
Simplified cladogram of Archosauria.

More detailed phylogeny of the Ornithodira, with concentration on the early dinosaurs.

I. Origin of the Dinosaurs
Under the shadow of their crurotarsan cousins, a group of
small archosaurs evolved. The Ornithoidira
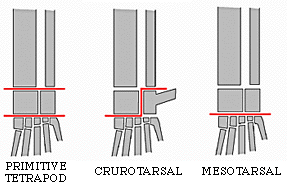
("bird necks") differed from typical diapsids by having elongate tibiae and metatarsi, suggesting
that they were even faster than typical reptiles. All (except perhaps for the most primitive) had
cervical vertebrae greatly modified so that they look very different from dorsal vertebrae, but
instead could allow the neck to form an S-shaped curve (hence the name of the group).
One branch of the ornithodirans that appeared in the Late Triassic are the Pterosauria: the first group of powered fliers among the vertebrates. We'll look more at the pterosaurs in the third section of the course.
The other major branch of ornithodirans are the dinosauromorphs. The oldest dinosauromorphs are known from the Middle Triassic of Argentina. The taxa modified the hindlimbs further to evolve:
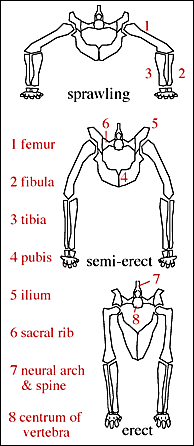
The combination of these traits allowed the little dinosauromorphs to run and accelerate in a bipedal mode all the time, not just at top speeds like typical diapsids. (It is not yet certain if the earliest dinosauromorphs such as Lagerpeton and Marasuchus were primarily quadrupeds or bipedal.
The parasagittal gait and hinge-like ankle also allowed dinosauromorphs the ability to move more actively and constantly rather than only in short bursts of speeds; thus, they were striders. Additionally, although early dinosauromorphs were small (~30 cm long for Marasuchus, 1-2 m long for "silesaurs" and basal dinosaurs, etc.), the presence of limbs directly underneath the body meant that this lineage to grow to much larger size than any previous clade while still remaining terrestrial and mobile (sprawlers relagated to a semi-aquatic life if they became too big to support their weight).
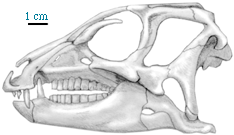
The largest dinosauromorphs other than dinosaurs are the recently discovered "silesaurs." These were quadrupedal herbivores about 2 m long. They and dinosaurs share the specialization of elongate pubes and ischia (which may have aided in increased respiratory ability: more air per breath). "Silesaurs" (not officially named yet) had been known from teeth and bones of the Middle and Late Triassic from around the world, but had been thought to be from primitive dinosauromorphs, ornithischians, sauropodomorphs, and/or theropods until the more complete specimens of Silesaurus itself let people know what this group looked like.
II. Dinosaur Groups and Relationships
Simplified cladogram of Dinosauria

"Silesaurs" are the closest relatives to Dinosauria, but they do not seem to be true dinosaurs: that is, they lack the shared derived features expected in the concestor of Iguanodon and Megalosaurus and its descendants. Those features (the shared derived features of Dinosauria) are:
(Parenthetically, at least some researchers suggest that "silesaurs" are a para- or polyphyletic group, and that some of them may actually be basal members of Ornithischia and thus true dinosaurs.)
Early dinosaurs were all around 1-2 m long bipeds with grasping hands. Footprints that might come from dinosaurs are found in the Middle Triassic of Argentina, but these may be from early members of the dinosaur lineage that evolved before the origin of Dinosauria proper.
The oldest known definite dinosaurs are from about 228 Ma, at the very beginning of the Late Triassic Epoch. The very oldest dinosaurs are from Argentina (the Ischigualasto Formation) and Brazil (the Santa Maria Formation), and new unnamed forms from India; fossils from the same age from Europe, North America, and elsewhere have turned out to be non-dinosaurian.
Dinosaurs divide into two major clades:
Members of both clades are present in the Ischigualasto and Santa Maria Formations.
Simplified cladogram of Ornithischia:

Ornithischians remain very rare throughout the Triassic. The oldest and most primitive is Pisanosaurus of the Ischigualasto Formation. The fossil is incomplete, so many apsects of its anatomy are uncertain. But it definitely shares the following transformations with later ornithischians:
Based on their tooth form, ornithischians were herbivorous. (That doesn't mean that they were exclusively plant eaters, of course! In the modern world, many "herbivorous" reptiles and mammals eat some meat.)
Later in the Late Triassic, new sorts of ornithischians appear. One of these is the newly discovered Eocursor of South Africa. Another is the long-lasting clade Heterodontosauridae. These groups share with later ornithischians the following transformations:
Heterodontosaurids were a primitive group of ornithischians that appear in the later Late Triassic, diversify in the Early Jurassic, and persist into the Early Cretaceous. They had skulls which are relatively deep and powerfully built, indicating that they ate fairly tough food. They also had premaxillary tooth row that were ventral to the maxillary tooth row and jaw joints that were ventral to the dentary tooth row: the result were jaws that brought the teeth together all at once (like a nutcracker), and not slicing (as in scissors, or as in most dinosaurs). These latter jaw adaptations evolved convergently in Ornithopoda, and so for a long time Heterodontosauridae was considered a clade within Ornithopoda.
Most astonishingly of all, it has only recently been discovered that at least the early Late Jurassic Chinese heterodontosaurid Tianyulong had a fuzzy body covering over at least part of its body! If this is found to be homologous to the protofeathers of coelurosaurian theropod saurischians it would suggest that the concestor of all dinosaurs was fuzzy, and that dinosaurs were thus fuzzy ancestrally! At present, however, there is enough uncertainty to make the homology between Tianyulong's fuzz and coelurosaurian protofeathers suspicious. (But do not be terribly surprised if in the future we discover that most dinosaurs were fuzzy to some degree or another!) (By the way, the initial reports placed Tianyulong in the Early Cretaceous, but the formation in which it was found has been redated to the earliest part of the Late Jurassic, around 160 Ma.)
Ornithischians more derived than Heterodontosauridae and Eocursor had greatly reduced hands, losing most of their grasping ability. This suggests a switch to jaws-only while obtaining food.
The remaining ornithischians includ the armored Thyreophora and the highly diverse Neornithischia (especially beaked Ornithopoda and ridge-headed Marginocephalia). We will explore the adaptations and diversity of these groups in the next three lectures.
Simplified cladogram of Saurischia:

Saurischians are named after a primitive trait ("lizard hips" = "forwards-pointing pubis"; the ancestral condition), but they are in fact united by some shared derived traits. These include modifications of the snout and of the vertebrae: however, these are rather technical features and we're not go into them in detail in this course. Additionally, some of the traits long thought to be basal saurischian traits (including some in the textbook) are now popping up in early ornithischians and in silesaurs, and so are not actually saurischian shared derived characters.
The saurischians were more common in the Late Triassic than their ornithischian sisters. The most primitive saurischians are the carnivorous Eoraptor of the Ischigualasto Formation and the Herrerasauria (the latter including Herrerasaurus of the Ischigualasto Formation and Staurikosaurus of the Santa Maria). These early saurischians resemble Theropoda (and carnivorous curotarsans) by having blade-like serrated teeth. Furthermore, herrerasaurs share with theropods an intramandibular joint between the dentary and post-dentary bones: this may have served as a shock absorber will feeding on live prey.
Some paleontologists regard Eoraptor and/or Herrersauria as primitive theropods. However, an alternative hypothesis (and the one shown here) is that these groups were simply early branches of Saurischia, and their similarities to Theropoda are either retention of primitive features or convergences. Instead, Theropoda and herbivorous Sauropodomorpha are united as Eusaurischia, based on the following transformations lacking in Eoraptor and Herrerasauria:
The most primitive sauropodomorphs have recently been suggested to form a clade Guaibasauridae. One of the most complete of these, and the oldest, is Panphagia from the Ischigualasto Formation: it is thus a contemporary of (and possible food source for) Eoraptor and Herrersaurus. Slightly younger is Saturnalia of the Santa Maria Formation. In the next younger unit above the Santa Maria (the Caturrita Formation) another basal sauropodomorph Guaibasaurus has been found. As Sauropodomorpha is the sister group of Theropoda and is present by the time of the Ischigualsto, there must have been theropods present at that time that we have not found (unless Eoraptor and/or Herrerasauria really are theropods).
We will explore sauropodomorph and theropod diversity in weeks to come.
(A special note: Owen's Dinosauria traits re-examined: If we look at some of the traits that Owen used to recognize Dinosauria, we now find that some were actually inherited from pre-dinosaurian ancestors (e.g., parasagittal stance, from the early dinosauromorphs) and others evolved convergently between Iguanodon, Hylaeosaurus, and Megalosaurus (giant size; more than two sacrals). Still, even though the traits to recognize them have changed, we still use the name "Dinosauria" to unite these reptiles.)
III. Dinosaurs in the Late Triassic
Although dinosaurs appeared in the early part of the Late Triassic ecosystems, they were just a minor part.
For example, other animals were far more common in the Ischigualasto Formation than Pisanosaurus,
Eoraptor, Herrerasaurus, and Panphagia. Dinosaurs (either herrerasaurs or theropods) were not the
top predators in any known Late Triassic assemblage: that distinction goes to big crurotarsans. By the end
of the Late Triassic, sauropodomorphs HAD become the largest and most common herbivores in some ecosystems (away
from the equator).
Recent work shows the earliest dinosaurs and the earliest members of most dinosaur lineags showing up in the southern continents (especially South America and southern Africa) first. In fact, ornithischians are not known outside of these regions until the Early Jurassic: their niche in North America, Europe, etc., is exploited by "silesaurs" and/or herbivorous crurotarsans instead. New discoveries from the lower part of the Chinle Formation of the American Southwest shows that diverse non-dinosaurian dinosauromorphs were still a major part of the assemblage well into the later part of the Late Triassic.
So the Late Triassic is the time WHEN DINOSAURS SHARED THE EARTH!. At the end of the Late Triassic, though, there was a mass extinction (the Triassic-Jurassic Extinction). We will explore the details and causes of it at the end of the second section of the course. For now, though, we'll point out that this event wiped out:
With no serious competition remaining, dinosaurs radiated into numerous forms. From the beginning of the Early Jurassic onward, DINOSAURS RULED THE EARTH.
To Next Lecture.
To Previous Lecture.
To Syllabus.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}