Key Points:
•Looking at their most basal members, Trilobita and Chelicerata show strong similarity, but this is soon overwhelmed by evolutionary novelties on either stem.
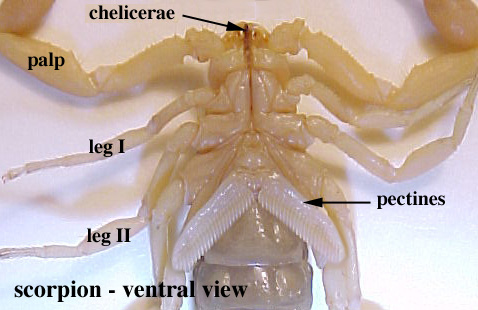
•Chelicerate tagmosis shows specialization of the prosoma (with six pairs of appendages including chelicerae and pedipalps) and opisthosoma(with book gills or their derivative).
•Living pycnogonids are total freaks. Fossil ones, only slightly less so.
•Xiphosura contains plesiomorphic members with distinct chelicerae but unspecialized pedipalps. The group is probably paraphyletic.
•Arachnida, the terrestrial chelicerates, internalize their book gills to form book lungs, and are specialized for external digestion of prey.
•Fossil Stem cheliceramorphs occur in the Burgess Shale.
•Chasmataspidida represents a clade or grade group intermediate between Xiphosura and Eurypterida.
•Eurypterida is a major radiation of large marine chelicerates during the Paleozoic. These were apex predators during the Ordovician and Silurian.
•In the later Paleozoic, euryapsids managed to survive by invading marginal brackish environments and developing some facility at emerging from the water.
•Their geographic distribution suggests that most eurypterids couldn't cross open ocean, so they were geographically restricted.
•The phylogenetic relationships between eurypterid groups and arachnids is unclear.
•Scorpion-like arachnids were among the first metazoans to emerge from the oceans. They occupy a variety of ecomorphs. Aranae (spiders) are especially derived and successful.
•Fossil arachnids include proper scorpions, trigonotarbids, Haptopodids, and Idmonarachne.
•Beyond a spider/whip-scorpion clade - Tetrapulmonata - arachnid phylogeny is unresolved.
"People will be able to survive without honeycreepers and monk seals. But if the wolf spider is in trouble, we are in trouble, too."
(Susanna Moore.)
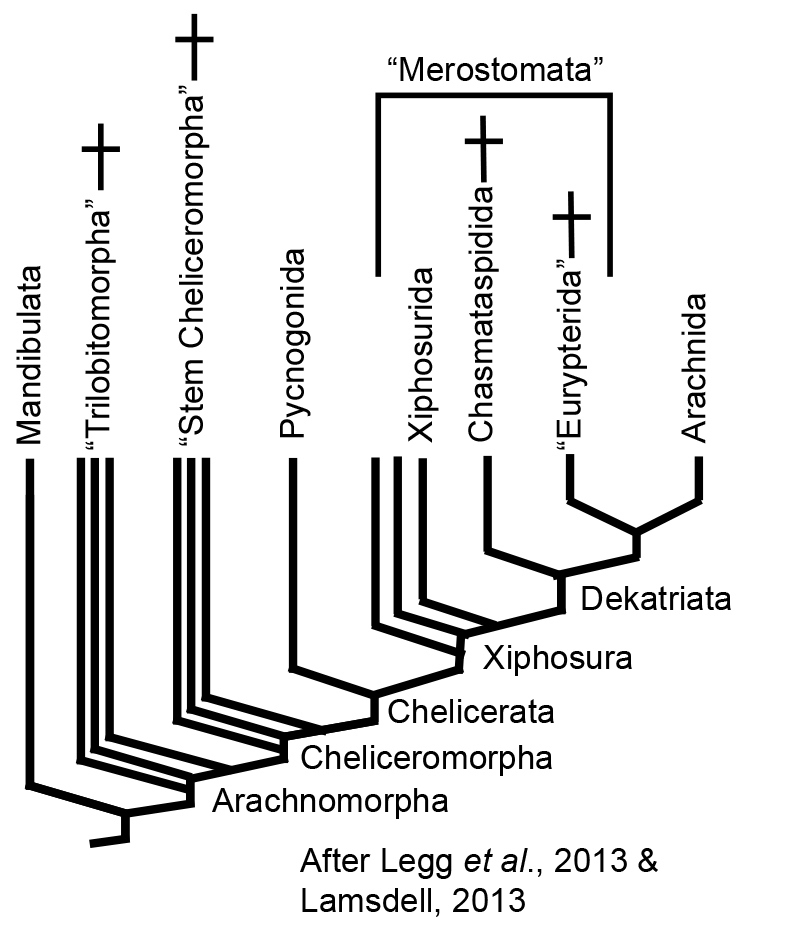
Trilobites and chelicerates are united in the monophyletic group Arachnomorpha: ("Artiopoda" of some researchers.) Synapomorphies are too technical to discuss here, however a comparison of very early trilobites and chelicerates shows their general similarity to their last common ancestor. For a thorough review, see Legg et al. 2013.
Note, for example the post-anal telson (spine), a feature soon lost in trilobites but retained and elaborated in chelicerates. Indeed, one group, Aglaspida, was traditionally regarded as basal xiphosurans, but have recently been identified as basal trilobitomorphs (Ortega-Hernandez et al., 2013)
We have considered Trilobitomorpha previously. Now we turn to Cheliceromorpha - Chelicerata and its closest relatives.
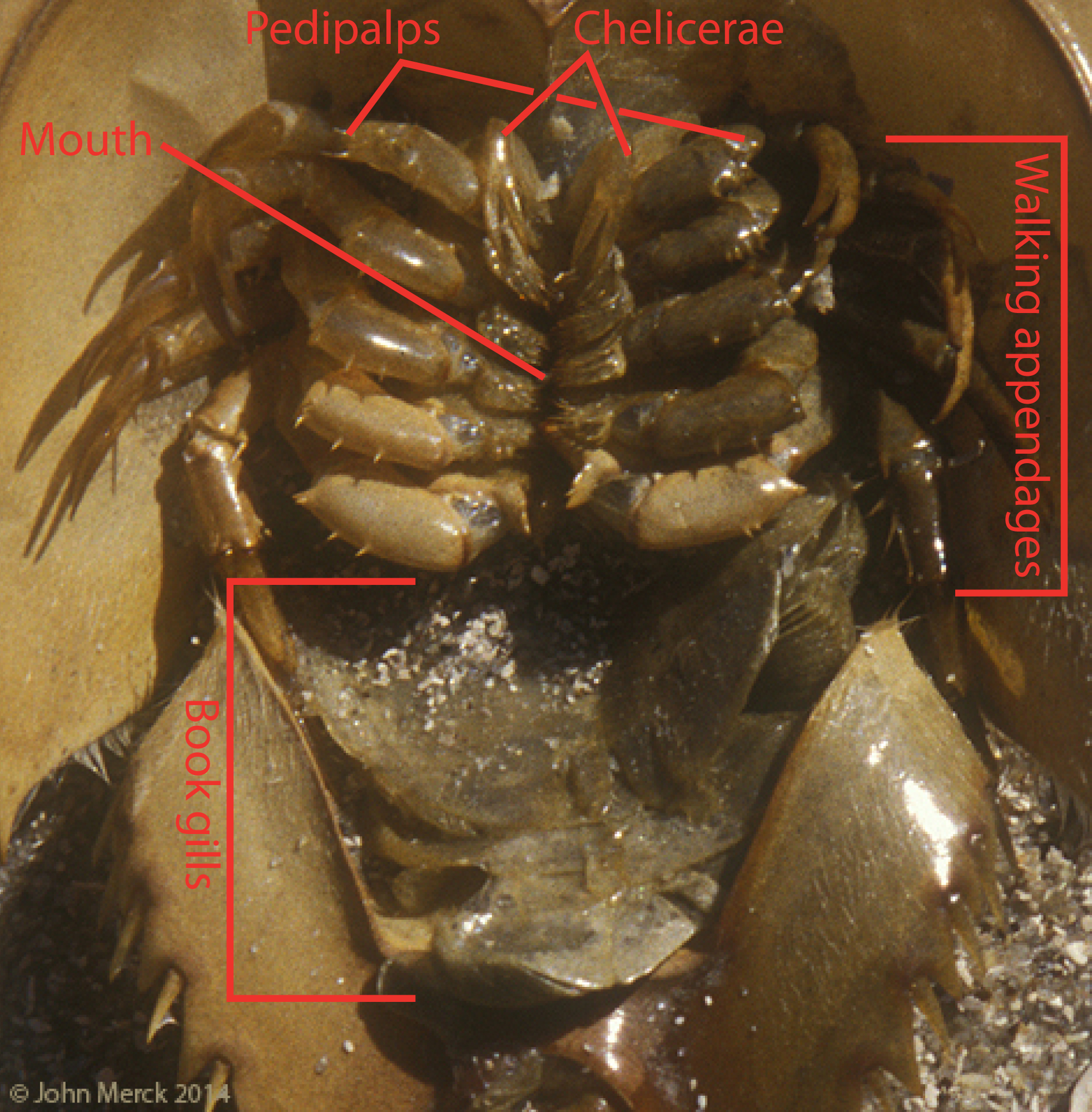
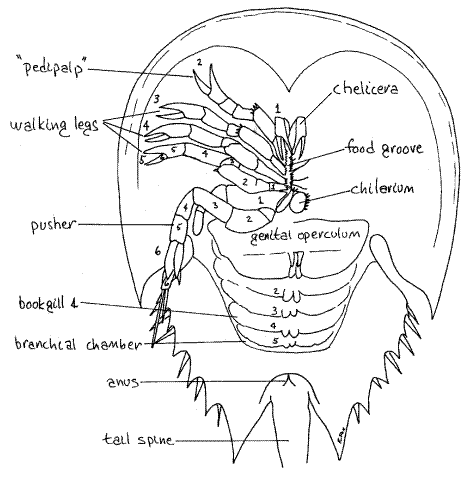
Anterior segments fused to form prosoma (bearing the head and thorax, with six pairs of appendages)
posterior form opisthosoma (primitively segmented); often followed by a
telson, or tail spine
Tagmosis:
NO antennae
Six pairs of uniramous prosomal apendages derived from the endopodite (inner ramus) of ancestral biramous limbs
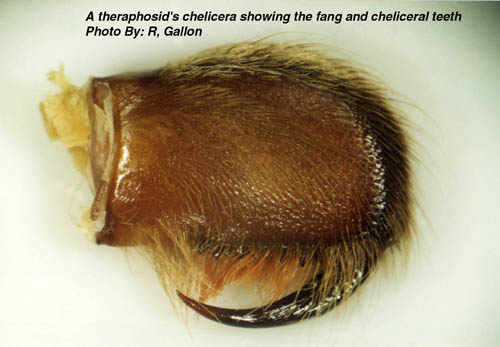
Chelicerae: shred food and transfer to mouth
Pedipalps: Second pair of limbs often specialized for various functions include seizing prey, walking, and copulation
Four pairs of walking legs
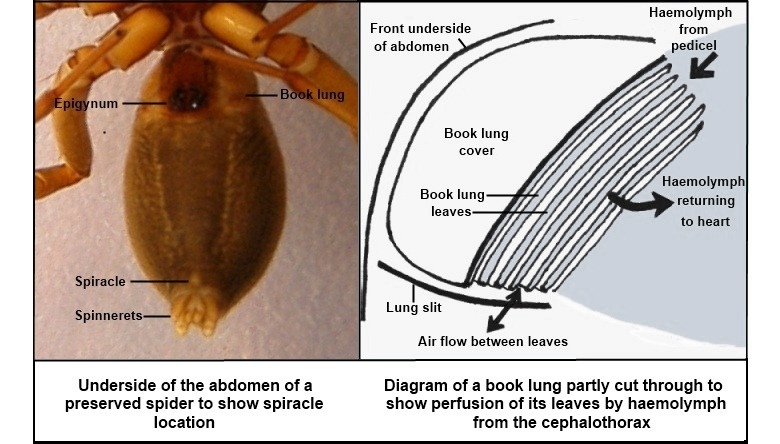
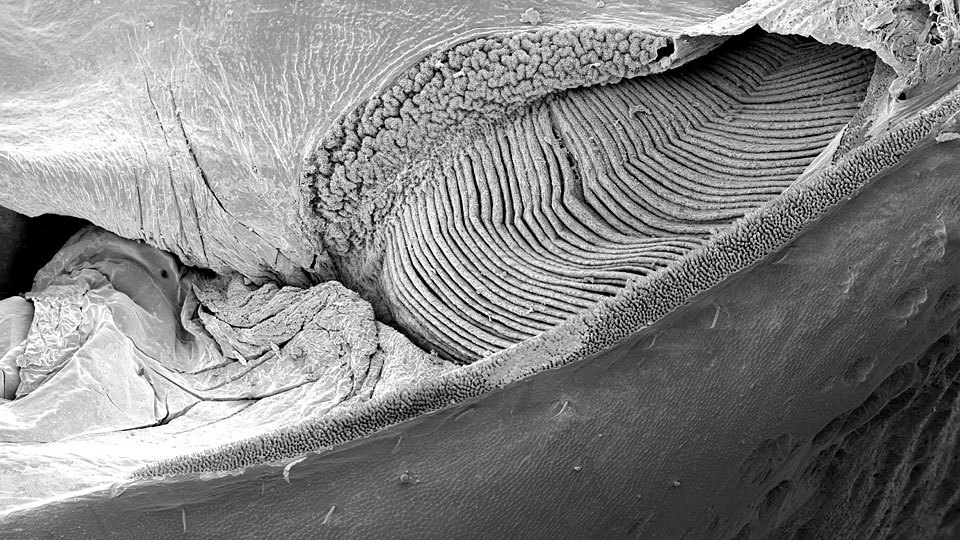
Opisthosomal segments are either without limbs or bear book gills - specialized appendages used for gas exchange derived from ancestral exopodite.
This is the limb pattern in all living chelicerates and most fossils, however the Briggs et al. 2012 description of biramous limbs in a Silurian stem xiphosuran reveals that the earliest chelicerates retained the ancestral arthropod condition.
Trunks are greatly reduced, with gut tube and other organ systems occupying the proximal limb segments.
The opisthosoma is reduced to a small nub.
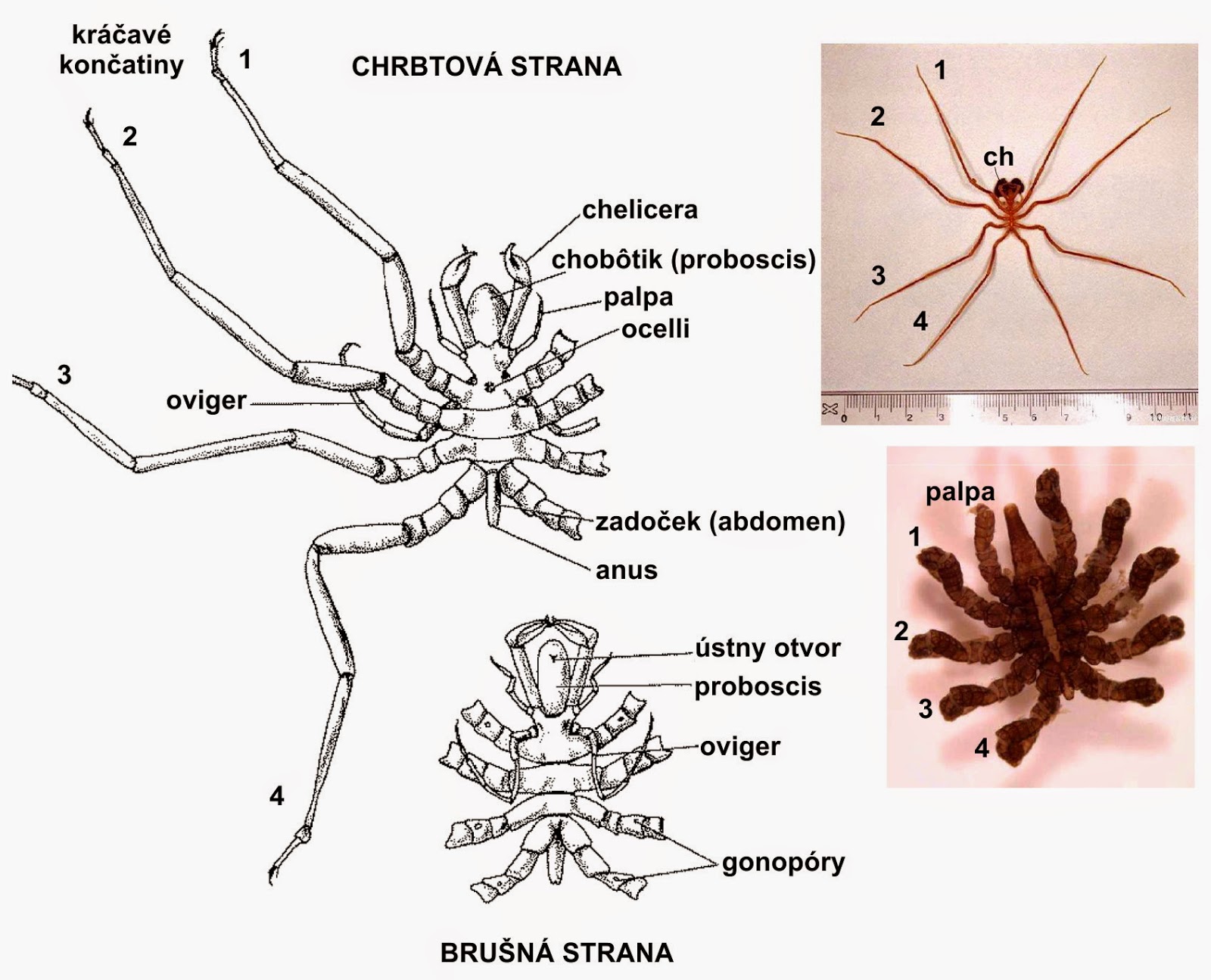
Tagmosis: Almost the entire body is prosomal. Limbs:
Chelifores homologous with chelicerae
"Palps" - pedipalp homologs?
"Ovigers" Males have specialized third limbs for grooming and transporting fertilized egg mass.
Four pairs of walking limbs
Note: Except for the first two pairs, limb homology is problematic here. If we count ovigers then, in comparison with other chelicerates, pycnogonids have an extra pair of limbs. Maybe they result from some kind of duplication, or maybe the last pair of walking limbs is homologous to the first book gills of other chelicerates.
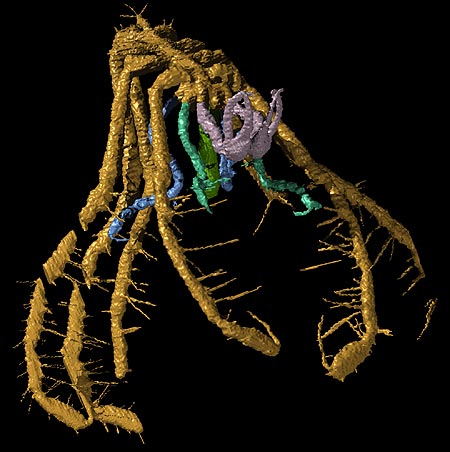
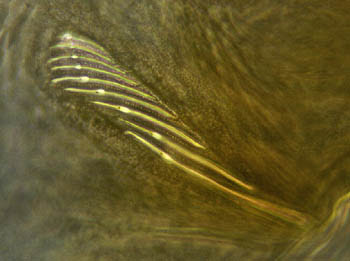
Pycnogonid fossil record: Six fossil pycnogonids are known, one being a larva (Cambrian) and the others adults (Sil. - Dev). Still, this sparse sample demonstrates the usefulness of fossils in phylogenetic analysis:
Living pycnogonids lack many features we associate with arthropods, E.G.:
Compound eyes
Telson
Whether we see this as a reversal or ancestral depends on our picture of the arthropod stem.
If we exclude fossil arthropods from analyses and polarize analyses with outgroups like priapulids, pycnogonids show up as the sister taxon of other arthropods.
The inclusion of fossil arthropods clarifies that these losses in pycnogonids are secondary and allows them to be the sister taxon of other chelicerates.
Fossil pycnogonids, themselves, tell us this much. During the Paleozoic, they had:
Marine, but a few fossil brackish and freshwater forms. Living members can tolerate brackish water and come onto land for short periods of time to spawn.
Opisthosoma is reduced and, in more recent forms fused into an inflexible opisthosomal shield.
Retain compound lateral eyes and simple mid-line ocelli. (Ancestral for Euarthropods.)
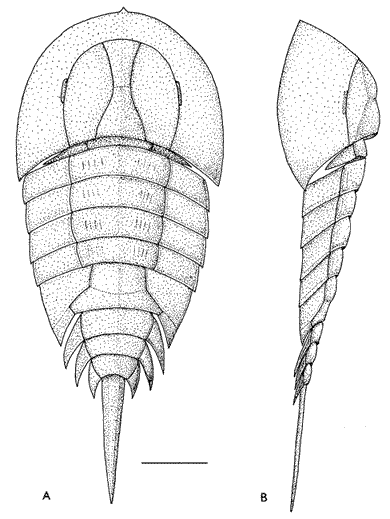
From the Triassic onward, these are very similar to living ones, like Late Jurassic Mesolimulus.
Paleozoic "xiphosurans" (right) show clear segmentation of the opisthosoma.
Arachnida (land chelicerates) (Sil. - Rec.) The land chelicerates. Among the first land animals, the arachnids constitute a diverse radiation into many ecological roles. Their diversity is discussed below. General characteristics:
Although some are adapted to life in or near water, all are air-breathing.
The book gills are internalized into a book lung facilitating air breathing. (Some members also use a tracheole system similar to that of insects.)
Compound eyes are significantly reduced or eliminated
Chelicerae, pedipalps, and walking legs are strongly differentiated
Feeding systems are modified so that digestion is, to some extent, external. (M'mmmm!)
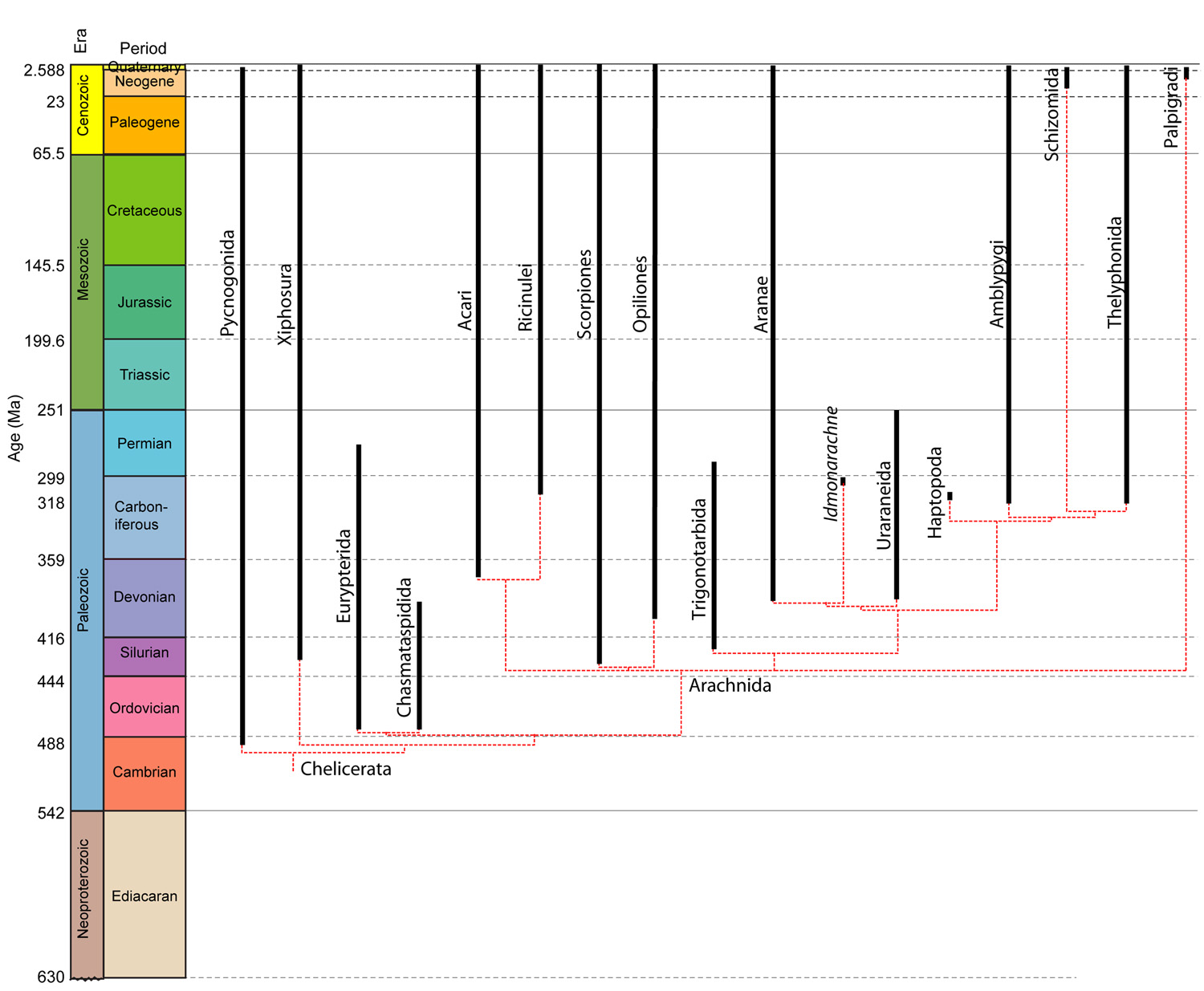
Cheliceromorph phylogeny: Chelicerates have one obvious morphological synapomorphy: chelicerae. Based on their innervation, they appear to be homologous to the antennae of trilobites and most other arthropods (See Legg et al. 2013.) Thus, chelicerates do not so much lack antennae as possess them in a derived form and use them for a different purpose. This seems like a tenuous way to unite this much morphological disparity, however fossil forms fill the gaps and tell their own interesting stories.
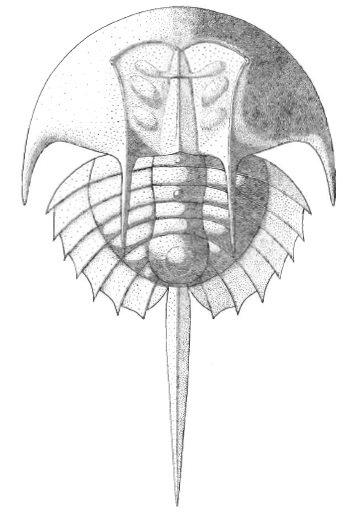
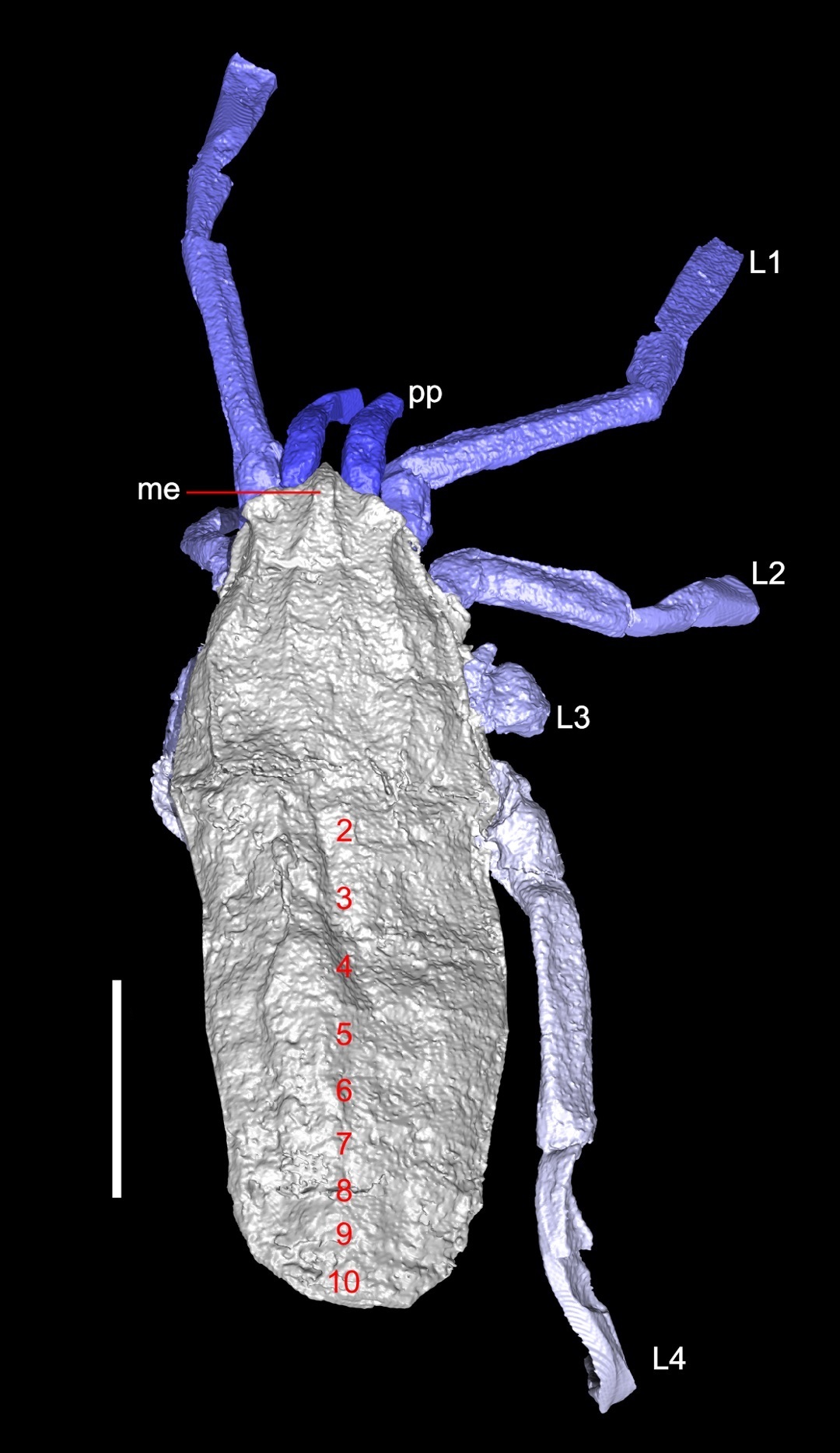
Stem cheliceromorphs (Cambrian) The diversity of animals closer to chelicerates than to trilobites is substantial, and includes some favorites of the Burgess Shale like Sanctacaris (right). Like chelicerates, it has:
A fused prosoma
Six pairs of prosomal appendages, including five robust grasping limbs and one antennate pair. Each grasping limb is biramous, with an antenna-like ramus and a stout one. None have pincers.
Opisthosomal segments each bear a pair of swimming paddles.
the Linnean group "Xiphosura" contained horseshoe crabs and similar fossil creatures.
"Merostomata" contained horseshoe crabs, chasmataspidids, eurypterids, and other marine chelicerates.
"Merostomata" has long been known to be paraphyletic. Lamsdell, 2013 argues that Xiphosura as traditionally defined is, as well and redefines it phylogenetically to be the last common ancestor of living horseshoe crabs and arachnids and all descendants.
The monophyletic Xiphosurida -the clade of horseshoe crabs - is nested within it.
Although one soon tires of "horseshoe crab-grade" fossil chelicerates, things do happen in this part of the tree, where we see trends toward the complete loss of exopodites from the proximal appendages, and the stabilization of the number of opisthosomal segments.
Dekatria: (Ordovician - Quaternary) The last common ancestor of Chasmataspidida, Eurypterida, and Arachnida and all descendants.
Synapomorphy: Thirteen opisthosomal segments.
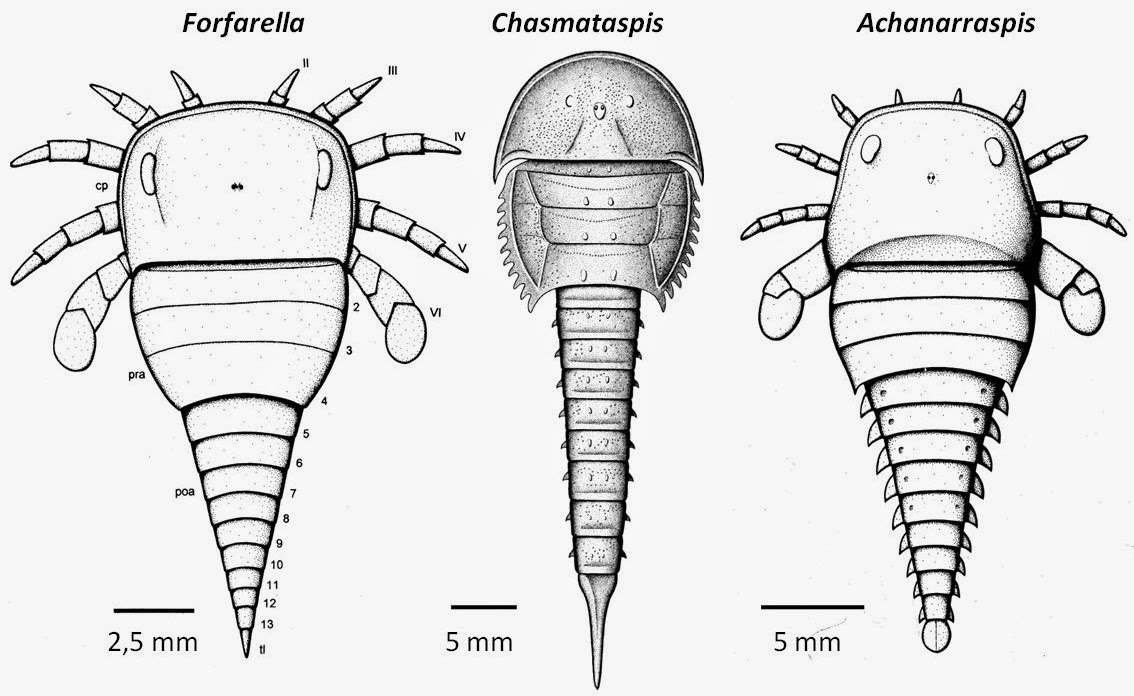
Chasmataspidida (C - D) A minor group whose general morphology spans the gap between Xiphosura and more derived chelicerates. Characteristics:
The final pair of walking limbs modified as paddles.
Opisthosoma divided into anterior preabdomen and posterior postabdomen
Book gills are limited to the preabdomen and concealed in pouches.
These characteristics are shared with eurypterids and (in modified form) arachnids. Chasmataspidids' synapomorphy:
The first three segments of the opisthosoma fused into "buckler."
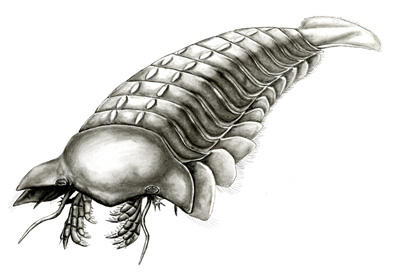
Eurypterida (sea scorpions) (Ord. - Per.): Finally, an ecologically significant, diverse, and widespread chelicerate group. Eurypterids appear in the Ordovician and become top marine predators during the Silurian. Post-Silurian members become increasingly specialized for near-shore and brackish habitats. Characteristics:
Tagmosis resembles that of chasmataspidids, but preabdomen has seven segments and postabdomen only five.
Reached large sizes (more than 2 m long)
Seem to have been more heavily mineralized than xiphosurans or arachnids, possibly because of larger body size.
Feeding ecology: Ranged from xiphosuran-style hunting and scavenging to attacking large prey. In derived forms, either chelicerae or first walking legs could be developed as powerful claws
Sixth appendage pair modified as paddle. Based on joint reconstructions this tended to be used in rowing rather than subaqueous flight. Extimated top speed is about 2.5 X body length per second.
Anteriormost six segments of opisthosoma typically very broad: form preabdomen - Derived book gills enclosed in gill chambers superficially resembling sternites. These chambers have, on their roof, vascularized gill tracts that apparantly functioned to retain water when the creature was on land (a development of the simpler arrangement of chasmataspidids.)
The telson could be broadened into a paddle or remain slender. In some, it is hollow - conceivably used to transmit venom as in scorpions.
Alas, eurypterid systematics are still largely Linnean. Although some phylogenetic analyses have been performed (E.G. Tetlie and Cuggy, 2007 and Lamsdell et al. 2010) but suffer from:
Changing hypotheses of appropriate outgroup choices
Thus, we don't really know if Eurypterida is monophyletic. Significant, because it is often informally suggested that they are paraphyletic with respect to arachnids.
Eurypterid ecological strategies: Eurypterids were top marine predators during the Silurian but had been replaced by vertebrates by the middle Devonian. It's tempting to think that they hung on thanks to their ability to invade marginal habitats, as many held out in brackish environments and seem to have been able to make excursions onto land. Seem to have been euryhaline. Depositional settings of eurypterid fossils indicate that they ranged from fresh to marine water, that individual animals seemed to be tolerant of a broad range of salinity, and that different groups tended to have preferences for a particular environment. Some have limbs that seem robust enough to allow excursions onto land. Evidence includes:
Retention of four pairs of robust walking legs in taxa like Stylonurus (above right).
Internalization of book gills should protect from desiccation.
Thus we have evidence for eurypterids that both ought to be able to come onto land and direct evidence that they did so. Perhaps their ecology was like that of some crabs.
Biogeography: Eurypterids seem to have evolved in the waters around Laurentia, Avalonia, and Baltica (consolidated in Silurian as the paleocontinent "Laurssia.") Their limited distributions suggest that they were restricted to shallow waters. A few managed to disperse to other continents by "rafting" on moving continental crust. Only members of Pterygotidae (right) seem to have been able to disperse across open oceans. (Tetlie, 2007)
Chelicerae, pedipalps, and walking legs are strongly differentiated
External digestion (Yuck!)
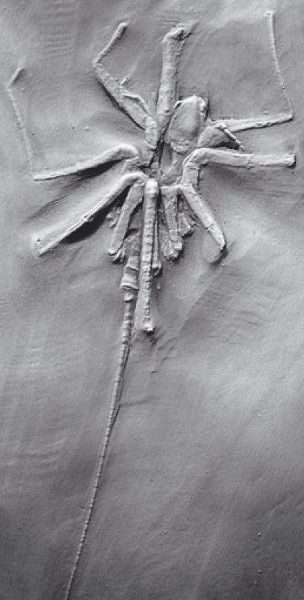
Arachnids and eurypterids are each considered monophyletic, nevertheless it is difficult to look at a scorpion and not wonder whether they, and their derived terrestrial relatives, might not be nested within Eurypterida. By all accounts, scorpions appear early and represent an ancestral form. (Note the number of preabdomen and postabdomen segments.)
Scorpiones (Scorpions Sil - Rec.) Scorpions were among the first animals to invade the land. All scorpions are predatory and employ their telsons to transmit venom.
Acari (Mites and ticks. Dev - Rec.): Very sparse record, but known from the Devonian onward. Their anatomy is dominated by the demands of extreme miniaturization. Contain two major groups:
In some, chelicerae specialized for stabbing and sucking.
harvestman (daddy long-legs)
Opiliones (Harvestman (daddy long-legs). Dev - Rec.): Vanishingly small record. Generalized feeders of forest floors. Their distinctiveness comes from the plesiomorphies they retain, including a broad connection between body segments, a segmented opisthosoma, and the absence of a venomous bites. Some derived features.
Broadly resemble spiders but have segmented opisthosoma in which tergites are divided into three longitudinal sections.
Chelicerae are blade-like but not modified for venom injection. Nevertheless, the presence of filtration hairs around mouth indicates that food was digested externally
Pedipalps unspecialized (but some bear a small claw.)
Here be dragons. Schultz, 2007 underscores the basic problem in a morphological analysis. He (and others) identifies one likely clade - Tetrapulmonata, containing spiders and the various whip scorpion groups. however he can't resolve other relationships near the base of Arachnida.
Sharma et al.'s 2014 phylogenomic analysis suggests that very rapid evolution of portions of the arachnid genome swamps phylogenetic signal. Correcting for this by using only slowly evolving portions of the genome, they reach the surprising conclusions that:
Acari is polyphyletic, with ticks being closer to Tetrapulmonata and mites closer to pseudoscorpions.
Arachnida, itself, is paraphyletic (!!!) with Xiphosurida emerging as the sister taxon of Ricinulei.
Is our assumption that chelicerates became terrestrial only once valid at all? Stay tuned!
James C. Lamsdell, Simon J. Braddy, and O. Erik Tetlie. 2010. The systematics and phylogeny of the Stylonurina(Arthropoda: Chelicerata: Eurypterida). Journal of Systematic Palaeontology 8(1): 49-61.
David A. Legg, Mark D. Sutton & Gregory D. Edgecombe. 2013. Arthropod fossil data increase congruence of morphological and molecular phylogenies. Nature Communications 4(2485).
J. Ortega-Hernandez, D. A. Legg, S. J. Braddy. 2013. The phylogeny of aglaspidid arthropods and the internal relationships within Artiopoda. Cladistics. 29: 15Đ45.
Jerome C. Regier, Jeffrey W. Shultz, Andreas Zwick, April Hussey, Bernard Ball, Regina Wetzer, Joel W. Martin, and Clifford W. Cunningham. 2010. Arthropod relationships revealed by phylogenomic analysis of nuclear protein-coding sequences. Nature 463, 1079-1083.
Prashant P. Sharma, Stefan T. Kaluziak, Alicia R. Perez-Porro, Vanessa L. Gonzalez, Gustavo Hormiga, Ward C. Wheeler, Gonzalo Giribet. 2014. Phylogenomic interrogation of Arachnida reveals systemic conflicts in phylogenetic signal. Molecular Biology and Evolution August 2014.
O. Erik Tetlie and Michael B. Cuggy. 2007. Phylogeny of the basal swimming eurypterids (Chelicerata; Eurypterida; Eurypterina). Journal of Systematic Palaeontology 5(3).









.jpg)








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}