"Why then is not every geological formation and every stratum full of such intermediate links? Geology assuredly does not reveal any such finely-graduated organic chain; and this, perhaps, is the most obvious and serious objection which can be urged against the theory." -- Chapter 9 "On the Imperfection of the Geological Record", On the Origin of Species by Means of Natural Selection (1859), Charles Darwin
"Since we proposed punctuated equilibria to explain trends, it is infuriating to be quoted again and again by creationists -- whether through design or stupidity, I do not know -- as admitting that the fossil record includes no transitional forms. The punctuations occur at the level of species; directional trends (on the staircase model) are rife at the higher level of transitions within major groups." --The Panda's Thumb (1980), Stephen Jay Gould
BIG QUESTION:How do new species form?
Speciation: The Pattern of the Origin of Species
As with many things, we run into problem with typological thinking: the idea that there are ideal types of things, and that we judge a specimens membership in a group by how well it conforms from that type. Instead, we find that variation is the reality. So we need to use population-based thinking. (Next lecture we will add tree-based thinking.)
Darwin's species concept is worth revisiting:
"I look at the term species as one arbitrarily given, for the sake of convenience, to a set of individuals closely resembling each other, and that it does not essentially differ from the term variety, which is given to less distinct and more fluctuating forms. The term variety, again, in comparison with mere individual differences, is also applied arbitrarily, for convenience sake." Chap. 2 of The Origin
"Species are only well-marked varieties" Chap. 4 of The Origin
"The only distinction between species and well-marked varieties is, that the later are known, or believed, to be connected at the present day by intermediate gradations, whereas species were formerly thus connected" Chap. 14 of The Origin
An important issue which is commonly forgotten comes out here: descendants are descendants of only a small part of any ancestral group! That is, entire species do not evolve into entire other species. Instead, only some small subset of any given species population is the ancestral group leading to a particular descendant. This points to several different aspects:
It is entirely possible for the ancestral species to still be around even though some subpopulation might give rise to a later species
There is no reason why for a given ancestral group that only a single subpopulation gives rise to descendants. Thus, we have a reason for divergence of multiple descendants from a single ancestral species
Physical geographic isolation of a subgroup and a change of environmental condition favoring evolution of distinct traits would greatly increase the chance in some small subgroup on the fringes (a peripheral isolate) to be most easily split off as a potential ancestor of a new species
Note: this relates to a common anti-evolutionary rant, which goes a long the lines of "if people evolved from monkeys, how come there are still monkeys". Ignoring lots of other problems with this statement (such as the fact humans didn't evolve from any living monkey species; that "monkeys" aren't one thing, but are a vast number of species; etc.), this misses important aspects of how evolution works! Just because some monkeys evolved into apes which evolved into humans does not require that ALL monkeys evolved into apes and ALL apes evolved into humans. Plus, it doesn't mean that monkeys were TRYING to evolve into humans, or DESTINED to do so.
(Here's a way to restate an analogy to this anti-evolution argument: "If [for example] your ancestor came from Ireland (or Norway, or India, or whatever), why are there still Irish/Norwegians/Indians/etc.?")
Adaptive (or Fitness) Landscapes
One metaphorical device that is sometimes useful to use when talking about evolution is the idea of an "adaptive landscape" or "fitness landscape". Imagine all the variables in an organism's circumstances reduced to a surface with peaks and valleys. The height of the peak reflects the fitness of the organism. Evolution will always favor populations moving "uphill" (increasing fitness). But they can only move to the nearest peaks through ordinary selection, even if there are higher peaks elsewhere. That is because moving downhill would mean a decrease in the fitness of the descendant populations. So selection only moves towards "local" optima in normal situations.
However, large mutational "jumps" might place a descendant on a point on the landscape far from its ancestor, allowing it to move to adaptive peaks not accessible to the earlier forms. And the landscape itself "shifts", because the environment in which the organisms exist change, meaning what controls "fitness" will be different over time.
Speciation is the process of the origin of a species. It doesn't happen immediately or instantaneously: it is indeed a process rather than an instantaneous event. (In fact, except in rare cases, it is unlikely that it you there during it that you would recognize it as such.)
Some aspects of the origin of species to consider:
Area of origin: Did the new species arise within the main range of the ancestral species (sympatric, "same homeland"); alongside the ancestral range, with no major barriers to gene flow (parapatric, "parallel homeland"); on the edges of the species range, with some substantial (but not necessarily total) barrier to gene flow (peripatric, "edge of the homeland"); or by either subdividing the original population or by isolating a part of it (allopatric, "other homeland")?
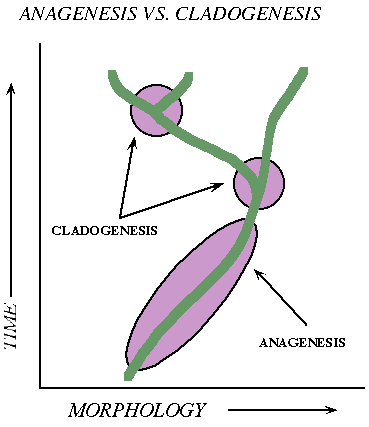
Trends or branches: Did the the new species arise by the main population itself shifting as a group (anagenesis, "no origin") or by splitting/subdivision of the lineage (cladogenesis, "branching origin")
During the 20th Century (especially during the first half), evolutionary biologists assumed the dominant trends were sympatry and anagenesis. However, as a better understanding of genetics was developed, some (including Mayr) argued that allopatry, peripatry, and parapatry (which all require cladogenesis) were actually more common.
The problem, of course, is that speciation takes time, and field biologists are unlikely to observe it. If only there were some sort of record of changes over time. Say, for example, a fossil record...
Creeps or Jerks: Phyletic Gradualism vs. Punctuated Equilibrium
Prior to the 1970s most paleontologists considered a model which became known as phyletic gradualism:
Speciation is mostly sympatric: change occurs mostly within the ancestral range
Speciation is mostly anagenetic: the whole population changes together
Evolutionary change is largely continuous
As a consequence, fossil species were all simply chronospecies: arbitrary sections of a lineage with no particular justification for dividing the lineage up one way or the other
Speciation is mostly para/peri/allopatric: change occurs mostly by peripheral isolation
Speciation is very dominantly cladogenic: branching (either subdivision of the whole population, or the origin of a peripheral isolate) is the most common
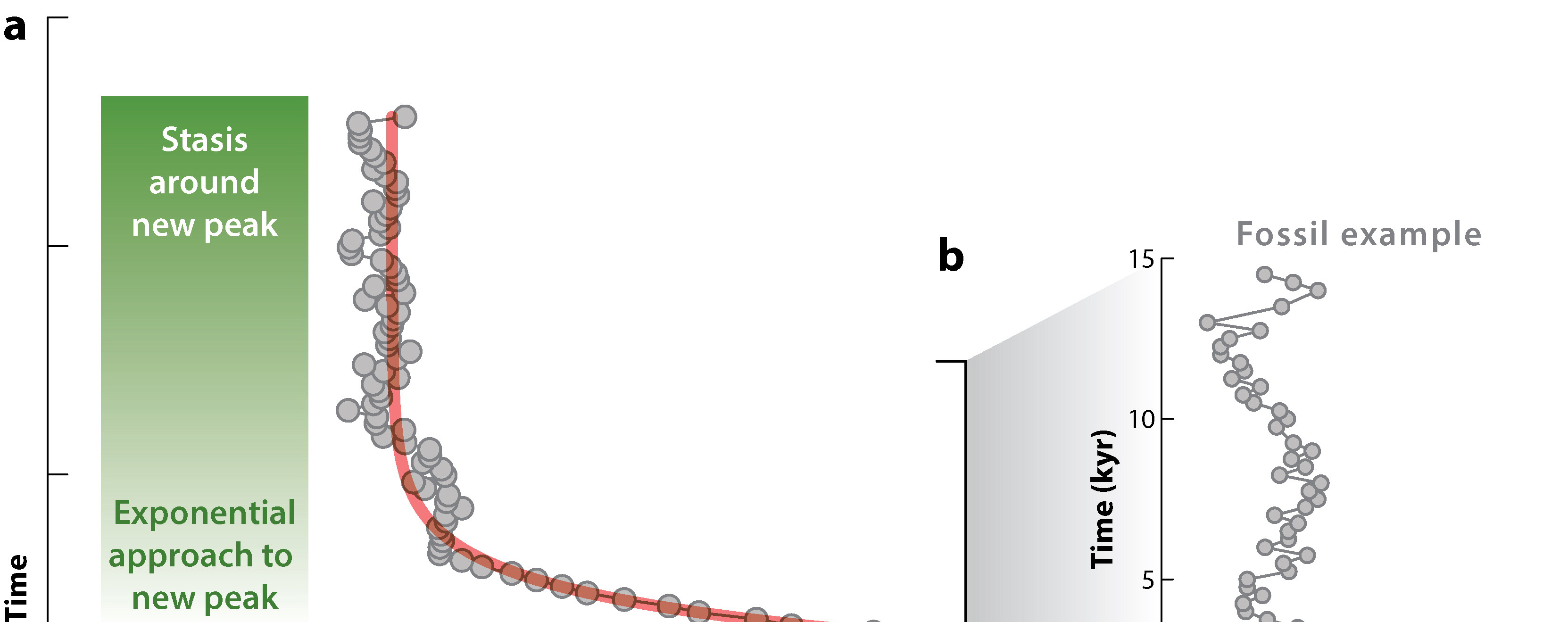
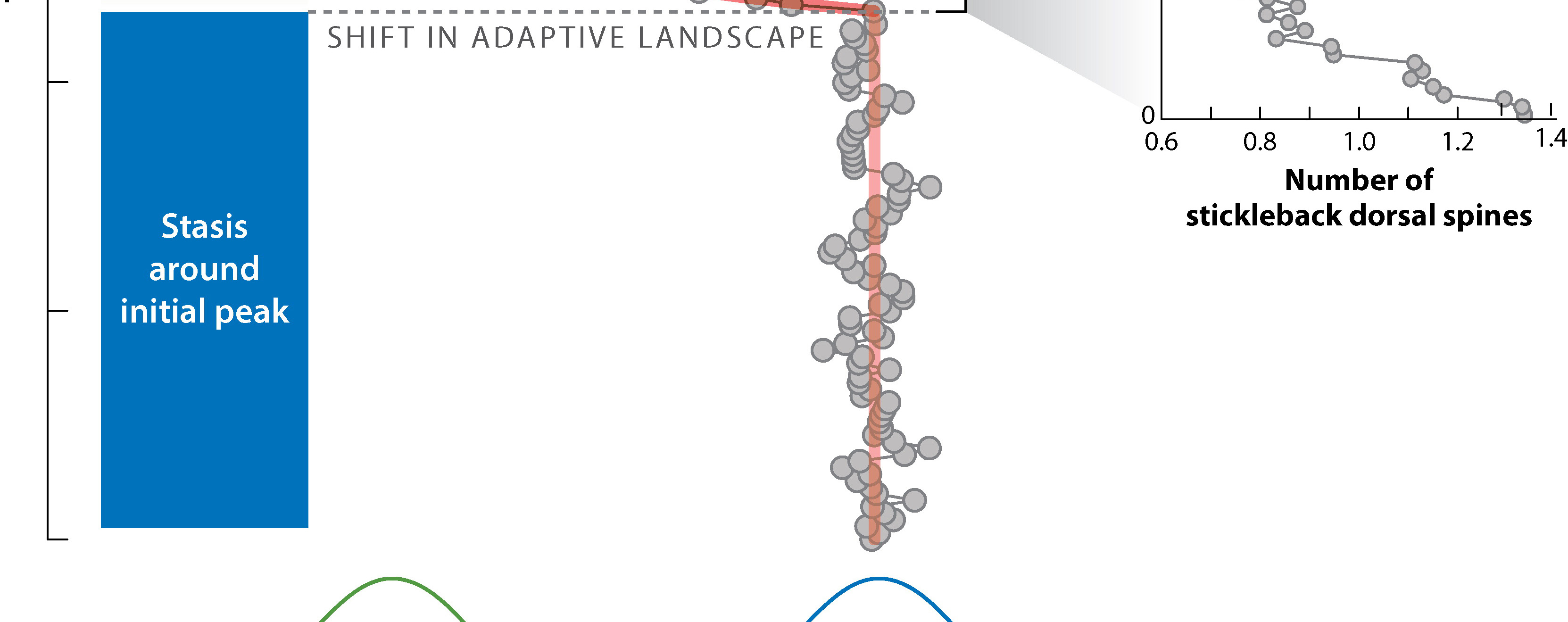
Evolutionary change is not continuous: instead, during most of the history of a species it remains relatively stable (no major directional change) (the "equilibrium" part), and most of the change is restricted to a small portion of its history (the punctuation event)
As a consequence, fossil species have a fairly distinct beginning (the punctuation)
The punctuated equilibrium model helped explain some aspects of paleontology. As Darwin noted (see quote at top), we do not see an endless series of slight gradations, each stratum with a slightly different version. Instead, species remain largely unchanged for most of their duration, with new closely-related species appearing suddenly in the fossil record. In fact, if it weren't like this, biostratigraphy would not really work! As the punctuated equilibrium supporters argued, "stasis is data".
During the 1970s and 1980s (and continuing today, but at with much less rancor), the debate over "evolution by creeps" vs. "evolution by jerks" continued. At least in the fossil record, punctuated equilibrium seems
How long are punctuation events? In a rare case, Smithsonian paleontologists Gene Hunt, Michael Bell, and Matthew Travis found that a population of the stickleback species Gasterosteus doryssus got isolated in a lake in Nevada in the Miocene Epoch. In this particular case, there were annual layers, allowing them to measure an excellent sample over time and document its change. They found the period of transition from the ancestral form to the descendent took only about 2000 generations (about 4000 years), after which the population was mostly stable. Events on the 103 year scale are unlikely to show up in the fossil record except in such situations (high sample size, restricted location, annual record), as the fossil record is much better at picking up events at the scale of 104, 105, or greater.
Supporters of the punctuated equilibrium model had to wonder how equilibrium was maintained. Evolutionary stable scenarios seem to be at least part of the reason.
Why the punctuations? A likely cause is that environmental changes are rather quick on the geologic time scale, with stable conditions in between. Rapid shifts in climates will result in shifting population ranges, shifts in habitat availability, etc. This leads to the prediction that we should see evolutionary shifts (speciations, extinctions, etc.) concentrated at moments of environment change: the so-called "Turnover-Pulse Model".
In summary, punctuated equilibrium may well be due to the following combination of aspects:
Abrupt environmental change, resulting in shifting selective conditions
Evo-devo-type evolution, allowing for a few genetic changes to result in more significant changes of form
Evolutionary stable scenarios maintaining equilibrium between punctuation events
A historical note: a close read of The Origin shows that Darwin did consider cladogenesis and parapatry/peripatry as critically important in most speciation, and that anagenesis of the main part of the ancestral population was almost never the case.
"Missing Links" As An Outdated Concept
One of the most problematic terms in popular discussion of evolution is "missing link". Coined by Lyell, it was used in the 19th Century for a potential but as-of-then undiscovered link between humans and other primates. Subsequently is has been used as the idea of an as-yet undiscovered intermediate form between two particular species.
The term is problematic for a couple reasons:
Once you actually find the specimen, it isn't missing any more!
More importantly, we are unlikely to find any individual that is a direct ancestor of the later forms
The chances that any individual making it in to the fossil record is very, very small. And even the chance that every population, or even species, will show up in the fossil record is small.
An ancestor is a very specific thing: not merely an earlier relative, but specifically an individual who was in direct line of descent with the later form. (Your great great great great uncle is NOT your ancestor, while your great great grandfather is.)
So it has a very small chance that given fossils are the ancestors of the species you are interested in. However, it might well be a more general early relative, and that could be useful indeed.
Transitional Forms
Finding a direct ancestor might be hard, but finding an early relative is easier. And since relatives share many of their same traits, finding early relatives can help establish the anatomical (and ecological, and so forth) transitions involved in that part of the tree of life.
Darwin pointed out that there is not the continuous series of transitions that might expected from a gradualistic model of evolution in the fossil record, but noted that the fossil record was great for higher-level transitions. And this record is vastly better now than in the 1850s!
Here are a handful of interesting transitions recorded in the fossil record:
The various taxa in there might not be direct ancestors of the later branches, but they do show the set of traits associated with a given phase of that evolutionary transition
You don't always have to look at the whole organism: you can concentrate on the transitional states of a particular part of the body
Although not shown here, you can actually use contemporary organisms to look at the transitional phases of a particular organ system or behavior or whatever, with the understanding that you are not looking at the actual ancestors.
"Intermediate forms" is another related term used in the field. Basically, however, every taxon is intermediate between its closest relative and the groups more distantly related.
An important thing to revisit before we move on: the specter of typological thinking. Our minds like to think of discrete types of things. However, when dealing with evolution, there is a continuum of form from one to another. Remember: at no time did a mother of one species give birth to a daughter of another species! It is only from a distance in time do we see the accumulation of changes.
This applies for groups above the species as well. At not moment in the history of life would you witness a population of one major group giving rise to a population belonging to another major group. It would always look just like ordinary speciation. It is only from a distance that we see the Tree of Life.
Macroevolutionary Patterns Macroevolution is the term for evolution above the level of the species. In a real sense, it is the summed effect of multiple rounds of microevolutionary (below the species level) changes. But some patterns only become apparent on the grander scale.
We have already seen correlated progression (the summed affect of adaptions consistent with a particular mode of life) and divergence (the splitting of one ancestral group into two or more distinct descendant lineages) as examples of macroevolution. Here are a few more:
Adaptive Radiations: when many diverse lineages with many modes of life descend from a common ancestral population in a geologically short period of time. This might happen when an ancestral population colonizes a region previously uninhabited (and thus there is no competition), or when a competitor group goes extinct, freeing up ecological niches (ways of life), or when a new adaptation allows an entirely new mode of existence. In nearly all cases adaptive radiations occur when there is a reduction or lack of competition; thus, many different variants among the descendants can be successful.
Niche Incumbency: when one taxon is already adapted to a particular niche, it is difficult for another taxon to slip into that mode of life in the same time and place. After all, the incumbent already has adaptations which allow it to live in that mode, but the new taxon will not yet have acquired those traits. It is hard to compete if you haven't started the race yet! So the means by which one taxon can replace another in the same role is often for some reason beyond sympatric evolution of the new group leading to direct competition with the incumbent, such as:
extinction of the incumbent due to some other reason
or, alternatively, evolution of the new group into that niche in some other region and migration into the incumbent's habitat with all the appropriate adaptations already in place
Convergence: the independent evolution of similar adaptations from dissimilar ancestors. This is almost always the result of the different groups evolving into a similar niche. For instance, tunas, fast-swimming sharks, dolphins, and the extinct ichthyosaurus have a shared torpedo-like body shape, due to independent selection for fast-swimming motion due to their common mode of life (rapid predators).
Exaptation: a very common pattern. The word means "to be fit out of" (just like adaptation means "to be fit towards"). Evolution only rarely works by the addition of brand-new body parts. Instead, normally a part already existing begins to be employed in a new fashion. For some time it might be doing multiple functions, but may eventually the new fashion might take over entirely. For instance, our arms and legs are homologous to the fins of fish. There were early fish that used their fins both to swim and to push themselves through vegetation and over ground (we still have some fish today that do this, like mudskippers and snakeheads). These ancestors of terrestrial vertebrates used these modified flippers for swimming and for walking, but their descendants (including us) use them only for walking. (Well, or flying, or digging, or picking stuff up, or [ironically] in swimming again in whales and the like!)
Heterochrony: literally "different timing". Evolution can happen by shifting the timing of development in ontogeny, so traits that normally developed earlier might develop later, or where development might continue beyond the ancestral version. This is divided up into to major patterns: peramorphosis, where the descendant species evolves into a "hyperadult" relative to the ancestor; and paedomorphosis, where the descendant is a "juvenilized" version of the ancestor.
You can have both patterns showing up in different aspects of the same species. For instance, human heads are paedomorphic compared to our relatives: we have round skulls and flat faces, like juvenile apes but not like adults. On the other hands, our legs are peramorphic: while most apes have legs that stop growing at around the toddler phase, ours keep on growing to become very long.


{kind=link}
{kind=link}
{kind=link}
{kind=link}