubbs, Benjamin C. Moon, Beatrice H. Heighton, Michael J. Benton, 2023. The locomotor ecomorphology of Mesozoic marine reptiles. Palaeontology 66(2) e12645.
- Yi-Wei Hu, Qiang Li & Jun Liu, 2024. A new pachypleurosaur (Reptilia: Sauropterygia) from the Middle Triassic of southwestern China and its phylogenetic and biogeographic implications. Swiss J Palaeontol 143, 1 (2024). https://doi.org/10.1186/s13358-023-00292-4
- Sarah Jamison-Todd, Benjamin C. Moon, Andre J. Rowe, Matt Williams, Michael J. Benton, 2022. Dietary niche partitioning in Early Jurassic ichthyosaurs from Strawberry Bank. Journal of Anatomy 241(6): 1409-1423
- Benjamin Kear, Victoria Engelschiøn, Øyvind Hammer, Aubrey Roberts, and Jørn Hurum, 2022. Earliest Triassic ichthyosaur fossils push back oceanic reptile origins. Current Biology 33(5).
- A Krahl, N. Klein, P. M. Sander, 2022. Determination of muscle strength and function in plesiosaur limbs: finite element structural analyses of Cryptoclidus eurymerus humerus and femur. PeerJ. 2018; 6: e4955.
- Anna Krahl, Andreas Lipphaus, P. M. Sander, Ulrich Witzel, 2022. Evolutionary implications of the divergent long bone histologies of Nothosaurus and Pistosaurus (Sauropterygia, Triassic). PeerJ. 2022;..
- Li Ji , Lehrmann Daniel J. , Luo Yong-ming , Adams Thomas L. , Yu Mei-yi , Liao Jun-ling , Qin Yan-jiao , Li Yu-lan , Wang Yue, 2023. Soft tissue preservation in the Triassic pachypleurosaur Keichousaurus hui: evidence for digestive tract anatomy, diet, and feeding behavior. Frontiers in Ecology and Evolution 11 DOI=10.3389/fevo.2023.1186354
- Liu, J., Wu, F., & Qiao, Y., 2024. A new basal hupehsuchian from the Early Triassic of South China and its implication to the phylogenetic relationships of Ichthyosauromorpha (Reptilia: Diapsida). Historical Biology, 1Đ11. https://doi.org/10.1080/08912963.2024.2354791
- Cheng Long, Ryosuke Motani, Jiang Da-yong, Yan Chun-bo, Andrea Tintori, and Olivier Rieppel, 2019. Early Triassic marine reptile representing the oldest record of unusually small eyes in reptiles indicating non-visual prey detection. Nature Scientific Reports, 9:152.
- Michael Maisch. 1997. Phylogeny, systematics, and origin of the Ichthyosauria - the state of the art. Palaeodiversity 3: 151-214
- Michael Maisch and Andreas Matzke. 2000. The mixosaurid ichthyosaur Conntectopalatus from the Middle Triassic of the German Basin. Lethaea 33: 71-74
- Miguel Marx, Peter Sjovall, Benjamin Kear, Martin Jarenmark, Mats Eriksson, Svan Sachs, Klaus Nilkens, Michiel Op De Beeck, Johan Lindgren. 2025. Skin, scales, and cells in a Jurassic plesiosaur. Current Biology, Feb, 2025
- Judy Massare. 2021. Narial structures in Ichthyosaurus and other Early Jurassic ichthyosaurs as precursors to a completely subdivided naris. Paludicola 13(2):128-139
- John Merck. 1997. A phylogenetic analysis of the euryapsid reptiles. PhD Dissertation. University of Texas at Austin. 708 pp.
- Ryosuke Motani. 1999. Phylogeny of the Ichthyopterygia. Journal of Vertebrate Paleontology 19: 472 - 495
- Ryosuke Motani, Nachio Minoura, Tatsuro Ando. 1998. Ichthyosaurian relationships illuminated by new primitive skeletons from Japan. Nature 393, 255 - 257
- Ryosuke Motani, Da-yong Jiang, Andrea Tintori, Olivier Rieppel, Guan-bao Chen. 2014. Terrestrial Origin of Viviparity in Mesozoic Marine Reptiles Indicated by Early Triassic Embryonic Fossils. PLoS ONE 9(2): e88640. doi:10.1371/journal.pone.0088640, 255 - 257
- Ryosuke Motani, Da-Yong Jiang, Guan-Bao Chen, Andrea Tintori, Olivier Rieppel, Cheng Ji, and Jian-Dong Huang. 2015. A basal ichthyosauriform with a short snout from the Lower Triassic of China. Nature 517, 485-488.
- James M. Neenan, Nicole Klein, and Torsten M. Scheyer. 2013. European origin of placodont marine reptiles and the evolution of crushing dentition in Placodontia. Nature Communications 4, 1621 (2013)
- Elizabeth Nicholls. 1999. A reexamination of Thalattosaurus and Nectosaurus and the relationships of the Thalattosauria (Reptilia: Diapsida). PaleoBios 19(1): 1-29,.
- Elizabeth Nicholls and Makoto Manabe. 2004. Giant ichthyosaurs of the TriassicŃa new species of Shonisaurus from the Pardonet Formation (Norian: Late Triassic) of British Columbia. Journal of Vertebrate Paleontology, 24(4): 838-849.
- F. R. O'Keefe, L. M. Chiappe, 2011. Viviparity and K-Selected Life History in a Mesozoic Marine Plesiosaur (Reptilia, Sauropterygia). Science 333(6044), 870-873.
- F. R. O'Keefe, R. A. Otero, S. Soto-Acuna, J. P. O'gorman, S. J. Godfrey, and S. Chatterjee, 2017. Cranial anatomy of Morturneria seymourensis from Antarctica, and the evolution of filter feeding in plesiosaurs of the Austral Late Cretaceous. Journal of Vertebrate Paleontology 37(4).
- Ali Pourfarzan, Donald M Henderson, and Jaime G Wong. 2022. Fluid dynamics, scaling laws and plesiosaur locomotion. Bioinspiration and Biomimetics 17(5)
- Yu Qiao, Jun Liu, Andrzej S. Wolniewicz, Masaya Iijima, Yuefeng Shen, Tanja Wintrich, Qiang Li & P. Martin Sander. 2022. A globally distributed durophagous marine reptile clade supports the rapid recovery of pelagic ecosystems after the Permo-Triassic mass extinction. Commun Biol 5, 1242 (2022)
- Christiano Renesto, Cristiano dal Sasso, Fabio Fogliazza, and Cinzia Ragni. 2020. New findings reveal that the Middle Triassic ichthyosaur Mixosaurus cornalianus is the oldest amniote with a dorsal fin. Acta Palaeontologica Polonica 65(3): 511Đ522
- Olivier Rieppel. 1997. Part 2: Sauropterygia. Introduction. In Callaway, J. M and Nicholls, E.L. (eds). Ancient Marine Reptiles. Academic press. London. 107-119.
- Rieppel, O., Sander, P. M., & Storrs, G. W. (2002). The skull of the pistosaur Augustasaurus from the Middle Triassic of northwestern Nevada. Journal of Vertebrate Paleontology, 22(3), 577Đ592.
- Torsten M. Scheyer, James M. Neenan, Timea Bodogan, Heinz Furrer, Christian Obrist & Mathieu Plamondon. 2017. A new, exceptionally preserved juvenile specimen of Eusaurosphargis dalsassoi (Diapsida) and implications for Mesozoic marine diapsid phylogeny. Sci Rep 7, 4406 (2017). https://doi.org/10.1038/s41598-017-04514-x
- A. G. Sennikov. 2019. Peculiarities of the Structure and Locomotor Function of the Tail in Sauropterygia. Biology Bulletin volume 46, pages751-762
- Qing-Hua Shang, Xiao-Chun Wu, Chun Li. 2020. New Ladinian Nothosauroid (Sauropterygia) from Fuyuan Yunnan Province, China. Journal of Vertebrate Paleontology 40(3) e1789651-2.
- Tiago Simões, Michael Caldwell, Mateusz Talanda, Massimo Bernardi, Alessandro Palci, Oksana Vernygora, Federico Bernardini, Lucia Mancini, and Randall L. Nydam. 2018. The origin of squamates revealed by a Middle Triassic lizard from the Italian Alps. Nature volume 557, pages706Đ709.
- Tiago Simões, Oksana Vernygora, Michael W. Caldwell & Stephanie E. Pierce . 2020. Megaevolutionary dynamics and the timing of evolutionary innovation in reptiles. Nature Communications volume 11, Article number: 3322 (2020).
- Glenn Storrs. 1991. Anatomy and Relationships of Corosaurus alcovensis (Diapsida: Sauropterygia) and the Triassic Alcove Limestone of Wyoming. Bulletin 44. Peabody Museum of Natural History, Yale University. 141 pp.
- Wei Wang, Qinghua Shang, Long Cheng, Xiao-Chun Wu, Chun Li. 2022. Ancestral body plan and adaptive radiation of sauropterygian marine reptiles. iScience 25(12)
- Tanja Wintrich, Shoji Hayashi, Alexandra Houssaye, Yasuhisa Nakajima, and P. Martin Sander. 2017. A Triassic plesiosaurian skeleton and bone histology inform on evolution of a unique body plan. Science Advances 3(12)
- Andrzej S. Wolniewicz, Yuefeng Shen, Qiang Li, Yuanyuan Sun, Yu Qiao, Yajie Chen and Jun Liu. 2023. An armored marine reptile from the Early Triassic of South China and its phylogenetic and evolutionary implications. eLife 12:e83163.
- Xiao-Chun Wu, Yen-Nien Cheng, Chun Li, Li-Jun Zhao, and Tamaki Sato. 2011. New information on Wumengosaurus delicatomandibularis Jiang et al. 2008 (Diapsida: Sauropterygia) with a revision of the morphology and phylogeny of the taxon. Journal of Vertebrate Paleontology 31(1):70-83,
- Guang-Hui Xu, Qing-Hua Shang, Wei Wang, Yi Ren, Hong Lei, Jun-Ling Liao, Li-Jun Zhoa, and Chun Li. 2023. A new long-snouted marine reptile from the Middle Triassic of China illuminates pachypleurosauroid evolution. Scientific Reports 13(16).
- Bastiaans, D. 2024. Thalattosauria in time and space: a review of thalattosaur spatiotemporal occurrences, presumed evolutionary relationships and current ecological hypotheses. Swiss J Palaeontol 143, 36 (2024). https://doi.org/10.1186/s13358-024-00333-6