


"'Please, friend, let there be no more tears at this parting. Not water tears from you nor crystal tears from me, would that I were able to manufacture them. It's a gesture I envy you. A small but intriguing physical difference.'" -- Arthropod-like alien Ryozenzuzex to his human friends, from Alan Dean Foster's (1982) Nor Crystal Tears
"Trilobites survived for a total of three hundred million years, almost the whole duration of the Palaeozoic era: who are we johnny-come-latelies to label them as either 'primitive' or 'unsuccessful'? Men have so far survived half a per cent as long." -- Richard Fortey, Trilobite: Eyewitnesses to Evolution (2000)
"The careful, embroidered stitches delineated a coil of some sort. It looked rather like a halved snail shell, but the interior was divided into dozen of intricate chambers.
"Is that a nautilus?" he asked.
"Close, but no. It's an ammonite."
"An ammonite? What's an ammonite? Sounds like an Old Testament people overdue for smiting."
"Ammonites are not a biblical people," [Actually, they were! The Ammonites, or "children of Ammon", are mentioned in several passages in the Old Testament -- TRH] she replied in a tone of strained forbearance. "But they have been smited."
"Smote."
With a snap of linen, she shot him a look. "Smote?"
"Grammatically speaking, I think the word you want is 'smote.'"
"Scientifically speaking, the word I want is 'extinct.' Ammonites are extinct. They're only known to us in fossils."
"And bedsheets, apparently." -- Tessa Dare, A Week to Be Wicked (2012)
and
"'Hammonis cornu,' or 'horn of Ammon,' which is among the most sacred stones of Ethiopia, has a golden yellow colour and is shaped like a ram's horn. The stone is guaranteed to ensure without fail dreams that will come true." -- Pliny the Elder, Naturalis Historia (written 77-79 CE), XXXVII.LX
BIG QUESTION: How do the histories of the trilobites and ammonoids compare?
A Pair of Paramount Fossilizers
Trilobites and ammonoids are among the best studied and most famous groups of fossil marine invertebrates. Great numbers of studies have been conducted on each. They have attributes in common that make their respective fossil records so spectacular:
However, there are some significant differences:
Trilobites (technically Trilobita ranged from fairly early in the Cambrian until dying out at the Permo-Triassic mass extinction (so a range from 521-252 Ma). Thus they survived the Ordovician/Silurian and Late Devonian mass extinctions. In some ways they are practically the "poster children of the Paleozoic", coming in with the Cambrian Explosion and perishing with the P/Tr. Ammonoids (technically Ammonoidea) first appear around 409 Ma as part of the Devonian Nekton Revolution. They survive the Late Devonian, P/Tr, and Tr/J mass extinctions, but are wiped out in the K/Pg.
Trilobites have been known to people for as long as we've picked up interesting fossils. Some, such as the Ute Nation of the American Southwest, even recognized them essentially for what they are: the traditional name for them is pachavee ("little water bug"). The first description of one in the scientific literature is by Rev. Edward Lhwyd, who in 1698 detailed a specimen he considered "the skeleton of some flat Fish". The "Dudley locust" or "Dudley bug" is a common species found near Dudley, England. In 1749 Charles Lyttleton described them as Pediculus marinus major trilobos ("large three-lobed marine louse"), and thus gave us the origin of the name "trilobite". (Incidentally, that species is now known as Calymene blumenbachii.)
By the way, a great resource for information about trilobites is Sam Gon III's A Guide to the Orders of Trilobites.
Trilobite is one clade within the Arthropoda, the "bugs" (broadly defined). Their closest living relatives are the chelicerates, which include horseshoe crabs (Xiphosura) and arachnids (Arachnida: spiders, scorpions, mites, ticks, daddy longlegs, and a bunch of groups you probably have never heard of) as well as the extinct water scorpions (Eurypterida). More distant relatives are the Mandibulata, a clade including the living Myriapoda (centipedes and millipedes) and Pancrustacea, which itself include the many branches of crustaceans (a paraphyletic grade) and the Hexapoda (insects and their kin). All arthropods share traits in common, such as:
More than 20,000 species distributed among over 5000 genera are known. (In other words, we know of more species of trilobite than of modern and extinct mammals combined.) All have some variation of their basic three-lobed body plan. The three lobes in particular are the central axial lobe (which contains the main part of the body) and the right and left pleural lobes (extensions of the body that cover the legs and gills). Trilobites are also famously divided into three sections from front to back: a cephalon (head), a thorax, and a pygidium ("trilo-butt"). The entire top surface of the trilobite is not just chitin: instead, this surface is made of calcite (including the eyes), making the carapace of a trilobite wonderfully preservable.
On the underside the only calcified part is the hypostome, a backwards-pointing "jaw" flap. Otherwise, the undersurface doesn't normally preserve. But there are several Lagerstäaut;tten where exceptional preservation allows us to see antennae, legs, gills, and so on. (Linkage to this site does not constitute endorsement for the sale of fossils.)
There are two major branches of Trilobita. Polymerida> represent the vast majority: "classic" trilobites which are typically larger, with many thoracic segments. Polymerids show up in the Cambrian Explosion and survive until the P/Tr extinction. The Agnostida are a group of tiny trilobites with only a few thoracic segments with reduced eyes or even without eyes. They appear shortly after the first polymerids, thrive in the Cambrian and Ordovician, then go extinct at the Ordovician-Silurian Mass extinction. There have been many debates about the agnostids, with some studies suggesting they were not even trilobites, and some suggesting they were nektonic or planktonic. A study from January 2019 confirms them as trilobites, and with life habits similar to the oldest polymerids but specialized for deeper, darker water. Agnostids seem to have arisen by paedomorphosis, as their few thoracic segments and large pygidia are reminiscent of larval polymerids.
Speaking of which, the ontogeny of trilobites is well studied due to the preservation of the different instars. In fact, there are some cases where the recently molted shells have been found.
Most adult trilobites range from about the size of a dime to a computer mouse. However, the largest trilobites were fairly big as an arthropod goes, with only some eurypterids, giant Carboniferous myriapods, and perhaps a few crustaceans being larger.
Trilobite eyes are the subject of much research. Like the rest of the carapace, they too are made of calcite. As in most arthropods, the eyes are compound.
Given their large number of species, there wasn't just one way of life among trilobites. A wide variety of life habits are found in Trilobita:
There is a great number of trace fossils, including some documenting feeding on worms.
Over their history, we tend to see an increase in more advanced eyes, greater spines, and especially a larger pygidium (allowing for enrollment): all of these traits suggest that defense against increasingly sophisticated predators might have been major factors in trilobite evolution.
Trilobites are an example of a clade that peaked in diversity early on, but persisted at lower levels for a long period of time. Their diversity dropped at the Ordovician/Silurian mass extinction, and again at the Late Devonian. Only one clade (Proteida) was actually present in the Carboniferous and Permian Periods. Being benthic organisms with calcareous shells, they were part of the vulnerable groups at the Permo-Triassic mass extinction.
As with trilobites, humans have long known of ammonoid fossils. The Roman writer Pliny the Elder referred to these fossils as cornu Hammonis, or "horns of Ammon", which is the source of our modern word (which was introduced into the scientific literature formally as "ammonite" in 1732 by naturalist John Philip Breyn and as "Ammonoidea" in 1848 by paleontologist Karl Alfred von Zittel). (Ammon is the Romanization of the Egyptian god Amun, whose animal form was a ram. Even in his human form, Ammon had rams' horns on the sides of his head. The typical planispiral corrugated ammonoid shell does in deed resemble rams' horns.)
In Medieval England these fossils reminded people of coiled snakes, and so were called "snakestones". Sometimes people even carved snake heads onto specimens to make them look more "complete". One legend was that St. Hilda petrified and decapitated snakes in her path, resulting in these fossils being common in some regions. Other parts of their world had their own legends: for instance, the Blackfoot nation of the northern Rocky states called them "bison stones", and Medieval Germans as "dragon stones".
In the Renaissance western naturalists first had contact in detail with the living Nautilus (which does not live in Atlantic or Mediterranean waters, but does swim around in the Indo-Pacific.) This allowed these natural philosophers to understand the basic kind of animal that produced ammonoid shells: cephalopods.
"Ammomnoid" or "Ammonite": Outside the technical scientific literature, it is more common to encounter the word "ammonite" than "ammonoid". Is there a difference? Technically "ammonite" refers to either to those taxa with the complex ammonitic suture pattern (see below) or to the clade Ammonitida (the clade containing all Jurassic and Cretaceous ammonoids). "Ammonoid" refers to any member of Ammonoidea, including all ammonites. In general discussion some people refer to all ammonoids collectively as "ammonites" isn't bad, but that is imprecise.
Cephalopoda is one of major clades of Mollusca. Mollusks typically have a shell comprised of an inner layer of aragonite (the nacreous or "mother-of-pearl" layer) and external layers of calcite. The shell is produced by a surface called the mantle. It starts from a tiny protoconch in the larva and expands from this by adding new shell along the outer edge (the aperture). Mollusks ancestrally have a radula or rasping chitinous ribbon, used initially to scrape algae from the substrate. The most speciose groups of mollusk (the "Big Three") are the Gastropoda (whelks and other sea snails, land snails [including land slugs], and sea slugs), the Bivalvia (clams, including cockles, mussels, scallops, oysters, and many many more), and Cephalopoda. (Less common living groups are the Tryblidiida, the Scaphopoda (tusk shells), the Polyplacophora (chitons), and the Aplacophora (shell-less molluscan "worms")).
Cephalopods are ancestrally characterized by having a single conical (and often coiled) shell with is camerate (that is "chambered"): the living fleshy body is only in the most forward ("body") chamber separated from previous chambers by septa (partitions). The earlier camerae are connected to the living body by a specialized structure called the siphuncle: this allows the chambers to be filled with air or fluid, modifying their buoyancy. Today only the nautilids (the living genera Nautilus and Allonautilus) retain external shells; however, ammonoids and many extinct "nautiloids" (a paraphyletic grade) of the Paleozoic and Mesozoic had these shells.
Cephalopods (barring the very earliest forms from the Cambrian) seem to have been nektonic, and the belly foot that characterizes ancestral mollusks (and retrained in gastropods, tryblidiids, and polyplacophorans) and transformed it into arms ("tentacles" in general use, although malacologists [mollusk workers] reserve that term for elongated arms with suckers only near the tip). They are ancestrally predatory, using a powerful beak composed of chitin and proteins to bite into prey.
In the modern world the nautilids are the only shelled clade. The vast majority of cephalopod species are members of Coleoidea, a group which have internalized or lost the shell. This is the group of squids, cuttlefish, and octopuses. An extinct group of coleoid, the belemnoids, may be paraphyletic with respect to coeloids: in these forms, the shell is internalized but still calcified. During the Paleozoic a great number of shelled cephalopods existed: collectively referred to as "nautiloids".
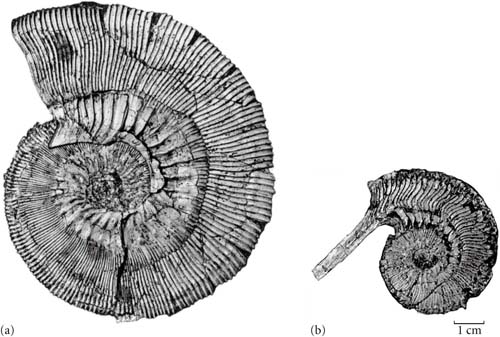
Ammonoids lie phylogenetically between Nautilida and extinct nautiloids on the one side, and coleoids on the other. Like nautiloids they retained an external camerate shell. In most cases this shell is planispiral (wrapped along a single plane, like a cinnamon bun), rather than the trochospiral shell of a gastropod (there the aperture migrates along the central axis during growth). There are, however, almost-straight shelled ammonoids as well as various heteromorph forms (about which we'll see more below.)
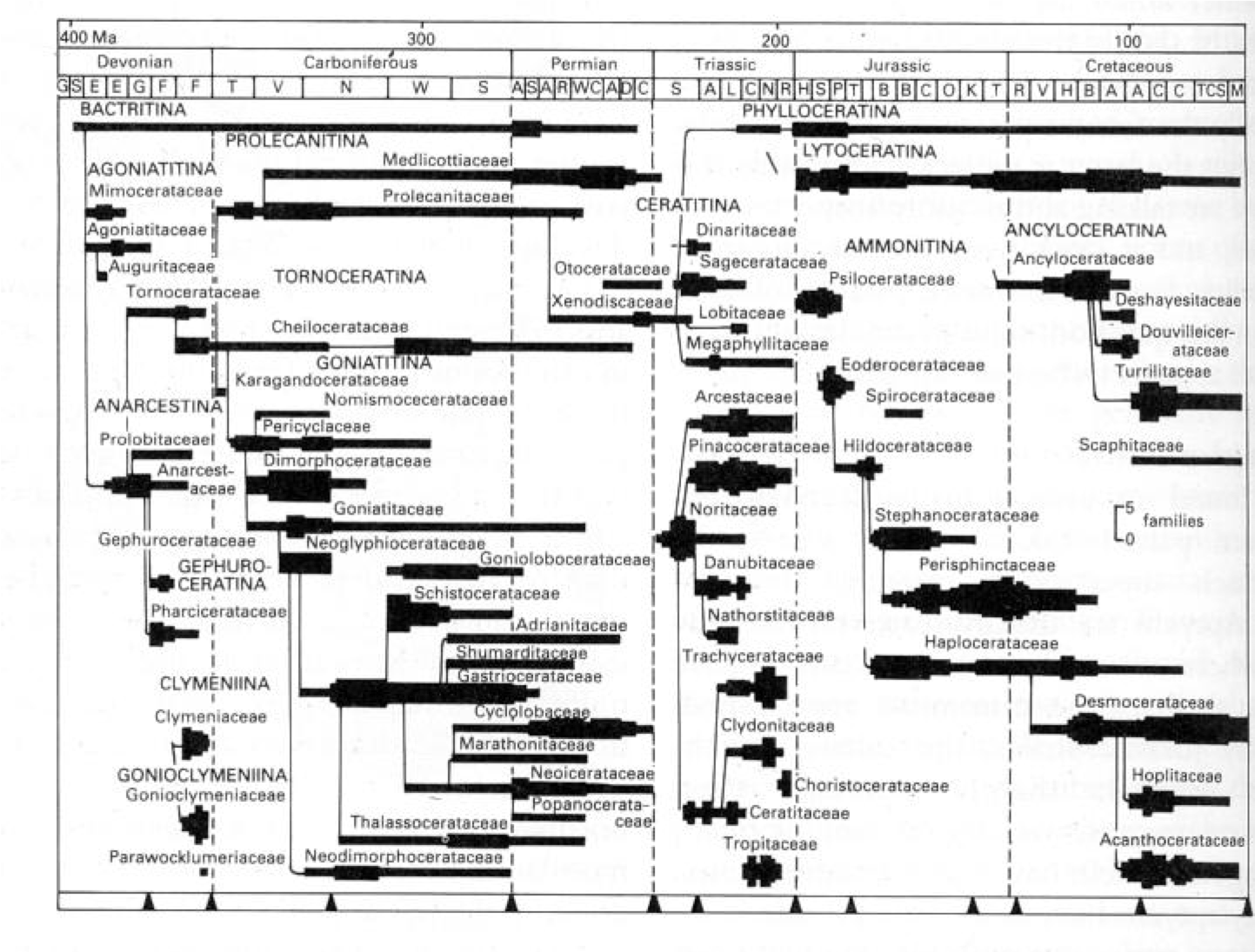
Ammonoidea is one of the most successful of all cephalopod clades, with more than 10,000 known species. They first appear early in the Devonian as part of the Devonian Nekton Revolution, and die out at the K/Pg boundary (indeed, for generations their disappearance was used as the primary indicate that boundary.)
The shells of ammonoids are distinguished from nautiloids by several features. Where as the siphuncle is in the middle of the septum in a nautiloid, it is off to the edge in ammonoids. Also the shape of the margin of the septum differs between the nautiloid-grade and the ammonoids. The suture line is the name given to the contact between the septum and the inner wall of the shell. Although obscured from the outside view in a living animal (or a complete fossil), the walls of cephalopod shell fossils often wear away, allowing you to see the suture line. In nautiloid-grade forms the suture line is a relative simple curve, where as it becomes more and more complex in various forms of ammonoids. (The "goniatitic", "ceratitic", and "ammonitic" suture lines are all found in Ammonoidea.) The function of the increased complexity of the suture has been the subject of considerable research. In general it seems secure that this increase (and with it, increased points of contact between septum and shell) would increase the mechanical strength of the shell overall, but what specific adaptive circumstance (speed? depth of diving? size? something else?) is being selected for remains uncertain.
Another ammonoid trait is the transformation of the lower part of the beak into paired aptychi. These seem to have multiple functions: as a scoop to aid in feeding, and as a cover when the body retracts into the shell (basically, using a giant armored lower lip to protect your insides).
Sadly, the soft tissues of ammonoids remain largely unknown. Thus the size and number of arms they had, whether any of these were tentacles, the complexity of their eyes, and so forth remains undetermined. However, they almost certainly did NOT look like an octopus or squid (with long, muscular arms and tentacles) shoved into an ammonoid shell, as nearly all artists draw! Modern nautilids have a large number of relatively short arms, and belemnoids had ten relatively short arms (often with calcareous or chitinous "claws". Positioned phylogenetically between these clades, it is more likely ammonoids had shorter arms rather than very long muscular ones like most extant coeloids.
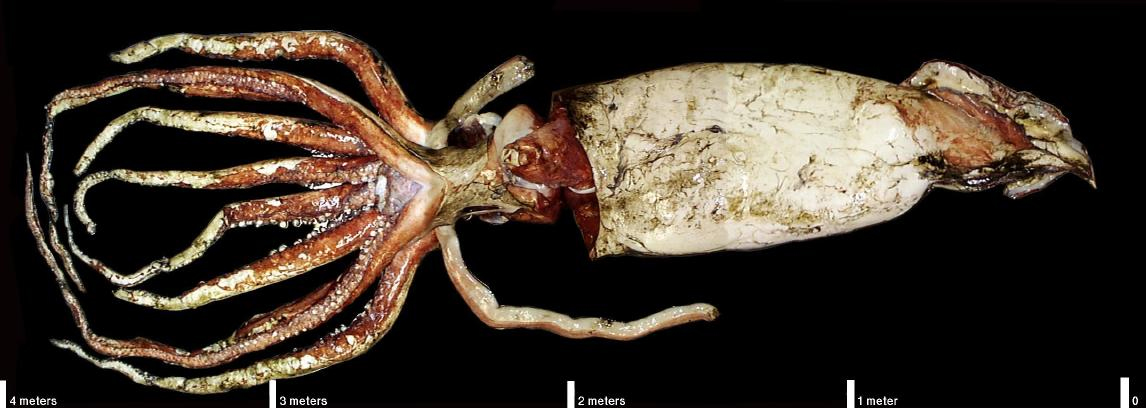
Ammonoids grew throughout their life. Adult body size of most ranged from about the size of an old half-dollar to that of a dinner plate, although finding some about the size of a car tire isn't that rare. The very largest ammonoid was the Late Cretaceous Parapuzosia, with a shell diameter up to perhaps 2.5 m. The living flesh of such an ammonoid would be more massive than a human, and much larger than the living giant octopus (Enteroctopus) or Humboldt squid (Dosidicus gigas), comparable in mass to the extant giant squid (Architeuthis dux), but smaller than the living colossal squid (Mesonychoteuthis hamiltoni) (at present the largest known invertebrate animal in history).
We can infer the life habits of ammonoids based on their shell shape (especially how flattened-vs-wide and smooth-vs-ribbed they are). Drifters had moderately flattened shells and often ribbed surfaces, and may be the ancestral forms. These seem to have eaten small-bodied invertebrates in the water column (although some bite marks on ammonoid shells suggest they may have eaten each other). Demersal forms were technically nektonic, but cruised just above the sea floor. These are often highly ribbed and had quite wide shells, and likely ate benthic epifauna. Nektonic forms with very flattened shells and smooth sides were likely the fastest ammonoids. However, this speed may not indicate they were chasing fast prey: these adaptations may have been to prevent them from being prey. Their relatively small aperture size suggest, in fact, that their arm musculature may have been less well-developed than other ammonoids, and like the heteromorphs they may have been largely planktonivorous. Vertical migrants are morphologically intermediate between the demersal and nektonic, and as the name implies are interpreted to have made daily migrations from dark deeper water to the shallower water at night (as do a great variety of modern plankton and nekton, including the Humboldt squid.)
Finally, there are the heteromorphs, which abandoned the ancestral coiled planisprial shell for a variety of other body plans. A few had relatively straight shells, but most show some form of wild uncoiling. Their shells would be extremely unstreamlined, suggesting they were slow swimmers. They seem to have been abundant in the middle of the water column, and so likely were not feeding on benthic species. They sieve-like shape of their radulae is consistent with them being planktonivores on soft-bodied and/or small prey.
Indeed, the numerical abundance of ammonoids relative to their nautiloid cousins suggests that they fed relatively low on the food chain. It would be hard to support great shoals of ammonoids (as are found as fossils at some sites) if they were rapacious near-apex carnivores like giant squid, but if they were plankton-eaters this makes more sense. Indeed, during the Jurassic and Cretaceous the planktonivorous ammonoids continue to increase in abundance, and the heteromorphs are the most common variety by the Late Cretaceous.
Unlike nautilids and coleoids, whose larvae and juveniles are often benthic, ammonoid larvae appear to have been part of the plankton itself. As they mature, males and females look identical in early growth stages, but some (most? all?) genera show marked sexual dimorphism. The (probable) females are larger (macroconchs), while the (probable) males are smaller (microconchs) with a pair of projections (lappets) probably used to position themselves in front of the female and/or guide the specialized inseminating arms to place the spermatophore (sperm packet) into her mantle cavity.
Ammonoids have a very different evolutionary history than trilobites. As we saw above, trilobites peaked in diversity in the Cambrian and Ordovician, and subsequent mass extinction trimmed down the survivors of the previous ones until there were none left. In contrast, ammonoids rebounded after each major mass extinction, with big adaptive radiations. Indeed in the Late Cretaceous ammonoids were at least as diverse as they ever were, and perhaps even at their highest diversity.
Why then did the speciose and common ammonoids die out at the K/Pg while the rarer nautilids survived? It may be that the K/Pg hit the nektonic and planktonic communities more strongly than the benthic ones, unlike the P/Tr and Tr/J. Ammonoids had two strikes against them: their larvae were part of the plankton and the better part of their diversity was planktonivores. Additionally, it might simply have been a matter of random chance. In the previous mass extinctions very few species of ammonoid actually made it through to become the foundations of the next wave of radiations. It may simply have been that at the K/Pg that number wound up as "zero" rather than "a few".

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}