Fall Semester 2022 Deuterostomia II: Crown group echinoderms
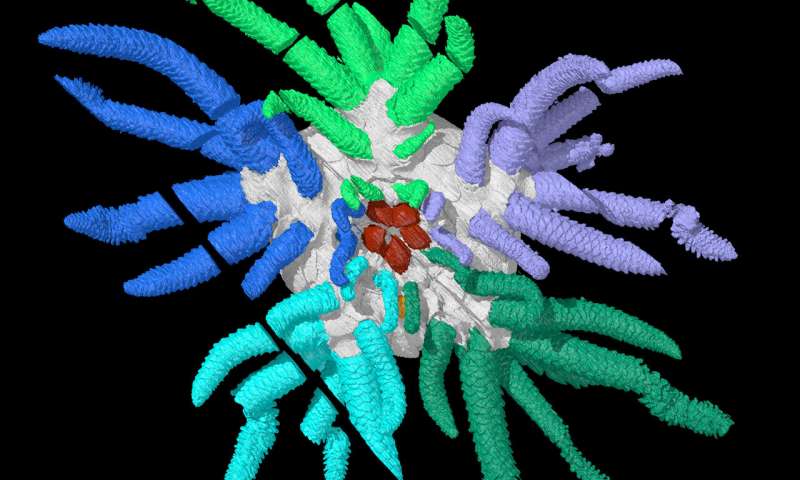
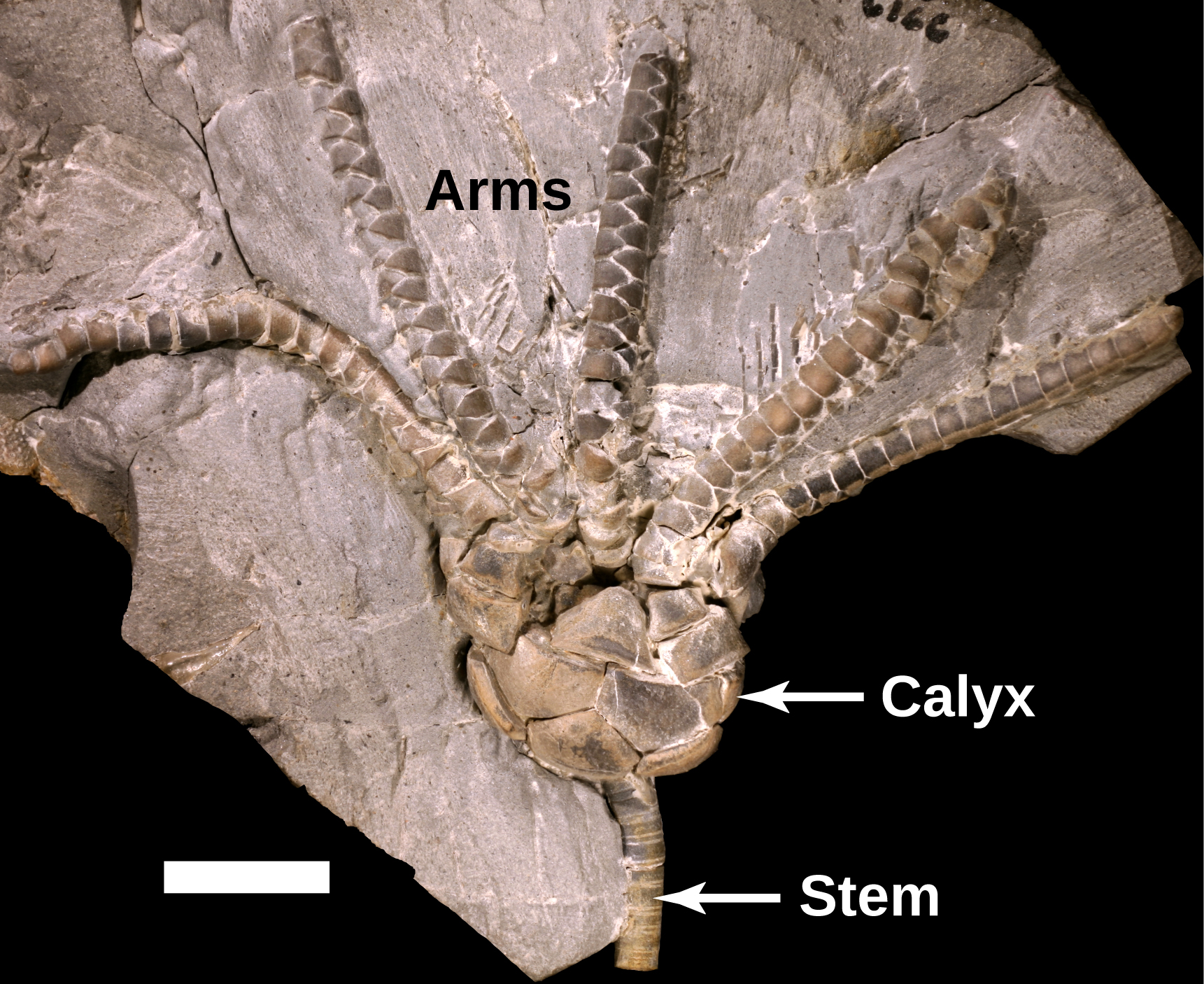
Uintacrinus
Key Points:
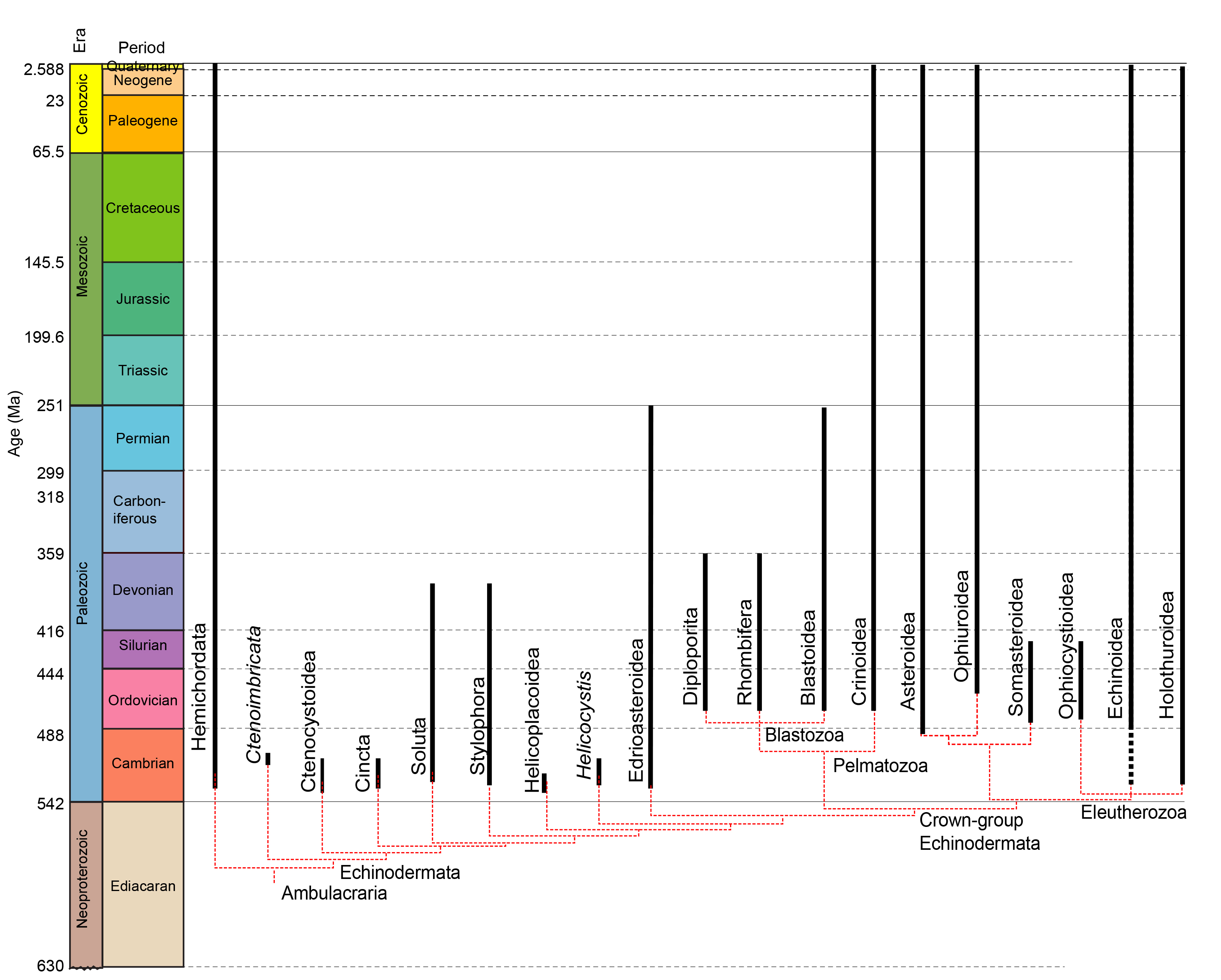
•Hypotheses of the phylogeny of living echinoderms are currently in a state of upheaval. This review assumes that both Pelmatozoa and Eleutherozoa are monophyletic, following the emerging majority view of the last decade.
•Crinoid morphology is distinct from that of other stalked echinoderms.
•Of the major crinoid groups, only Articulata survived into the Mesozoic.
•The Post-Paleozoic world has been hard on crinoids, who have either retreated to marginal habitats or significantly altered their morphotype.
•Although we think of Eleutherozoa as the "modern" echinoderms because of their success in the Post-Paleozoic oceans, they are roughly the same age as Pelmatozoa.
•Eleutherozoa contains Asterozoa (starfish and brittle stars) and Echinozoa (echinoids and holothurians) and their fossil relatives.
•Ophiuroidea (brittle-stars) are scavengers on particulate and larger food fragments, very common in the deep oceans.
•Asteroidea include predatory forms.
•Somasteroids - stem asterozoans - are flattened, five-armed deposit/suspension feeders.
•In contrast to other Eleutherozoans, echinoids have a robust fossil record.
•Echinoid evolution presents a series of interested paraphyletic grade groups leading up to Clypeasteroida - sand dollars.
•Holothurians have an especially poor record, as their skeletons are not integrated. Encompass a range of shallow and deep marine deposit feeders.
•Ophiocystioideans represent stem echinozoans with features suggestive of echinoids and holothurians.
"Never ask a starfish for directions."
(Anonymous)
We now explore the crown group of Echinodermata, containing:
Pelmatozoa with crinoids as living representatives.
Ambulacra are lined with brachioles - elongate feeding structures supported by skeletal elements and supporting tube feet. (Link to Gogia with conspicuous brachioles.)
Stalks present for elevating main body off of the substrate. (Only some are developed for attachment to substrate.)
Tend to have relatively complex internal breathing structures developed from the WVS.
Main bodies called thecae may be large but typically compact - spherical or ellispoidal without "arms' containing extensions of normal coelom.
Strong pentameral symmetry and beautiful geometric regularity.
Ambulacra:
Lined by brachioles
Ambulacral grooves covered by lancet plates.
Ambulacra terminate at the apex of the theca in the center of which is the mouth.
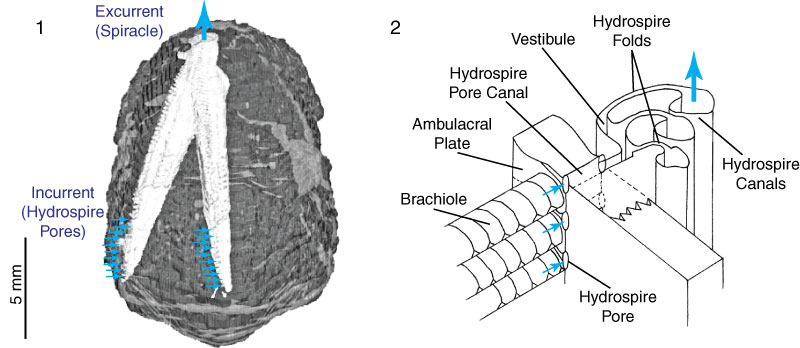
Ambulacra lined by inlet pores that led to complex pleated WVS chambers - hydrospires - that functioned in gas exchange.
The mouth is surrounded by five spiracles that act as outlets to the hydrospires and exits for gametes from gonads.
One spiracle is larger than the others. It is a joint spiracle - anus, the anispiracle.
Blastoid stalks allowed them to attach to the substrate.
Evolutionary trends: Blastoids became common in the Early Carboniferous (a great time for stalked echinoderms generally) declined, then resurged in the Permian before being stomped by the Permian extinction.
The remaining groups are traditionally termed "cystoids."
Paraphyletic, but similar in having a stem that didn't terminate in a holdfast. Apparently they crawled around then stuck their thecae up when they found a good place to suspension-feed. Generally, their thecae were somewhat irregular and slightly flattened (i.e with a top and bottom.) We see two clades major (among many others).
Diploporita: (Early Ordovician to Late Devonian), respired through diplopores (paired holes giving access to WVS.)
Evolutionary trends: Rhombiferans and dipoporites were never common, but achieved their peak early - in the Ordovician, then declined slowly to be extinguished in the Late Devonian.
(Sea lilies and feather stars) (Ord. - Rec.) Finally, a group with living representatives.
The crinoid body plan:
A stem consisting of poker-chip shaped columnals from which small tentacles - cirri - branch. The stem terminates at a holdfast. Despite this, the crinoid is capable of moving around when it wants.
At its "lip," the cup branches into five arms, which often subdivide. Each arm bears an ambulacrum on its upper side and an outpouching of the proper coelom internally.
These are sometimes lined with calcareous pinnules that assist in filtration and tube-feet that snag food particles and convey them to the ambulacral groove.
Because the tube feet are not used for locomotion, they lack ampullae.
The top is the oral surface or tegmen. The mouth is in the center with the anus to one side, as in edrioasteroids and blastoids.
By convention, the aboral surface is toward the stem.
Crinoids don't move much if they have found a good place to feed, but may crawl or swim to good feeding cites on a daily basis.
Suspension feeders: The crinoid points its oral surface downstream then arches its arms into the current. Crinoids can quickly orient themselves on their stems using a bizarre trick: The connective tissue running down their stems can be partially emulsified by nervous impulses, thus rapidly altering stiffness. Wow.
There may be plates on the tegmen covering the mouth and anus. In some cases, the anus is armored with an anal pyramid to guard against parasitism by gastropods who seemed to have viewed crinoid butts as sort of a meal ticket.
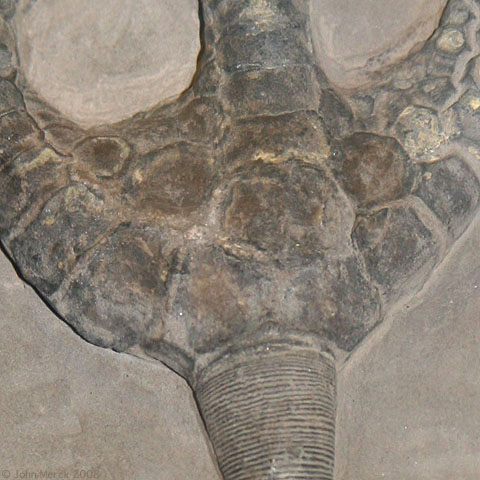
Actinocrinites gibsoni
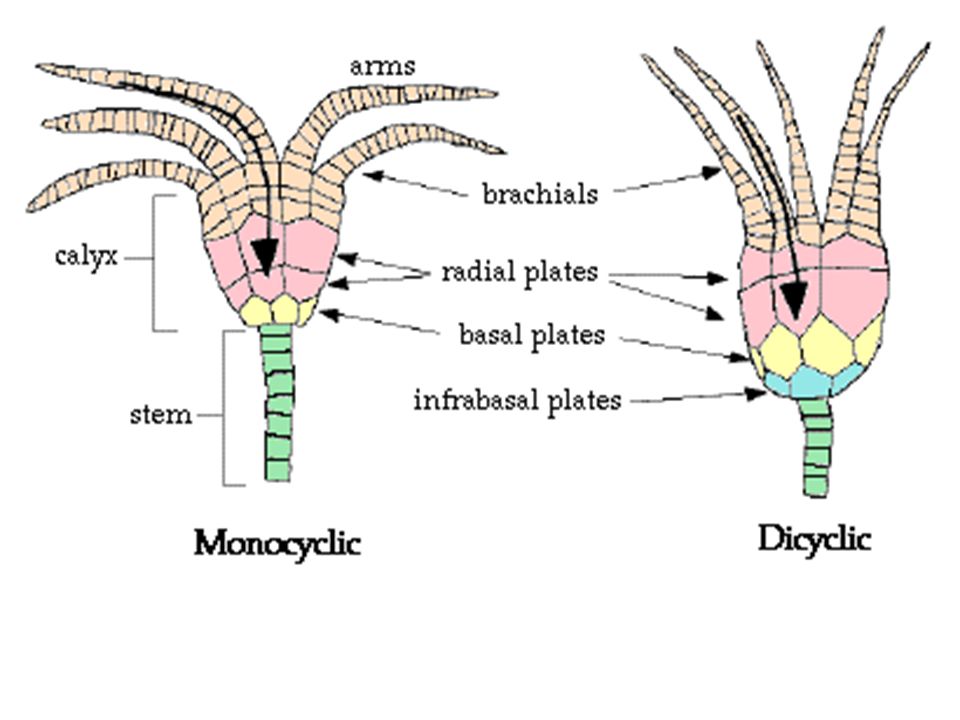
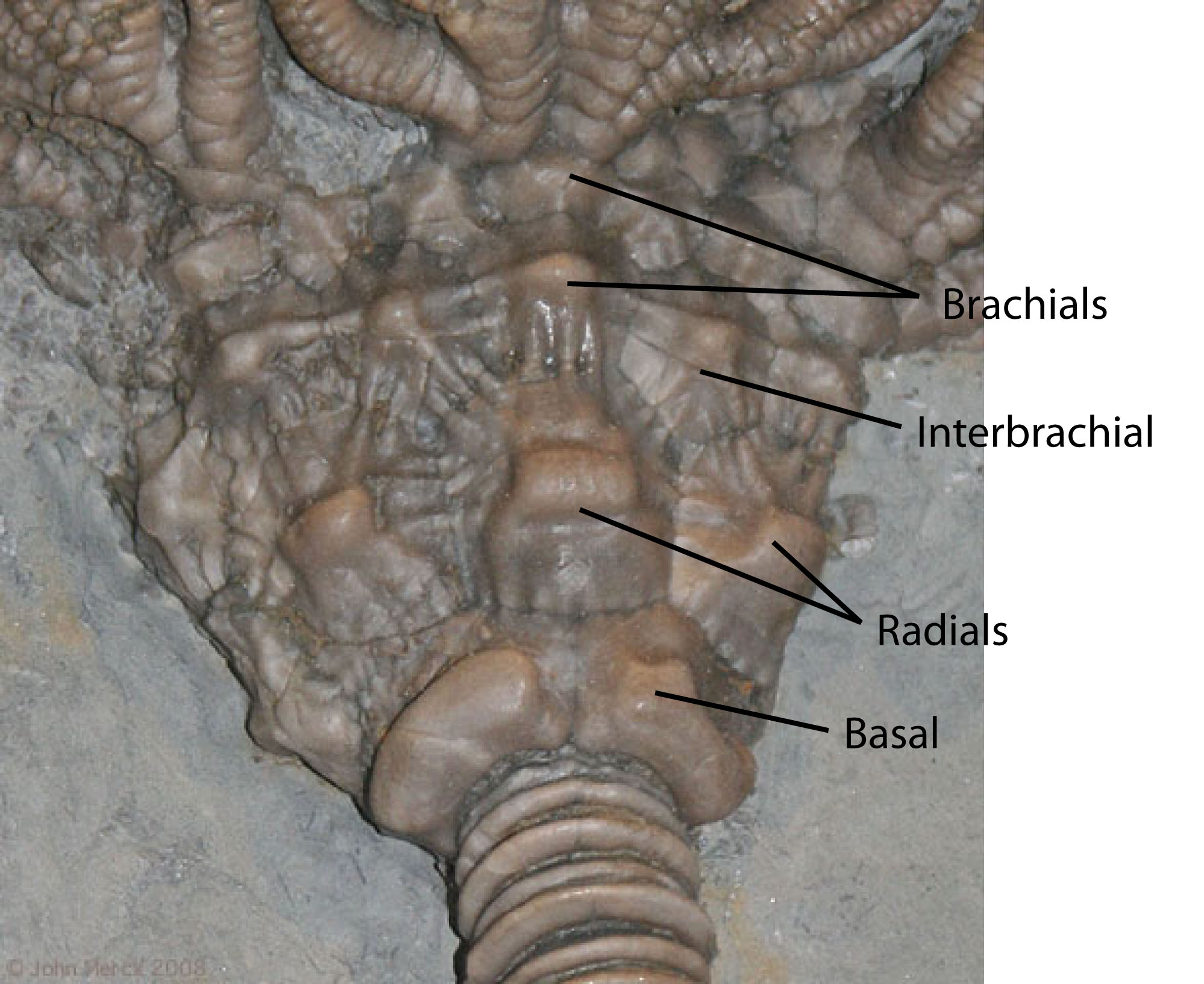
What is your assessment of the morphology of the crinoid at right:
Monocyclic or dicyclic?
Are branchial or interbranchial plates present?
Are arms uniserial or biserial?
Crinoid systematics:
Phylogenetic systematists have only just begun their exploration of crinoids, so we offer no synopsis of phylogeny here. Interested students can refer to Ausich et al. 2015. Traditionally, crinoids break down into three major groups that we present for descriptive purposes:
Simple type of structure, consisting of radials, basals, and anals, with some forms having brachial plates.
The arms lack pinnules.
Disparida contains tiny forms, with calyx heights of 1mm. Others like the "pea crinoid" Pisocrinus are remarkably abundant in the Silurian., and these, along with similar Devonian forms, had calyces of 2 to 4 mm.
Actinocrinites sp.
Camerata: (Ord. - P)
Rigid calycal plates.
Thick plates of the tegmen
Have fixed brachial and interbrachial plates forming part of the calyx.
Paleozoic (starting in Ordovician, but especially Early Carboniferous - right) crinoids were common in calm shallow water, favoring back-reef facies and the landward sides of barrier islands - environments with well circulated water but not too much energy.
Permian extinction hurt. In post-Paleozoic world, surviving crinoids are rare. Modern crinoids stalked are restricted to deep oceans - depths >100 m.
Stalkless Comatulids are the most diverse living group, and are common in tropical reefs. They walk around on their cirri or swim slowly to find a good spot for suspension feeding.
The general impression: The post-Paleozoic has been a hostile time. Crinoids have either retreated to realms where predators are rarer (deep oceans) or evolved greater mobility to avoid them.
Note, however, Mesozoic Pentacrinitidae. Large crinoids with long stems attached to flotsam. Thus invading pelagic planktonic realm.
There are more radical suggestions of an unexpected crinoid radiation (amazed scientists exclaim!!!)
Eleutherozoa:
(Ord. - Rec.) Eleutherozoa contains the living groups of non-stalked echinoderms. These became the dominant echinoderms of the post-Paleozoic world, and continue to diversify. Do not think, however, that they are of recent origin. The four major Eleutherozoan groups are roughly as old as blastozoans and crinoids. Thus, although relative abundances and diversities have changed, echinoderms diversified quickly into their modern groups during the Ordovician. The illusion of Eleutherozoans being "new" is heightened by the poor preservation potential of all groups except for echinoids.
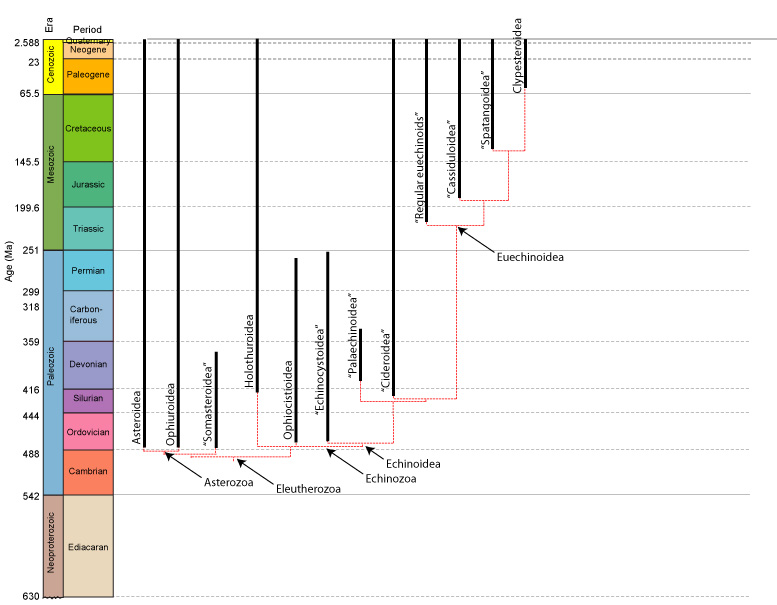
Eleutherozoan phylogeny:
More recent cladistic analyses support the following tree, however in the last twenty years, enough been suggested that a strict consensus would be a whisk-broom polytomy:
Eleutherozoa characteristics:
Not necessarily synapomorphies:
In most cases, tube-feet are adapted for locomotion, as opposed to suspension-feeding.
Ancestrally, the mouth is ventral and the anus dorsal, completely erasing any vestage of original bilateral symmetry. We speak only of oral and aboral surfaces. (But many derived forms modify this.)
No stalks. Is the loss of stalks a synapomorphy or were Eleutherozoans simply derived from non-stalked edrioasteroid-like forebears? Good question.
Eleutherozoan systematics: The cladogram at right represents a decent consensus of current analyses of Eleutherozoan phylogenies with one huge caveat: Two reasonable hypotheses exist for the placement of Ophiuroidea:
The Asterozoa hypothesis (Telford et al., 2014) holds that ophiuroids and asteroids are sister taxa
The Cryptosyringid hypothesis (Reich et al., 2015) maintains that ophiuroids are sister taxon to Echinozoa.
Here we default to the Asterozoa hypothesis, but the issue is unsettled.
Somasteroid in oral view
Somasteroidea - (Ord. Dev.) These appeared slightly prior to the appearance of ophiuroids and asteroids, and were extinguished by the Devonian extinctions.
Characteristics:
Resemble asteroids in having bodies with indistinct central disks and five arms, however they are significantly flattened.
The skeleton of the aboral surface is simple and not rigidly integrated, consisting of simple papillae.
The skeleton of the oral surface consists of ambulacral ossicles and radiating branches of virgals giving the skeleton a feather-like profile.
Speculated to have been benthic suspension or detritus feeders.
Somasteroids are argued by some to be the plesiomorphic morphotype for modern asterozoans.
Made up of a single row of large calcite plates termed vertebrae or vertebral plates. (Confusing??!!)
Although tube feet are present, they lack ampullae (in this respect, similar to crinoids). They are used strictly for feeding. Unlike in other echinoderms, movement of the arms is effected directly by muscles. Thus, ophiuroids walk/climb around on their five arms, making them the fastest moving echinoderms (woo hoo!).
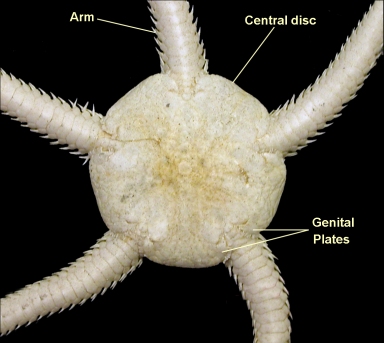
Commonly called "brittle-stars" because of their tendency to shed arms when molested. these are regenerated in a matter of months.
Central disks:
Mouth is ventral in the center of the oral surface.
Anus is absent. Waste is expelled through mouth.
There may be several madreporites that open on the oral surface, near the mouth.
Oral surface covered with small calcite scales, but aboral surface bears five pairs of large shields.
Poor fossil record as they tend to break apart after death. Nevertheless, some trends are visible:
Whereas modern ophiuroids have only one row of "vertebrae" per arm. Early members (from Early Ordovician), termed "oegophiuroids" (paraphyletic?) had double rows of alternating vertebrae and open ambulacral grooves, as in the earliest fossil ophiuroid, Pradesura (right).
Ophiuroid ecology:
Capable of capturing small food particles by crinoid-style suspension feeding using tube feet.
Can use entire arms to capture larger particles, including small prey items.
Present in shallow marine environments, especially reefs, where they hide in crevices or free-load off of sponges.
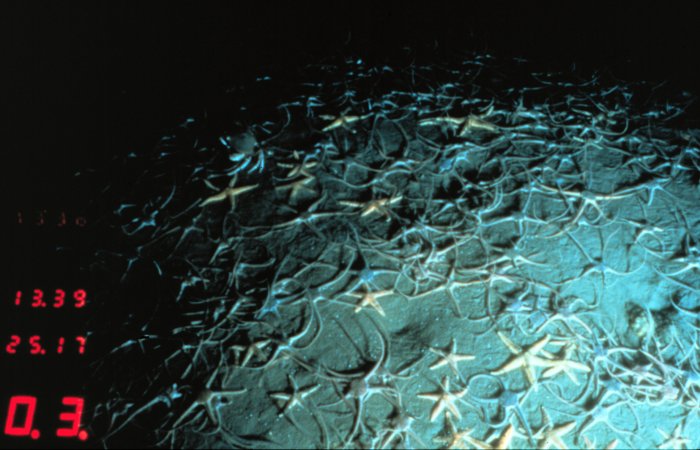
Become very common in deep oceans, where they, along with holothurians, constitute the bulk of the biomass.
One odd group, the basket stars have long subdivided arms and are more fully committed to suspension feeding.
Asteroidea ("starfish" or "sea stars") - (Ord. Rec.)
Taken as "standard" echinoderms for illustrations of tube-feet, the WVS, etc. only because they are familiar and accessible. Actually quite idiosyncratic.
Characteristics:
As in ophiuroids, a central disk and arms are present, but in this case poorly demarcated.
Made up of many small plates that tend to fall apart upon death.
Tube feet sport ampullae and are used strictly for locomotion/grasping. Unlike ophiuroids, asteroids don't move their arms directly by muscular action.
Capable of autotomizing (voluntarily severing) arms to sidetract predators and have prodigious powers of regeneration.
Central disks:
Mouth is ventral in the center of the oral surface.
Anus is very small, owing to characteristic feeding mode (see bleow)
Have two distinct stomachs, a cardiac stomach above the mouth, and a pyloric stomach above the cardiac stomach. Branches of the pyloric stomach, the pyloric caeca (sing. caecum) radiate into each arm.
Predators, preying on largish prey. How this is done depends on the size of the prey item:
Smallest items ingested directly at the mouth.
For larger items, the cardiac stomach can be everted to engulf the prey then pulled back inside.
For large bivalves, the valves are pulled apart slightly, the cardiac stomach everted into the shell, and the bivalve digested by the asteroid and its nutrients absorbed inside its own shell. When the meal is over, the asteroid zips up its cardiac stomach and is good to go. In this case, waste elimination isn't an issue, so the asteroid anus can be small because it doesn't get much of a workout.
Present in a wide range of marine environments.
Hudsonaster matutinus (Ord.)
Fossil Record: Although common today, aspects of starfish biology limit their preservation potential:
Their skeletons disaggregate upon death
The decay of internal soft tissues causes them to collapse and become misshapen, even when they don't fall apart.
They tend to favor hard substrates with low preservation potential.
And yet we have fossils of Ordovician starfish that are similar to living ones in anatomy. (E.G. Hudsonaster right). The Devonian, Permian, and Triassic extinction events each trimmed their diversity, but after each, they rebounded with a proliferation of new groups.
Galápagos green sea urchin Lytechinus semituberculatus
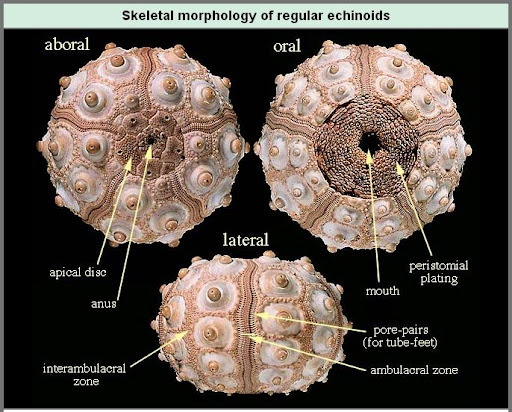
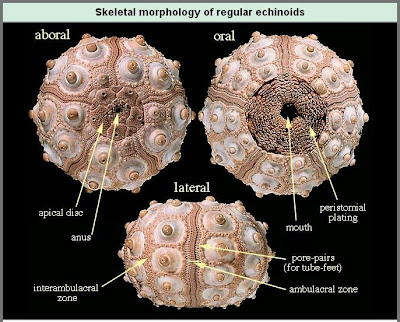
Palaechinoidea: (Sil. - P) Interambulacra contain regular, non-overlapping plates. No sign of spines, however the test is rigid. Roughly grapefruit-sized. (E.G.: Carboniferous Melonechinus indianensis - right)
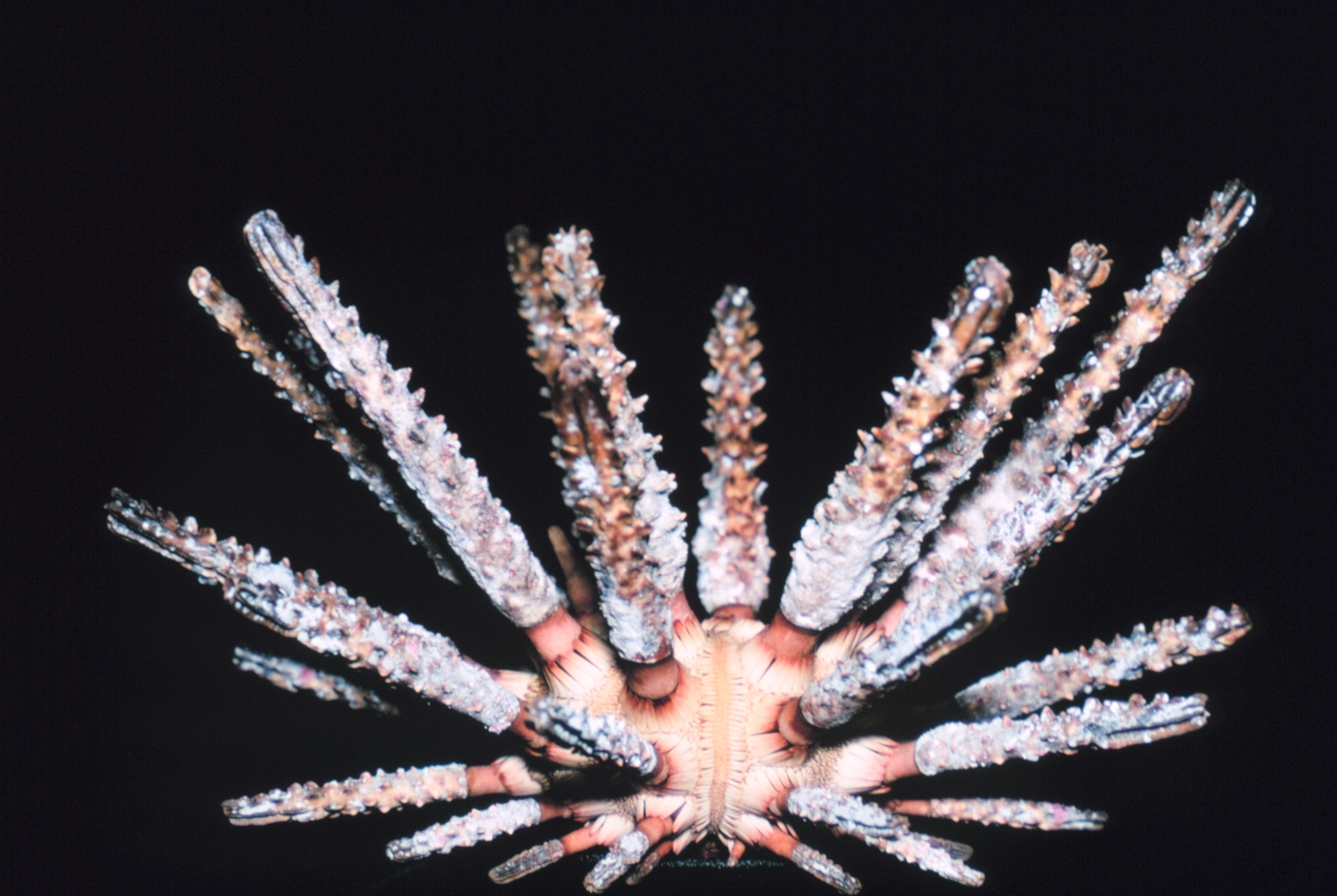
Stereocidaris hemigranosus (K)
"Cidaroidea": Slate-pencil urchins - (Dev. - Rec.) Paraphyletic.
Interambulacra contain one to two rows of plates, each with one support for a very large thick spine. Among all echinoids, only a handful of cidaroid lineages survived the Permian extinction to give rise to the full range of living echinoids. Palaechinoids and echinocystitoids were extinguished.
Tests of cidaroid Eucidaris thouarsii (left) and euechinoid Lytechinus semituberculatus (right)
Euechinoidea: Sea urchins - (Tri. - Rec.)
Monophyletic! Interambulacra contain two rows of plates, each with multiple supports for many slender spines (among many other technical synapomorphies).
Although many euechinoids retain this basic morphology, the group also contains members that have, progressively, re-evolved bilateral symmetry. As a group, these irregular echinoids are monophyletic, but traditionally they have been broken into paraphyletic grade groups.
"Spatangoida": (Heart urchins) (K. - Rec.) Paraphyletic.
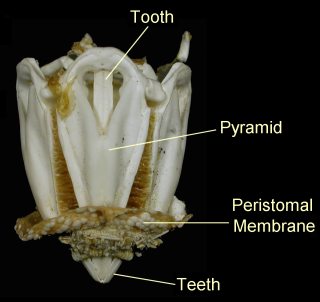
Aristotle's lantern completely absent.
Mouth migrates to anterior position.
Parascutella sp.
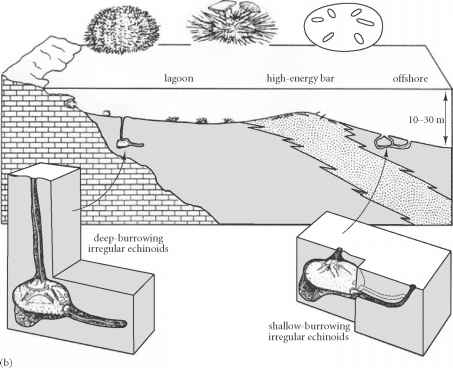
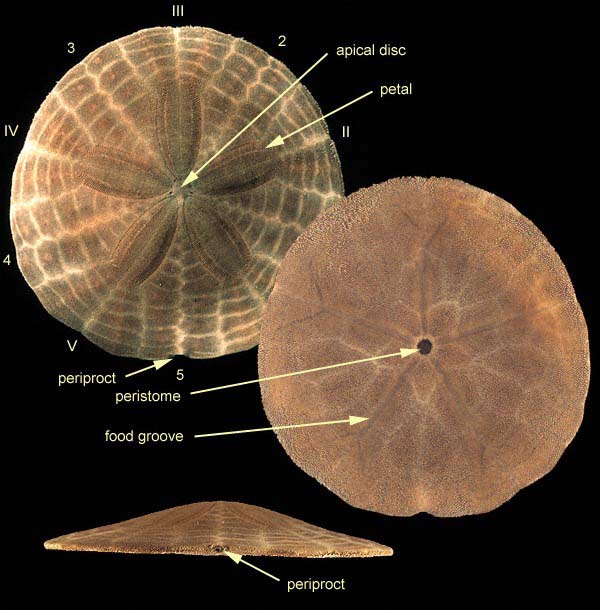
Clypeasteroida: (Sand dollars) (Paleocene. - Rec.) Monophyletic Highly specialized for life at sediment/water interface above wave base:
Test sharply flattened.
lunules - anomalous passages between oral and aboral surfaces reduce tendency to be flipped over by currents
Mouth is in ancestral position on oral surface.
The role of ambulacra of other echinoderms has been assumed by actinal furrows on the oral surface.
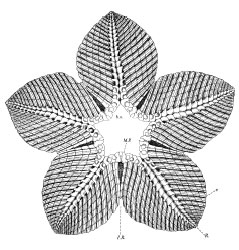
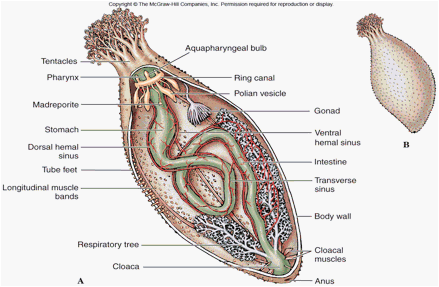
Holothurian shows four of five ambulacra.
Holothuroidea: (sea cucumbers) (Sil. (maybe Ord.?) - Rec.) Suspension and deposit feeders whose body plan resembles that of a regular echinoid that has been:
"Softened up" by the reduction of its hard skeleton
Stretched between its oral and aboral apices until it looks like a sausage
Laid on its side
The result is an animal with a mouth in front, an anus in back, two ambulacra on top and three underneath (on which it walks).
With the exception of the periesophageal ring elements encircling the mouth, there is no rigid test, only calcareous sclerites suspended in fleshy tissue. Thus fossil record is poor. Probable sclerites occur in the Ordovician and are certainly there by the Silurian. Good body fossils are very rare,
Burrowing in sediment is achieved by contraction of body wall muscles against hydrostatic skeleton.
Around the mouth (which is anterior) specialized tube feet are developed into suspension feeding tentacles in some. Others are deposit feeders.
The madreporite is internalized, thus the WVS loses its connection to the outside.
Internally, holothurians have significant intestines, and complex respiratory trees - breathing structures that form as outpouchings of the cloaca.
In contrast, the gonads discharge gametes near the mouth.
Coolest of all, holothurians respond to threats by partial or total evisceration of the intestines and respiratory tree through the anus. In some types, this is limited to the expulsion of special thread-like sticky/toxic cuvierian tubules, that seem to serve no other purpose. Eviscerated organs are eventually regenerated.
Thelonota anamas
Ecology:
Motile, mostly epifaunal deposit feeders that ingest sediment and digest the good parts, leaving castings of sediment.
Range roughly parallels that of ophiuroids - present, even common in reefs and shallow water (see image rt.) but fabulously abundant on the deep ocean floor.
What would an animal close to the close to the ancestry of Echinozoa look like? Arguably like an ophiocystioid. Considered by Rahman et al., 2019 to be a paraphyletic grade of stem holothuroideans.
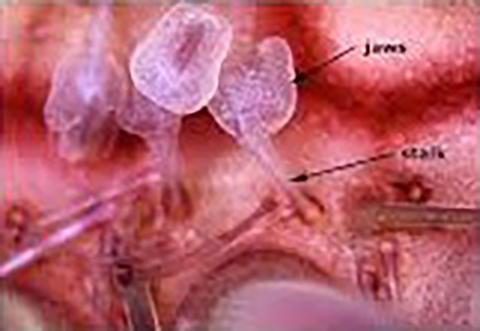
Resemble echinoids in having an oral surface that faces down, with a five part jaw-like apparatus, consisting of goniodonts. This apparatus is simpler than that of echinoids and its elements are oriented horizontally.
Depending on the taxon, the skeleton may consist of firmly sutured plates (like echinoids), or of elements free-floating in the skin (like holothurians).
Tube feet are few in number, large, armored with skeletal elements, and organized around reduced ambulacra. (Reminiscent of the robust tube feet of some holothurians, including the "sea pig," Scotoplanes.
The recent addition, Sollasina cthulhu (Rahman et al., 2019) reveals traces of a ring canal, and other soft tissue features.
Additional reading:
W. I. Ausich, T. W. Kammer, E. C. Rhenberg, D, F. Wright. 2015. Early phylogeny of crinoids within the pelmatozoan clade. Palaeontology, 58(6 937-952)
Imran A. Rahman , Jeffrey R. Thompson , Derek E. G. Briggs , David J. Siveter , Derek J. Siveter and Mark D. Sutton. 2019. A new ophiocistioid with soft-tissue preservation from the Silurian Herefordshire Lagerstätte, and the evolution of the holothurian body plan. Proceedings of the Royal Society B, 286(1900).
Adrian Reich, Casey Dunn, Koji Akasaka, and Gary Wessel. 2015. Phylogenomic Analyses of Echinodermata Support the Sister Groups of Asterozoa and Echinozoa. PLoS One. 2015; 10(3): e0119627.
Sumrall, C. 2015. Understanding the oral area of derived stemmed echinoderms. 169–173. In Zamora, S. and Rábino, I. (eds). Progress in echinoderm palaeobiology. Cuadernos del Museo Geominero, 14, 291 pp.
M. J. Telford, C. J. Lowe, C. B. Cameron, O. Ortega-Martinez, J. Aronowicz, P. Oliveri, R. R. Copley. 2014. Phylogenomic analysis of echinoderm class relationships supports Asterozoa. Proceedings of the Biological Society, 281(1786)
































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}