

The science of fossil shells is the first step towards the study of the earth.
-- Conchiologia Fossile Subappennina, Vol. 1 (1814), by Giovanni Battista Brocchi (1772-1826), early Italian geoscientist
BIG QUESTION: How did the marine life of the Paleozoic Era change over time?
Patterns of Biodiversity in the Paleozoic Marine Realm
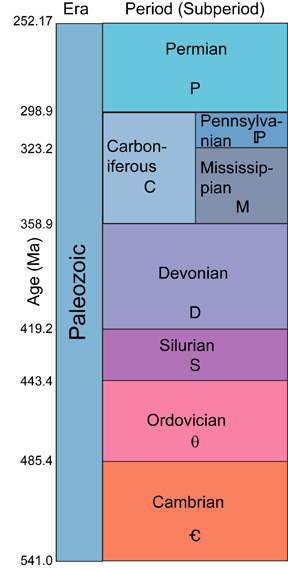
For reference, here is the geologic timescale for the Paleozoic Era:
In the 1981, paleontologist Jack Sepkoski used one of the first large-scale computer analysis of marine biodiversity through the fossil record. In it, he discovered that certain groups of organisms shared similar patterns through the Phanerozoic: if one became common, all became common; if one became rare, the others became rare as well. He termed these the evolutionary faunas, and found that most of the members shared similar suites of ecologies. These evolutionary faunas are the Cambrian, Paleozoic, and Modern faunas. (Don't let the names fool you! The "Cambrian fauna" is still present, but very rare; the Paleozoic fauna survives at moderate levels, and the Modern fauna goes back all the way to the Cambrian.)
The groups in each of the evolutionary faunas tend to share similar types of general traits:
In our examination of Paleozoic biological history, we'll take a look at certain major patterns:
Plus, we'll highlight a few major groups of marine life along the way.
The Cambrian Substrate Revolution (aka the Agronomic Revolution): Increasing grazing and increasing burrowing doomed the algal mat community. (Indeed, thick biofilms only grow today in places where metazoans are excluded). While the sediment-water interface was once firm and the nutrients in the sediment once trapped and excluded from the water, the Cambrian Substrate Revolution churned up the sediment, allowing for soupier sediment-water interface and more availability of the sediment nutrients into the water and vice versa.
A major group of fossil animals in the Cambrian (and persisting for the whole of the Mesozoic) were Trilobita, the trilobites. They were one clade within the Arthropoda, the "bugs" (broadly defined). Their closest living relatives are the chelicerates, which include horseshoe crabs (Xiphosura) and arachnids (Arachnida: spiders, scorpions, mites, ticks, daddy longlegs, and a bunch of groups you probably have never heard of) as well as the extinct water scorpions (Eurypterida). More distant relatives are the Mandibulata, a clade including the living Myriapoda (centipedes and millipedes) and Pancrustacea, which itself include the many branches of crustaceans (a paraphyletic grade) and the Hexapoda (insects and their kin).
More than 20,000 species of trilobite are known, distributed among over 5000 genera are known. (In other words, we know of more species of trilobite than of modern and extinct mammals combined.) All have some variation of their basic three-lobed body plan. The three lobes in particular are the central axial lobe (which contains the main part of the body) and the right and left pleural lobes (extensions of the body that cover the legs and gills). Trilobites are also famously divided into three sections from front to back: a cephalon (head), a thorax, and a pygidium ("trilo-butt"). The entire top surface of the trilobite is not just chitin: instead, this surface is made of calcite (including the eyes), making the carapace of a trilobite wonderfully preservable.
Given their large number of species, there wasn't just one way of life among trilobites. A wide variety of life habits are found in Trilobita:
The main reef builders of the Cambrian were archaeocyathid sponges (in earlier Cambrian only: almost no middle or later Cambrian reefs). Like some other sponges, they produced an internal calcareous skeleton.
Following the Cambrian is the Ordovician Period, noteworthy for the Great Ordovician Biodiversification Event (GOBE). During the Ordovician the Cambrian EF remains high, but also saw the vast increase in the diversity of the Paleozoic EF taxa. This saw the migration of animals away from the sediment-water interface, and the rise of far more predation. A side effect of this was the migration of some of the Cambrian EF into deeper water from the shallows.
Some of the noteworthy new groups of the Ordovician:
These, and many other taxa, resulted in a tremendous increase in the diversity of marine life. But the Orodvician was to end with the first, and one of thee most extreme, mass extinction events.
The Ordovician/Silurian Mass Extinction (443.8 Ma): Perhaps 85% of species are lost (although the total number is less than later ones: there were fewer total species known for the Ordovician than for later in the Phanerozoic.) There is devastation within brachiopods, bryozoans, graptolites, trilobites, and conodonts, but no major clades were lost. The Cambrian EF would never recover, and remained only a minor component of later biodiversity. A fair amount of evidence suggests there were two pulses of extinction, separated by about 1 Myr.
Despite some news items in the 2000s suggesting a "hypernova" or "gamma ray burster" (extremely huge supernova) explosion being the cause, there is zero (zip, nada, no) evidence for this. Instead, this event coincides with a major period of glaciation. It has been suggested that this might have been responsible for the extinction by perhaps reducing the available habitat area in the shallow marine realm (because of a major sea-level drop), and/or a major period of anoxia from global eutrophication, due to the influx of considerable amount of nutrients from material scraped off the continents from glaciers (or from the first communities of primitive land plants).
The world recovered, and during the Silurian and Devonian periods the level biodiversity equalled or exceeded that of the peak of the GOBE. One main factor was the rise of the tabulate-stromatoporoid reefs. The Silurian and Devonian were the greatest reef-building period in Earth's history: more reef material was produced then than before or since. And just as today, a big reef is more than the animals that build it: it is also the home to vast levels of biodiversity
The main reef organisms included:
The new apex predators of the Silurian were the Eurypterida (sea scorpions or water scorpions) (Ordovician-Permian). This group of arthropods produced some 3-m long giants (the largest and most massive arthropods in Earth history.) Some of these could come onto land for short periods of time, and one clade became entirely terrestrial in the Carboniferous (and were therefore "land sea scorpions"). Despite their name, they were not scorpions or arachnids, but were stem-arachnids.
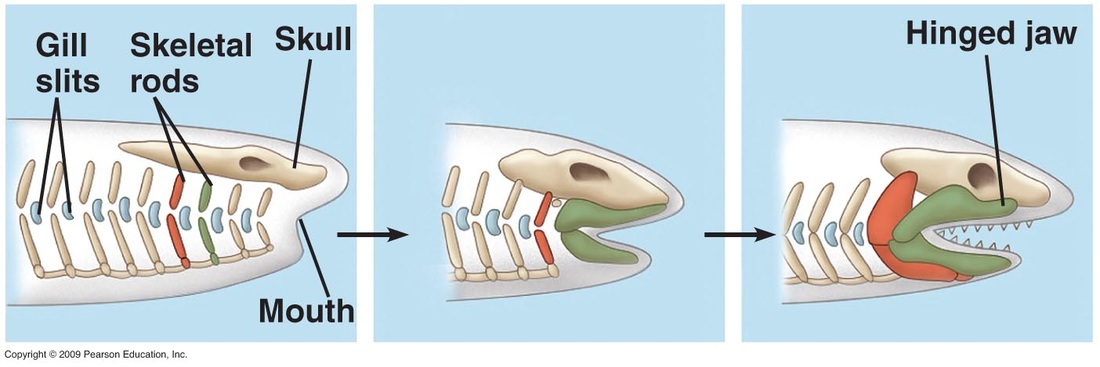
Silurian "ostracoderms" evolved paired fins, which increased their maneuverability. But much more important in vertebrate history was the evolution of jaws. Apparently derived from gill supports, the rise of jaws allowed for substantial changes in the mode of life of early vertebrates. Previously the gills served both to capture oxygen and capture food, and thus couldn't be simultaneously optimized for both. So "ostracoderm" size was small. In contrast, the origin of jaws allowed fish to capture food of various sizes and consistencies, and allowed the gills to become better at exchanging gases.
The clade of jawed vertebrates is called Gnathostomata ("jawed mouths"), and the crown-clade is Eugnathostomata ("true jawed mouths"). The two living clades are Chondrichhtyes (the "cartilaginous fish": sharks and rays, ratfish, and their extinct king) and Osteichthyes (the "bony vertebrates", including all the land vertebrates). The non-eugnathostome gnathostomes are the paraphyletic "placoderms": jawed, shelled, toothless fish. Chondrichtyians, osteichthyians, and "placoderms" were all present by the end of the Silurian, but they would undergo their main diversification in the Devonian.
Osteichthyians are notable not that they had bone, but rather that their bone was used in their endoskeleton and not simply as heavy scales. Additionally, osteichthyians ancestrally evolved lungs, to assist in getting oxygen by gulping it from the air. Their increased mineral storage (bones inside their body) and ability to gulp air suggests that osteichthyian evolution was strongly influenced by living in fresh water, which lacks salts (duh!!) and in which anoxia is more possible than in the shallow seas. But that story is for a later lecture. Silurian osteichthyians were stem forms, but during the Devonian the clade split into its two diverse surviving groups, the ray-finned Actinopterygii and the lobe-finned (and eventually legged) Sacropterygii.
During the Devonian Period, a new major cephalopod group arose: the Ammonoidea. This group of spiral-shelled cephalopods had complex suture lines (the contact between the septum separating the different chambers of the shell and the inner wall of the shell). Although obscured from the outside view in a living animal (or a complete fossil), the walls of cephalopod shell fossils often wear away, allowing you to see the suture line. In nautiloid-grade forms the suture line is a relative simple curve, where as it becomes more and more complex in various forms of ammonoids. (The "goniatitic", "ceratitic", and "ammonitic" suture lines are all found in Ammonoidea.) The function of the increased complexity of the suture has been the subject of considerable research. In general it seems secure that this increase (and with it, increased points of contact between septum and shell) would increase the mechanical strength of the shell overall, but what specific adaptive circumstance (speed? depth of diving? size? something else?) is being selected for remains uncertain. Ancestrally ammonoids were predatory, eating other cephalopods (even other ammonoids), fish, and other animals.
Also during the Devonian, the "placoderms" diversified into many ecological niches. One of the most important parts of this radiation was the rise of the Arthrodira. Arthrodires were fast swimming predatory forms, and the largest ones (such as 4-5-m long, great white shark-like Dunkleosteus) were the apex predators of the period. A limitation of this clade, however, was the lack of teeth. Although their powerful skulls had slicing blades, these were blades of bone: if they broke them on the shell of a victim, that would be life-threatening injury.
Consequently, the rise of teeth as an attribute of Eugnathostomata proved to be successful. Teeth (modified from armored scales) could be produced in great numbers, have various shapes for different functions, and could be lost and replaced throughout life.
(Incidentally, rivaling Dunkleosteus was weak-jawed Titanichthys. Paralleling what we have seen before (e.g., suspension-feeding giant radiodonts) and will see later (filter-feeding actinopterygians in the Mesozoic; filter-feeding giant rays and sharks in the Cenozoic; plankton-gulping giant whales in the late Cenozoic), this was a filter-feeding giant arthrodire.)
During the Devonian small chrondrichthyians and actinopterygians became common, but their main diversification would be in the Carboniferous. Sarcopterygian diversity also increased, but their greatest success in the Devonian would be in freshwater and later on land.
The increase in cephalopod (esp. ammonoid) and fish diversity in the Devonian is called the Devonian Nekton Revolution. Previously the nektonic taxa were mostly demersal: that is, they swam, but mostly lived and fed right above the sediment-water interface. But from the Devonian onward animals began to occupy more and more of the water column, all they way up to the water-air interface. Now the entire sunlit water column was occupied by life. A side effect of the increase of nekton was the greater transfer of fecal material and carcasses into the deep sea, as swimmers a) move up and down in the water column during diurnal migration, and b) they live farther and farther from shore. So the Devonian Nekton Revolution also increased the diversity of life in the deep ocean.
An ecological response to the Devonian Nekton Revolution was seen in trilobite defenses. There is an increase in more advanced eyes, greater spines, and especially a larger pygidium (allowing for enrollment): all of these traits suggest that defense against increasingly sophisticated predators might have been major factors in trilobite evolution.
As we will see in the next lecture, the colonization of land underwent a great increase during the Devonian. A dramatic side effect of this was another mass extinction event (or is it really two?)
A two-phase event: a first strong one between the Frasnian and Fammennian Stages of the Late Devonian Epoch (also called the "Kellwasser Event"), and the final Devonian/Carboniferous event (also called the "Hangenberg Event"). Some studies once supported a single major terminal Devonian event, with the Frasnian/Fammennian event as a result of the Signor-Lipps effect, but more recent work shows the double phase. This is characterized by a collapse of major reef community (the tabulate-stromatoporoid reef community); as today, reefs in the mid-Paleozoic were MAJOR centers of biodiversity, so collapse of the reef community dragged numerous other species with it. Additionally, there are losses of primitive grades and clades of fish ("ostracoderms", "placoderms", and acanthodians). There are also major extinctions within trilobites (again), eurypterids, echinoderms (with some major clade losses), conodonts (again), bryozoans (again), and ammonoids (for the first but not the last time). Many of these survived with only a few handful of species to repopulate the Carboniferous.
Although Otto Schindewolf proposed an asteroid impact as the cause for this (without any direct evidence), this is currently best explained as a result of the spread of vascular land plants! With the first rain forest and the first seed plants in the continental interior, there was considerable burial of excess carbon (leading to cooler temperatures and reduced continental shelf space). More importantly, increased mechanical and chemical weathering on the continents leads to increased nutrients into the sea leading to marine phytoplankton booms leading to eutrophication and anoxia. (Indeed, modern coral reefs are suffering from these effects locally because of increased runoff due to human activities in these regions.)
Unusual for a mass extinction, this/these extinction(s) did not really increase the rate of extinction. Instead, they suppressed the rate of origination of new species. Why this should be is not well understood.
In the wake of the Late Devonian Mass Extinction(s), only a single clade of trilobites were left, and the tabulate-stromatoporoid reefs were no more (although Tabulata persisted.) The "placoderms" were lost, but early in the Carboniferous the eugnathstomes (mostly chondrichthyians, but also actinopterygians and marine sarcopterygians) replaced their niches. In the role of apex predators for the Carboniferous and Permian were the edestoid chondrichthyians, including 6.7-m saw-jawed Edestus and the 7.6-m buzzsaw ratfish Helicoprion.
There were geochemical changes in the oceans that modified its diversity. There was no real replacement for the lost tabulate-stromatoporoid reefs, and in fact in the Early Carboniferous the rate of seafloor spreading decreased, switching the oceans from the calcite seas that they had been in since the early part of the Cambrian back to aragonite seas (in which massive calcite structures were harder to produce). Instead of reefs, "crinoid meadows" (really "pelmatozoan meadows", since blastoids were in them) and tracts of lacy bryozoans were common.
In the Permian some reefs came back. Instead of corals, the main framework builders were a mix of aberrant brachiopods, bryozoans, sponges, and calcareous algae. While not common, some Permian reefs are spectacularly well preserved.
Although not one of Sepkoski's "Big 5", it turns out that the third worst mass extinction event was hiding at the boundary between the Middle and Late Permian. Called the Guadalupian Mass Extinction or Capitanian Mass Extinction, it rates as one of the most severe in terms of species loss. It coincides with the eruption of the Emeishan Traps eruptions in China, which likely were the causal agent.
Diverse terrestrial animals and plants were lost, as well as the giant Paleozoic forms and many brachiopod clades. The Permian reef community collapsed, and the "Paleozoic"-style ammonoids die out, leaving only the clade that would persist and thrive in the Mesozoic.
However, far worse was on the horizon...

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}