

Key Points
- In the wake of the K/Pg mass extinction, mammals diversify into many modes of life.
- Different groups of mammals evolve different adaptation (particularly with regards to feeding and locomotion) for their new ecological roles.
- The rise and spread of grasslands as the world dries in the mid-Cenozoic created conditions that led to herd-dwelling grazers and pack-hunting predators.
- The Pleistocene Ice Age also produced a new selective pressure on mammals, driving many clades to adapt to the new cool-weather mammoth steppes.
- Many cold-adapted mammals had their origin in the Tibetan Plateau, already adapted to cold weather in that region, and spreading out was northern Eurasia and North America become as cold as the Asian highlands.
The small body size and burrowing habit probably favored mammalian survival relative to dinosaurs and pterosaurs, for instance. Mammal species diversity and body size both increase very quickly after the dust settled and the Cenozoic began. Indeed, the placental mammal diversification of the Paleocene Epoch is the original case study of the concept of "adaptive radiation."
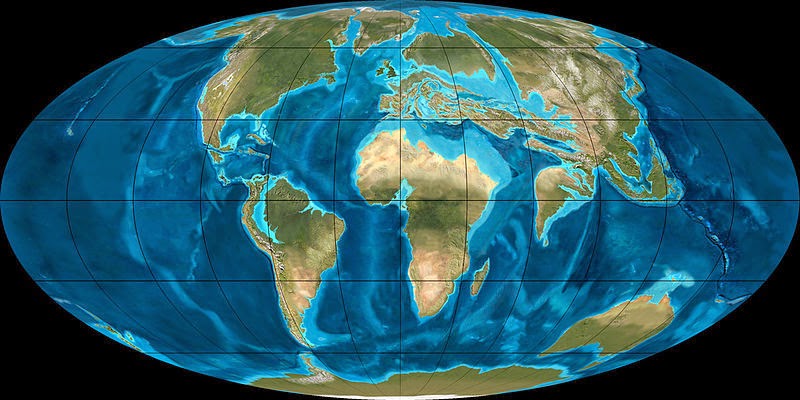
The global environment of the early Cenozoic (Paleocene and Eocene Epochs) was warmer and wetter than the modern world, not terribly dissimilar to the previous Cretaceous. Rain forests were common in many regions--even in the mountains of the American West. In the mid-Eocene it was warm enough that the Arctic Ocean in summer was covered by floating Azolla ferns. (In fact, over the space of 800,000 years the annual sinking and sequestering of these seasonal blooms in Arctic Ocean sediments drew atmospheric CO2 levels down from 3500 ppm to 650 ppm.)
The continents of the dawn of the Cenozoic were more separated than at present. As a consequence different continents were biologically isolated. As today, Australasia was an "island continent" (although connected to Antarctica, at least in the beginning), but South America, Africa, Madagascar, and India also lacked direct contact to other landmasses.
A consequence of this was that in the early Cenozoic there was a high rate of endemism (taxa unique to just one landmass). As today, Australasia was dominated by marsupials. Different groups of marsupials and non-marsupial metatheres (which were the dominant predators), as well as specialized endemic placentals. (Curiously, although marsupials are a Southern Hemisphere clade, Metatheria in the Cretaceous is only found in the Northern Hemisphere!)
The remaining regions were all dominated by various types of placentals. North America, Europe, and Asia had some limited connection, so many of the same clades were found in all three. Africa, India, and Madagascar were isolated; later in the Cenozoic the first two collide with Eurasia and there is an exchange of mammals and other animals.
As mammals evolved into apex predators, they required a new specialization: carnassials (shearing premolars and/or molars). This allowed them the slice of large chunks of meat, which ancestral teeth of mammals were not able to do. Carnassials evolved independently in the Sparassodonta (a South American clade of predatory stem-marsupials), in predatory crown-marsupials of Australasia, in "creodonts" (a paraphyletic assemblage of mid-Cenozoic carnivorous placentals in America, Eurasia, and Africa), and in Carnivora (the main clade of placental carnivorous mammals).
To give an example of the diversifications during the Paleocene and Eocene:
With isolation on different landmasses, convergences between different groups are rampant among Cenozoic mammals. Different clades evolve similar body forms due to similar life habits.
Introduction
The World for World is "Forest": Early Cenozoic Mammal Radiations
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Home on the Range: Mammals in a Cooler, Drier World
In the end of the Eocene the world began to dry and become colder. This transition is extremely quick (taking less than 100,000 years). The primary driver for this is the break up of Antarctica from Australia, forming a circumpolar current and the generation very cold bottom water. This cold bottom water (the Antarctic Bottom Water) is a major CO2 sink, sucking much of the carbon dioxide out of the atmosphere. This cooling of the world resulted in the first Antarctic glaciers.
{kind=link}
This event resulted in some extinction events of marine plankton and a major turnover of mammals. In Europe this is called the Grande Coupure ("Big Break") and in Asia and the "Mongolian Remodeling". Throughout the world many early Cenozoic groups die out.
A group that begins to thrive at this time is the grasses. The Oligocene Epoch (33.9-23.03 Ma) saw the rise of grasslands, at least in the form of desert grasslands (primarily taking over from both desert scrub and dry woodlands.) During the Miocene (23.02-5.333 Ma) short grasslands with deep soils really start to take over, and in the late Miocene and Pliocene (5.333-2.588 Ma) we see the spread of tall grasslands with exceedingly deep soils.
Grasslands are one of the most important modern biomes. There are many types and names for these: savannas and veldts of Africa, prairies of North America, pampas of South America, steppes of Eurasia, rangelands in general, etc. (Generally, savanna is used for grasslands with scattered trees, and some other terms for those few or no shrubs and trees.) (Incidentally, woodlands are generally biomes where there is >40% woody plant cover, typically smaller trees where the crowns of the trees do not overlap; while forests are biomes here there is >>40% wood plant cover, where the trees can be more than 20 m tall, and where the crowns of the trees do overlap (and thus small ground cover plants receive very little sunlight.))
Approximately 40% of the modern land surface is covered by some form of grassland. Grasslands typically have very deep soil (in large part because of the rapid turnover of plants). Grasslands are typically associated with dry but not desert conditions: only 500-900 mm/yr rain.
{kind=link}
Most of the grassland plants (both grasses and herbs) are annuals (live for just one year), so there is very rapid turnover of plants and thus VERY high productivity. In contrast, forests are dominated by trees which are decades-to-centuries old, and thus nutrients are sequestered in the plant biomass. In addition, although a given unit area of forest might have much more volume of plant matter, the great bulk of that is in the form of woody tissue which is not directly edible to typical vertebrates; in contrast, a very high percentage of a given grass plant is edible. As a consequence of rapid turnover and high percentage of accessible food, grasslands can support vast large vertebrate biomasses.
Fires are very, VERY common in grasslands. In fact, fires promote the maintenance and spread of grasslands, because grass can recover easily from fires, but trees and shrubs typically will need to recolonize in order to be present.
The changing of the environment resulted in major changes in the mammalian communities. To vertebrate zoologists, "grazer" specifies a grass-eater. (That is in fact the etymology of the word!) (Note however that marine biologists use "grazer" for almost any animal that bites off small immobile food, such as algae, polyps, bryozoans, etc.) "Grazers" is contrasted with "browsers" (animals that eat mostly herbs or the leaves of shrubs and trees), with "mixed feeders" as having both grass and browse in their diet.
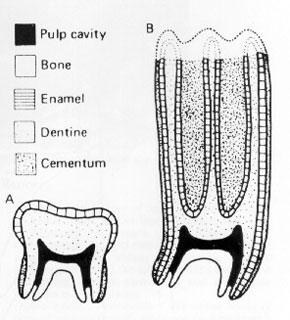
It had long been observed that the Cenozoic saw a drying of the planet, and a shift from a forest-dominated to a grass-dominated world. Certain adaptations in mammal lineages (such as the evolution of equids (horses), bovids (antelopes, including buffalo, bison, cows, goats, sheep), and the like) were thought to document evolutionary responses to this environmental shift. Grazers are distinct from browsers in a number of aspects of the jaws and teeth:
- Deeper jaws
- Deeper rooted teeth, often showing:
- Complex grinding surfaces
- Hypsodonty: high crowns to allowing them to wear down over the lifetime of the mammal due to the wear-and-tear of eating phytolith-rich grasses
- In a few groups, there is the evolution of actual hypselodonty; ever-growing teeth.
- Broader, flatter fronts of the snout to crop off a continuous surface.
{kind=link}
There are other proxies for grazing vs. browsing. For example, if you look at the surface of teeth under the microscope, browsers typically have fewer scratches and more pits, while grazers have more scratches than pitting. Also the isotopic signal of C4 grasses (about which more below) show up in the chemical composition of the bones and teeth of grazers.
Some other changes from the ancestral forested home to life of the grasslands are tendencies but are not absolute distinctions: you can find some forest dwellers with these adaptations, too:
- Larger body size (for instance, the African savanna elephant Loxodonta africana is bigger than both the African forest elephant Loxodonta cyclotis and the Indian elephant Elephas maximums (which is forest-and-woodland dweller).
- Alternatively, at least a taller stance or alert posture, to see further over the grasses
- Because the ability to hide (except in burrows) is more difficult in the open, increased emphasis on cursorial (running) adaptations (for escape, and for pursuit in the case of predators), such as:
- Elongation of the lower parts of the limb
- Concentrations of the muscles near the hips and shoulders
- Narrow, compact feet
In the case of ungulate (hoofed mammals), a more solid hoof
- Forming big herds for defense (and, in carnivores, packs in response)
{kind=link}
{kind=link}
{kind=link}
Many types of mammal (and other animals, to be fair!) diversified in the grasslands, but some of the greatest successes were among the ungulates: in particular, the Artiodactyla (the even-toed ungulates) and the Perissodactyla (the odd-toed ungulates). Indeed, within these two groups, there was huge diversifications in the Bovidae and the Equidae (respectively), both of whom were ancestrally forest-dwelling clades. The evolution of the equids in particular are an excellent example:
{kind=link}
- Early Eocene forest equids like Eohippus and Sifrhippus were quite small, with low-crown teeth and feet ending in four or five toes
- Mid-Late Eocene equids show somewhat higher crowns and somewhat larger body size, walking on three toes (but still more pad-footed than hoofed)
- Early Oligocene horses like Mesohippus are larger and longer legged; later Oligocene horses like Miohippus take these adaptations further
- Early Miocene horses include some forms still adapted to more forested environments, but show a major radiation of grazing horses. These (such as Parahippus and Merychippus) were effectively one-toed (the side toes were present but not primary weight-bearers) and hoofed, with taller grinding crowns and deep jaws. These were larger than earlier horses, but not yet the size of modern equids
- The late Miocene-Pliocene radiation of grazing horses resulted in lineages that include today's monodactyl grazers
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
It is worth bringing up an issue of the geography in the mid-Cenozoic:
{kind=link}
- North America, Eurasia, and Africa were all connected into a "World Continent", and had a common shared fauna. Prominent large-bodied members include the ungulates (perissodactyls, artiodactyls), proboscideans (elephants), carnivorans (the group that contains cats, dogs, and bears)
- As it is today, Australia and New Guinea were isolated from the rest of the world and its mammalian fauna was dominated by marsupials
- Similarly Madagascar and New Zealand have their distinctive unique and isolated faunas. Neither were dominated by grasslands, though, so aren't really of concern here
- Unlike today, South America was also isolated (until around 6-8 Ma). Its mammalian carnivores were mostly marsupial-relatives (but large birds were among the apex predators!); its herbivores included a native radiation of stem-perissodactyls (the SANU: South American Native Ungulates) and the xenarthrans (the sloth/armadillo/anteater group)
- The isolation of South America ended after 6 Ma, when the rise of the Isthmus of Panama allowed for the Great American Biotic Interchange (or GABI). Some South American mammals (including ground sloths, armadillos, porcupines, and opossums) headed north, and many World Continent mammals (including proboscideans, horses, tapirs, camelids, cats, dogs, peccaries, and eventually humans) headed south. Unfortunately the native South American fauna got the worse of this exchange, and many groups died out.
- Antarctica seems to have had a shared fauna with South America in the Paleocene and Oligocene, but it had iced over in the Oligocene and so didn't have a real distinctive land mammalian fauna
The Ice Age Cometh
Now, a little stratigraphy:
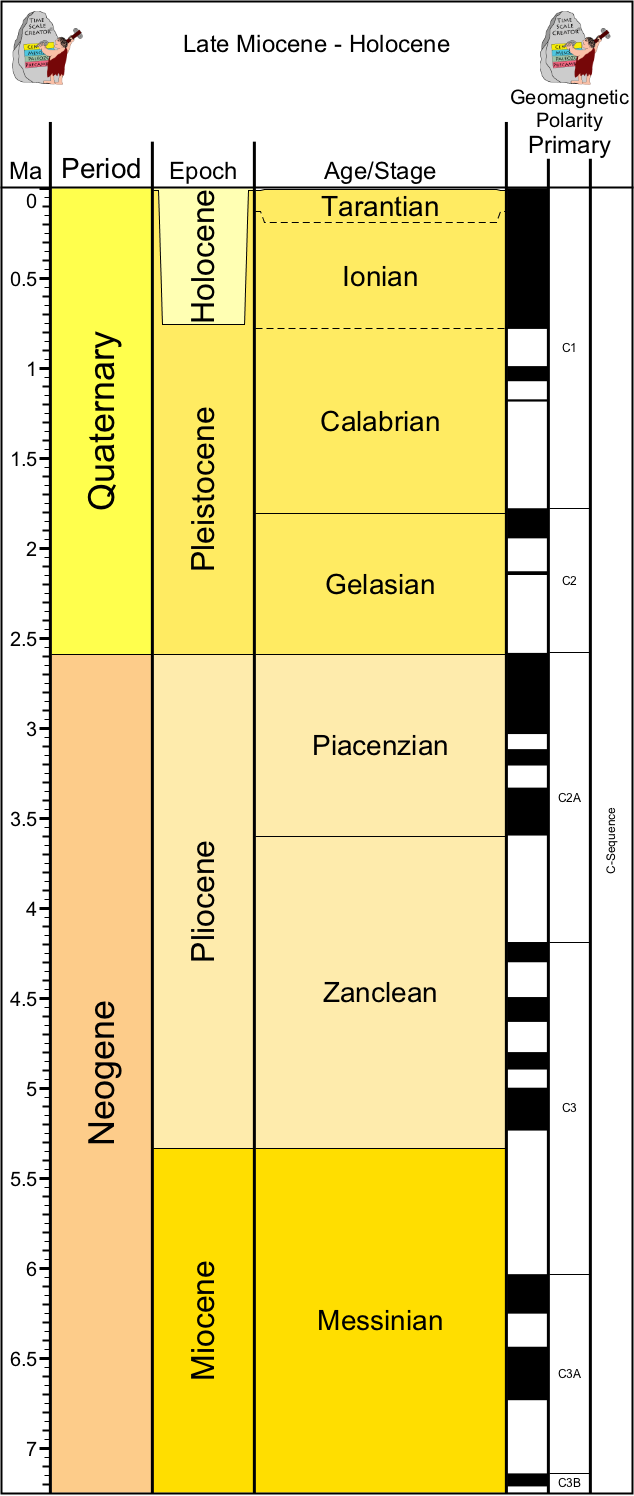
The most recent Period of Earth History is the Quaternary. Based on our modern stratigraphy, it began at 2.588 Ma with a shift from an Icehouse to an Ice Age world, and the start of the glacial-interglacial cycles. The time between the onset of the glaciation and the most recent deglaciation is the Pleistocene Epoch, and the time of the recent interglacial (including this moment right now, and now, and now...) is the Holocene Epoch. Overall, Quaternary climates are cold by Earth History standards: even the interglacials of the Pleistocene and Holocene are cooler than the preceding Pliocene and Miocene times, much less the Greenhouse world of the Eocene and earlier.
The Pleistocene has a characteristic fauna. We'll encounter the Pleistocene fauna of many other parts of the world in a future lecture; for this one we will concentrate on boreal (northern) Eurasia. Some of the more famous and dramatic were:
- Mammuthus primigenius (the woolly mammoth: also in North America)
- Coelodonta antiquitatus (the woolly rhino)
- Elasmotherium sibricum (an even larger, single-horned rhino; recently discovered to have been present 26 ka [earlier thought to have died out at 350 ka])
- The giant Irish elk ( Megaloceros giganteus)
- The aurochs (Bos primigenius, the giant wild ancestor of cattle, which persisted into historic times
- The cave lion ( Panthera leo spelaea) a subspecies of the modern lion (or, arguably, Panthera spelaea, a separate species)
- The scimitar-toothed cat (Homotherium latidens, a cousin of the even larger New World saber-toothed cat, which we will encounter later)
- The cave bear ( Ursus spelaeus)
- Cro-Magnon (Homo sapiens), and before them Neanderthals (Homo neanderthalensis) and related species (H. heidelbergensis and H. antecessor): more about these in later lecture;
- As well as the surviving animals of the Arctic North, such as musk ox (Ovibos moschatus and extinct related species), reindeer (Rangifer tarandus), and the Arctic fox (Vulpes lagopus): all of these are also present in Greenland and/or North America.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
These animals survived repeated glacial-interglacial episodes, moving North and South to track the cooler (but not fully glaciated) conditions they preferred.
The Pleistocene saw two major new biomes show up. The tundra is a lowland, generally coastal environment. The vegetation is often descended from mountain species, and are very low to the ground. It dominates the polar regions today. However, during most of the Pleistocene the cold environment was dominated by the mammoth steppe. This was dominated by grasses rather than low flowers, and had much higher diversity of plants (and of animals feeding them). Mammoth steppes had much higher productivity than tundra does. They required mammoths and other large animals to form: these fertilized the steppes with their feces, and exposed soils which allowed for various plant species to colonize and grow.
The boreal mammalian community was famously the Age of the Woollies: animals with thick heavy furs. Many show dry land feeding adaptations (high-crowned teeth, etc.), especially in the mammoth steppes. They often had shorter and stockier limbs than their southerly kin, and larger bodies (and thus having a lower surface area/volume ratio, and thus retained heat better.)
The Third Pole
But where did they come from? Some had origins from the Pliocene species that lived in the same region. For instance, woolly mammoths seem to be the descendants of the steppe mammoth Mammuthus trogontherii , itself descended from the Southern Mammoth Mammuthus meridionalis. Cave lions seem to nested phylogenetically within the lion-like panthers (lions and leopards), which were still living in the temperate zone (even Europe) in early historic times. And so forth.
{kind=link}
{kind=link}
But a new set of discoveries were made in the Tibetan Plateau in the last decade. The vast elevated Tibetan Plateau is sometimes called the "Third Pole". The conditions there are similar to polar regions due to the extreme cold.
A series of joint US/Chinese expeditions in the Zanda Basin have explored lake deposits containing abundant mammal fossil remains. These range from the late Miocene through the Pleistocene. They document several important discoveries:
%20reduced_1611717a-d910-4ece-a3b3-d3a0a5db1865.jpg){kind=link}
- The Pliocene Epoch Tibetan woolly rhino Coelodonta thibetana, the ancestor of the woolly rhino which spread out of Tibet to colonize the North with the onset of the Ice Age
- The ancestor of the Arctic fox shows a similar pattern
{kind=link}
In fact, there are some animals from the Zanda Basin fossil record whose descendants were to spread to other parts of the world, such as:
- Panthera blytheae, the oldest and basalmost species of the big cats, which spread both north and south
- <Protovis himalayensis, the oldest and basalmost member of the sheep group (many of which are mountain dwellers, even today)
- Sinicuon, the oldest of the clade of hypercarnivorous dogs that survive today as Cuon or dhole (a predatory dog of Asia, that in the Pleistocene was also found in Europe and North America) and Lycaon or African painted dog)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
The "Out of Tibet"/"Third Pole to the North Pole" hypothesis is compelling: after all, animals adapted to the cold Tibetan Plateau would do fine when the world began to chill. The Zanda Basin expeditions are on-going, so we will see what other Pliocene precursors of Pleistocene mammals may be discovered.
But what of Cro-Magnons, Neanderthals, and the like? Their story takes us back not to Asia, but to Africa.
Some Relevant Videos