


"Hope is the thing with feathers
That perches in the soul
And sings the tune without the words
And never stops at all." Emily Dickinson (1861)
and
"The members of class Aves so nearly approach the Reptilia in all the essential and fundamental points of their structure, that the phrase "Birds are greatly modified Reptiles" would hardly be an exaggerated expression of the closeness of that resemblance." -- Thomas H. Huxley. 1867. On the classification of Birds; and on the taxonomic value of the modification of certain cranial bones observed in that class. Proceedings of the Zoological Society of London 1867: 415-472.
BIG QUESTION: How did birds evolve from (other) dinosaurs, and how did they learn to fly?
Up, Up, and AWAY! Flying Animals
In the history of life, only four groups of animals have evolved powered flight:
To possess powered flight is to be volant. But there are other forms of aerial locomotion. These include the essentialy vertical parachuting and the more controlled and linear gliding
While there are only three groups of volant vertebrates, there are many modern gliding/parachuting ones: flying fish; flying frogs; flying lizards; flying snakes; sugar gliders (gliding marsupials); flying squirrels; colugos, or "flying lemurs". In addition, there were various extinct gliding reptiles and mammals in the Permian and Mesozoic.
Aves is the traditional Linnaean name for birds, and is used presently for crown-group birds (i.e., all descendants of the concestor of all living birds.) Avian is the adjective for issues of or concerning for members of Aves.
Pterosaurs are not birds. Nor are they dinosaurs. In contrast, EVERY bird species is a dinosaur! (Recall that Dinosauria = the concestor of Iguanodon, Megalosaurus, and Diplodocus and all of its descendants.)
At present there are between 9600 to 18,000 living species (depending on whether you are a lumper or splitter) of birds, and just 2000 years ago this number was probably an additional 3,000 more. (The difference was due to the extinction of many species of birds unique to individual islands of the Pacific and Indian Oceans: these suffered greatly as Polynesians (and later, Europeans) introduced pigs, rats, dogs, and goats to the islands.) In contrast, living mammals consist of some 4500-5600 species or so.
Many features make birds distinctive from their close living relatives among the reptiles:
It is almost impossible to mistake a bird for any other group of animal in the modern world, and vice versa. But as we will see, this is not true in the fossil record.
Because the distinctiveness of birds, it was difficult for biologists to determine the closest living relation among modern animals. Jean-Baptiste de Lamarck suggested that turtles were the closest relative to birds, with sea turtles and penguins representing transitional forms.
The fossil record of transitional forms between Aves and other reptiles was greatly improved by the discovery of Archaeopteryx lithographica from the Late Jurassic Solnhofen Lithographic Limestone of Bavaria, Germany. The first specimen (a feather) and the first skeleton (found in 1861; acquired by the Natural History Museum (London) in 1863) showed an animal that had:
The skeleton was not complete, but still achieved considerable attention. In the 1860s paleontologist T.H. Huxley used the London specimen as evidence that the newly discovered Dinosauria were more closely related to birds than to other groups of living or fossil reptiles.
In 1874 (or 1875) an even more complete skeleton of Archaeopteryx was discovered; in 1881 it was acquired by the Humboldt Museum in Berlin. This confirmed the presence of teeth in its jaws, and showed presence of individual fingers with claws.
In the 1870s Yale paleontologist described Cretaceous birds from the Niobrara Chalk of Kansas. These included flightless swimming Hesperornis and flying Ichthyornis. Both were far more closely related to modern birds than was Archaeopteryx, but still retained teeth in their jaws.
In the late 19th Century and earliest 20th Century several models of bird origins were suggested:
With the discovery of coelurosaurian dinosaurs like Compsognathus and Ornitholestes some paleontologists and others were convinced of the latter idea. When writing of Ornitholestes, H.N. Hutchinson (1910, Extinct Monsters and Creatures of Other Days) wrote:
The dinosaurian origin of bird hypothesis, however, was deflected by the well-meaning work of anatomical illustrator and amateur paleontologist Gerhard Heilmann (1855-1946). He wrote a series of articles in Danish between 1913 and 1916 which were collected and translated into English in 1923 as The Origin of Birds. He conclusive showed what Huxley and others had proposed: birds were some kind of archosaur. He compared birds to various groups of dinosaurs; to pterosaurs; to early crocodylomorphs; and to "thecodonts" (the then-fashionable name for the paraphyletic grade of basal archosauriforms, non-crocodylomorph pseudosuchians, and non-pterosaur, non-dinosaur ornithodirans).
Heilmann showed considerable shared derived features between birds and coelurosaurian theropods. But as no one had yet demonstrated a clavicle in a theropod, there was no feature that could evolve into a furcula. So the similarities between birds and dinosaurs had to be convergences. Instead, Heilmann pointed to "thecodonts" (in this case, basal archosauriforms) such as the then-recently discovered Euparkeria of the Middle Triassic of South Africa, which he deemed "sufficiently primitive" to be a bird ancestor.
Heilmann also proposed a scenario for the origin of birds: quadrupedal thecodonts which had adapted to life in the trees. These grew elongated scales which served as gliding surfaces on the arms, body, and tail in a long-armed hypothetical "proavis", which was the ancestor of Archaeopteryx and thus to later birds.
Heilmann's book was phenomenally successful, and dominated thinking on bird origins until the 1970s. Standard pictures for archosaur evolution showed birds, pterosaurs, ornithischian dinosaurs, saurischian dinosaurs, and crocodylomorphs all radiating independently from a common thecodont stock. (Note: dinosaurs were considered at best diphyletic [two independent origins] within Archosauria.) And the arboreal gliding proavis was the standard model for the origin of avian flight.
In 1969 John Ostrom of Yale described the dromaeosaurid Deinonychus. This was the first raptor dinosaur (deinonychosaur) known from relatively complete material, and showed considerable similarity with Archaeopteryx. In fact, Ostrom discovered a specimen of Archaeopteryx (miscatalogued as a specimen of pterosaur!) in a museum in the Netherlands. From comparisons of their skeletons, Ostrom demonstrated that deinonychosaurs (and some other coelurosaurs) and birds shared many traits, such as:
(Just recently that specimen that Ostrom discovered as a pterosaur was re-examined and found not to be Archaeopteryx, either. It is from the more primitive stem-bird lineage Anchiornithidae, and named Ostromia in his honor.)
Ostrom revived the dinosaurian origin of birds hypothesis, which a new specific sister-group relationship between Deinonychosauria and birds. Subsequently Heilmann's primary objection was removed: it was found that theropods did not merely have clavicles; they in fact ALL possessed a furcula!
The initial cladistic analyses of dinosaurs and archosaurs in general (in the early 1980s) focused on testing the dinosaurian origin of birds hypothesis. Ostrom's idea stood up to this test. In 1986 Jacques Gauthier (who would a decade or so later have Ostrom's chair at Yale after the latter's retirement) coined the name "Avialae" ("bird wings") for the group comprised of Aves and all taxa sharing a more recent common ancestor with Aves than with Deinonychosauria. At the time non-avian members of Avialae were still restricted basically to Archaeopteryx, Hesperornis, and Ichthyornis.
This hypothesis was not without its detractors. Prominent among these were paleornithologists Alan Feduccia and Larry Martin and physiologist John Ruben. At conferences in the 1980s, 1990s, and 2000s they referred to themselves as the "BAND" (for "Birds Are Not Dinosaurs"). Their primary arguments during the 1980s and 1990s were that Heilmann was correct and the similarity between coelurosaurs and birds were just convergences. Instead they argued that newly-discovered Middle or Late Triassic quadrupedal "thecodonts" (actually not even archosauriforms, but primitive diapers) such as Megalancosaurus and Longisquama were the ancestors of birds.
Simultaneous with these arguments, an explosion of discoveries (which has definitely NOT stopped!) of Mesozoic avialians has gone on. These are primarily due to several Lagerstätten yielding dozens of new genera (and literally thousands of specimens, often nearly complete skeletons and feather impressions):
Because of this, many steps and morphological transitions along the lineage leading to Aves are now documented among Jurassic and Cretaceous avialians and other coelurosaurs. Indeed, uniquely among flying vertebrates we have a problem in indicating exactly when "birdiness" is achieved, because we have so many gradual transitions in between. (In contrast, both pterosaurs and bats appear in the fossil record as pterosaurs and bats, not proto-pterosaurs and proto-bats.)
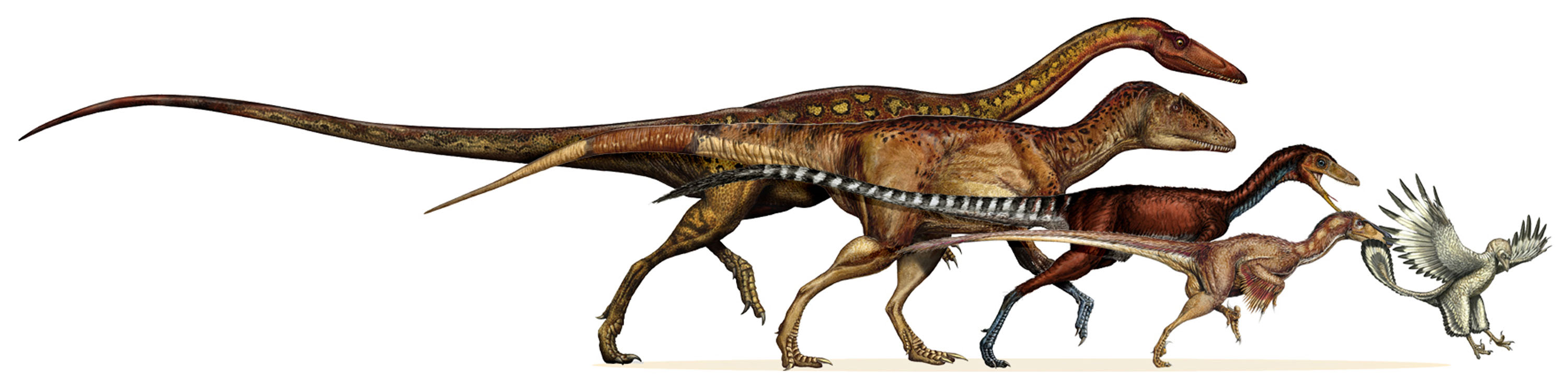
Until recently, the basalmost theropod known to have feathers was Archaeopteryx, although some researchers speculated that other theropods had them as well. And the feathers of Archaeopteryx were identical to the feathers of modern birds, so they didn't reveal much about the early phases of these structures. But fossils from lake sediments of the Middle and Late Jurassic of China and Siberia and the Early Cretaceous of Spain and China have given us a better understanding of the distribution of feathers and protofeather structures.
It was not just avialians that were discovered in these Lagerstätten. These also revealed the presence of feathers in non-avialian dinosaurs. The first discovered of these (in 1996) were simple apparently hollow down-like tufts: plumulose structures dubbed protofeathers on the compsognathid coelurosaur Sinosauropteryx. Subsequently similar structures were found on numerous other types of primitive coelurosaurs, including 1 ton tyrannosaurs such as Yutyrannus. And their presence in the ornithischians Tianyulong and Kulindadromeus hints at the possibility that protofeathers are basal to all dinosaurs. (And if pterosaurian pycnofibres are homologous to protofeathers, these might be basal to all Ornithodira).
Protofeathers obviously don't have a flight function, since they don't form an aerodynamic surface. However, they might serve other functions:
More derived are strap-like protofeathers: broad surfaces rather than tufts.
Four major clades of coelurosaurs (Avialae, Deinonychosauria, Oviraptorosauria, and [less definitively] Ornithomimosauria) show broad pennaceous feathers on the arms; the first three (collectively called Pennaraptora) have them both on the arms and the tail. These same pennaraptoran dinosaurs are also characterized by sideways-oriented shoulder joints, semilunate carpals, and direct brooding of the eggs. In Deinonychosauria and primitive members of Avialae (but not in Oviraptorosauria, at least as far as we know), there are long leg feathers as well.
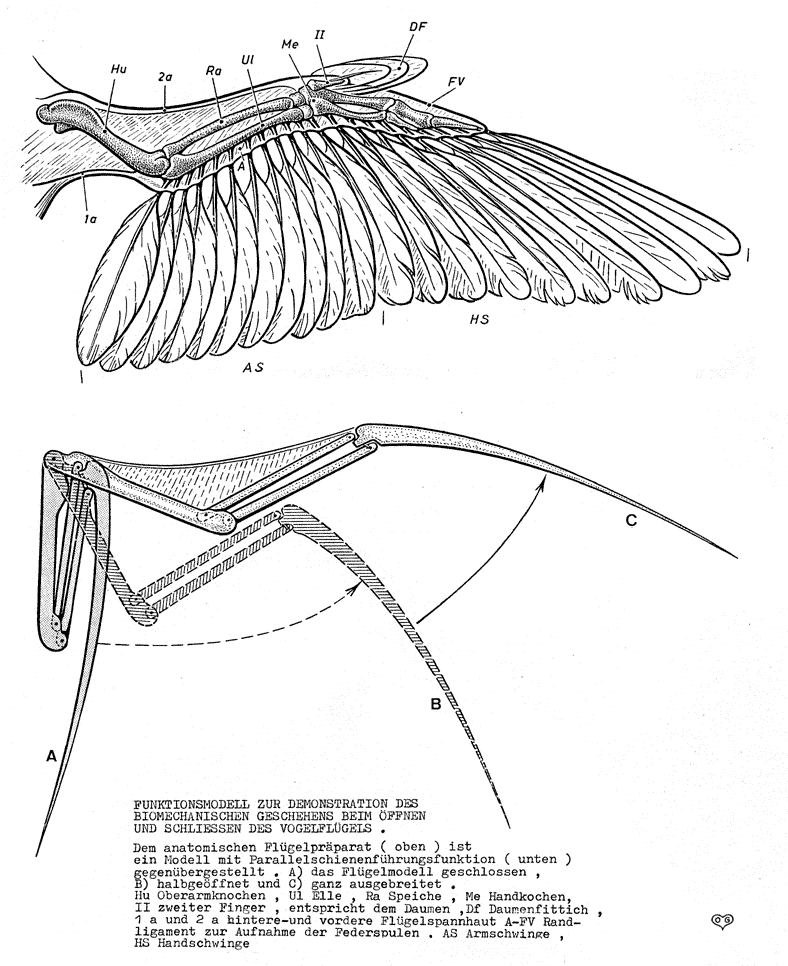
Pennaceous feathers are very diverse in modern birds: found in flight feathers on the wings and tail; contour feathers over the body; and various decorative feathers. All have a similar structure: a central shaft (rachis), with barbs coming off of it, barbules coming off the barbs, and hooklets coming off the barbules. Developmental biology shows that plumulose down and pennaceous feathers have the same developmental pattern, just with genes emphasizing different rates of development of various parts of the developing structure. These developmental stages match the observed stages in the fossil record.
The discovery of feathers on other groups of theropods has reduced the number of uniquely bird traits in Archaeopteryx. Indeed, discovery of new Jurassic feathered dinosaurs closely related to Archaeopteryx results in some phylogenetic analyses in which this "Archaeopterygidae" is not necessarily a basal clade of Avialae, but instead is a basal clade in Deinonychosauria or the sister group to Deinonychosauria and Avialae.
(Note: in response to the discovery of fully-feathered deinonychosaurs and oviraptorsaurs, the BAND have decided that these clades are NOT dinosaurs, but are basal birds convergent on coelurosaurs!)
So where do "birds" begin? Would we use Pennaraptora? Or Avialae? Or Aves (the toothless crown group)? Or some spot in between?
Evolution doesn't work by massive instantaneous transformations: a fully flying modern style bird did not hatch out of a totally non-flying dinosaurs egg! Instead, each phase along the way functioned in its own way, well enough for that dinosaur to make a living at it. And (as we'll see) it wasn't as if dinosaurs were trying to become birds; instead, each adaptive phase produced several divergent branches, some evolving in one direction and others in others.
Our knowledge of the phases of the origin of bird flight (anatomical, phylogenetic, and behavioral) have all greatly increased in the last decade. But before we see this newer understanding, let's review some terms and take an historical look at the problem.
Some key terms:
Traditionally, paleontologists have considered two main scenarios for the origin of bird flight:
(NOTE: during the 1970s-1990s, this debate was tangled up with a scientifically separate debate; that is, where birds fit phylogenetically among the archosaurs. The media in particular made the equation "'arboreal model = non-dinosaurian origin of birds'; 'cursorial model = dinosaurian origin of birds'". But these were actually separate debates. Even among those who recognized the dinosaurian origin of birds, some argued for the trees-down model, and others for the ground-up.)
This debate was primarily waged prior to the new discoveries of Early Cretaceous feathered coelurosaurs, which greatly increased our knowledge of the anatomy (integumentary and skeletal) of the basal members of the coelurosaur clades. Additionally, important observations of modern birds revealed a very significant locomotory behavior, previously overlooked.
In the early 2000s research by Ken Dial of the University of Montana's Flight Laboratory revealed a locomotory behavior in modern birds not previously realized. Birds (in this case chukar patridges) were discovered to run vertically up surfaces, aided by flapping their wings back and forth in order to generate traction against the surface. They called this behavior Wing Assisted Incline Running, or WAIR for short. They have since found many bird species with this behavior, even perfectly good fliers like pigeons:
Note that these birds are NOT climbing in the typical sense: they are literally running up the sides of trees. Dial and his team studied the ontogenetic (growth) changes in the ability for birds to use this behavior, and also experimented by trimming the feathers of birds to different lengths.
They found:
Additional work has shown that this behavior is widespread among modern birds.
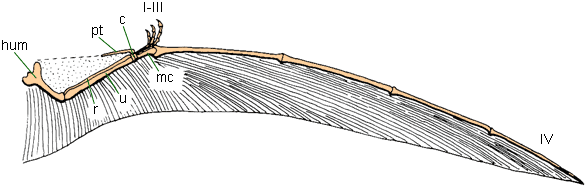
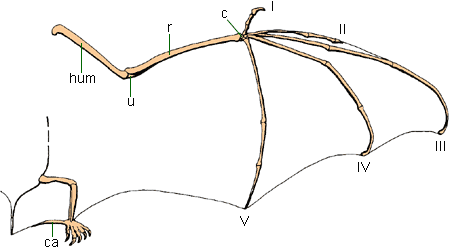
The apparatus required to use WAIR is:
All these attributes are present in many pennaraptorans (oviraptorosaurs, dromaeosaurids, troodontids, basal avialians). Additionally, modern birds use WAIR to escape predators: certainly a selective factor present in the Mesozoic, too! Furthermore, there are net selective advantages to slight increases in the length and breadth of the feathered arm surface: the sort of material that can easily be increased by natural selection.
WAIR might represent a "stepping stone" or "behavioral missing link" in the origin of flight. Small (or juvenile) maniraptorans might have used this method to escape predators. Now that they had the ability to get up into trees and other high spots, some lineages of maniraptorans might become specialized for life up on these high spots. Additional natural selection could favor further development of wing size and shape as an aid for getting back down off of high places (controlled flapping descent), or (eventually) from branch-to-branch.
Thus, WAIR serves as a functional link between cursorial and arboreal models (and organisms). It is a cursorial model in that wings begin in part as an aid to running locomotion (just vertical running); it is an arboreal model in that once pennaraptorans have an ability to get into the trees, evolution can further develop the forelimbs to get them back down to the ground. And all of these behaviors are still found in modern animals: no speculation of behaviors not currently seen needed.
A NEW SCENARIO FOR BIRD FLIGHT ORIGINS: The various recent discoveries of the skeletal and integumentary anatomy of various coelurosaurs (including basal avialians) and the behavioral and biomechanical evidence of modern birds suggests a more complete possible scenario for bird flight evolution than the historical "ground up" or "trees down" versions. Note that as with all evolutionary scenarios this would be a simplification, but the following is consistent with our current evidence: The various recent discoveries of the skeletal and integumentary anatomy of various coelurosaurs (including basal avialians) and the behavioral and biomechanical evidence of modern birds suggests a more complete possible scenario for bird flight evolution than the historical "ground up" or "trees down" versions. Note that as with all evolutionary scenarios this would be a simplification, but the following is consistent with our current evidence:
Why did Aves Survive the K/Pg?
People often note that birds--and only birds--among the dinosaurs survived the K/Pg extinction. But we can actually refine that. It is Aves--and only Aves--that survived. Other groups of Mesozoic birds, such as Ichthyornis and its kin, hesperornithines, and the diverse enantiornithines, did not survive. What makes the toothless birds different?
One possibility, of course, is that it was just luck. That is actually a difficult hypothesis to disprove (as it basically requires us to have access to alternate histories, to see if Aves goes extinct more or less often than Enantiornithes in these different timelines.)
However, there are some reasonable speculations as to why Aves might have survived while its close kin did not:
Whatever the case, the last part of the great Dinosauria clade to survive was among the most humble. Yet this lineage would diversify to become the most successful group (at least in terms of species numbers) of all the tetrapods.
Dinosaurs and feathers:
When birds had teeth:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}