


Key Points:
•Dinosaurs of all clades laid eggs in nests. Recent discoveries have shown that some dinosaur eggs were leathery, while others were crispy.
•Some dinosaur nests seemed to have been covered with vegetation; others were open to the air..
•In the pennaraptorans one or more parent brooded directly on top of the eggs to keep them warm.
•Baby dinosaurs were mostly very small relative to the adult body size.
•Compared to placental mammals, dinosaurs reproduced huge numbers of young. This indicates that chance of survival of any individual baby is very small.
•We can estimate the age at the time of death in dinosaurs by skeletochronology. It has been found that dinosaurs had rapid growth rates, reaching full body size in less than a decade for most groups, and less than two decades for even the largest.
Up until 2020 (freaking 2020, man... It changed everything...) another such trait was "Eggs with a hard calcium carbonate shell (as opposed to the leathery shells of lepidosaurs, turtles, egg-laying mammals)". Crocodilians have this trait, birds had this trait, and fossil dinosaur eggs (from titanosaurs, hadrosaurids, and a diverse assemblage of tetanurines including megalosauroids, carnosaurs, alvarezsaurs, therizinosaurs, oviraptorosaurs, troodontids, dromaeosaurids, and basal avialians) were known to have this. So it looked like a simple phylogenetic inference that all dinosaurs (indeed, all archosaurs) had this trait ancestrally.
One archosaur group shown not to have this was Pterosauria, which had the parchment-style egg shell. But this was best explained as a reversal.
Than along comes 2020. It is shown that the eggs of Protoceratops were in fact parchment style, and a paraphyletic series of basal sauropodomorphs also show a parchment style. So the state at the base of Dinosauria, and Ornithodira, and Archosauria is either ambiguous (using parsimony) or parchment (using Bayesian inference). If the latter, than the crispy shells of Crocodylia, Hadrosauridae, Titanosauria, and Tetanurae all evolved convergently.
Fossil dinosaur eggs and nests found in the rock record, as have embryos of most major dinosaur clades.
ALL non-avian dinosaur eggs are basketball-sized or smaller: NO dinosaur hatched from eggs the size of people!!
Some dinosaur nests associated with covered mats of vegetation: probably helped to keep warm (as in croc nests).
Some dinosaurs (pennaraptorans) found in "brooding position" over nests; unlikely to be found in dinosaurs which are too large (i.e., tyrannosaurids, hadrosaurids, sauropods, etc.) or lacking feathers. A recent study of the density of pores on archosaurian eggshells shows that most dinosaurs have characteristics of eggs from covered nests, but that pennaraptorans show open nests. This is strongly consistent with the record of brooding.
In primitive modern birds it is the male rather than the female which broods the nest: paternal care. These nests are laid by multiple females. In these paternal care cases, the male rather than the female typically watches over the young after they hatch. When plotted against body size, the volume of eggs of nests of dromaeosaurids, troodontids, and oviraptorosaurs more closely resembles the pattern seen in paternal caring birds than in maternal caring or biparental caring birds; this suggests that paternal care may be an ancestral (pennaraptoran?) trait that was passed on to primitive avians but modified in more derived birds. However, we do not have evidence yet to see if more basal dinosaurs had paternal care, maternal care (as in crocodilians), or biparental care. And indeed the statistics supporting the paternal care hypothesis in non-avian pennaraptorans has been called into question.
Most primitive modern birds are ground nesters; suggests that tree nesting did not evolve until well into the modern bird (Aves) radiation.
Dinosaurs tend to have nests of about a dozen or so eggs each: more than found in modern birds, less than in (for example) turtles. This is regardless of size: troodontids to titanosaurs! This implies that unlike placental mammals, dinosaurs could produce a dozen or so offspring a year regardless of size; among placental mammals, larger body size means LONGER gestation periods.
Two main potential life habits upon hatching:
Some evidence of these habits in hatchling dinosaurs:
Parental care of babies for at least several weeks is present in both modern crocodilians and modern birds, implying that this trait was present in archosaurs ancestrally. Birds have long been known to provision (bring food to) their babies; there are cases of crocodilians in captivity doing the same (see video below), but it is not yet known if they do this in the wild. But certainly crocodilian mothers bring their babies to safer places in ponds and lakes where they little ones could hunt.
After baby dinosaurs left the nest, at least seem to have travelled together. Packs/herds of of baby ankylosaurids, ceratopsians, and ornithomimosaurs are known; in at least the case of the ceratopsian Psittacosaurus and the ornithomimosaur Sinornithomimus these herds can include individuals of different ages, including adults. (The parents may have been with the other baby herds, but were too big to be buried. So it isn't entirely certain that the "young only" herds are due to that being the actual social structure: it might be the byproduct of taphonomy.)
Dinosaur Growth Rate and Skeletochronology
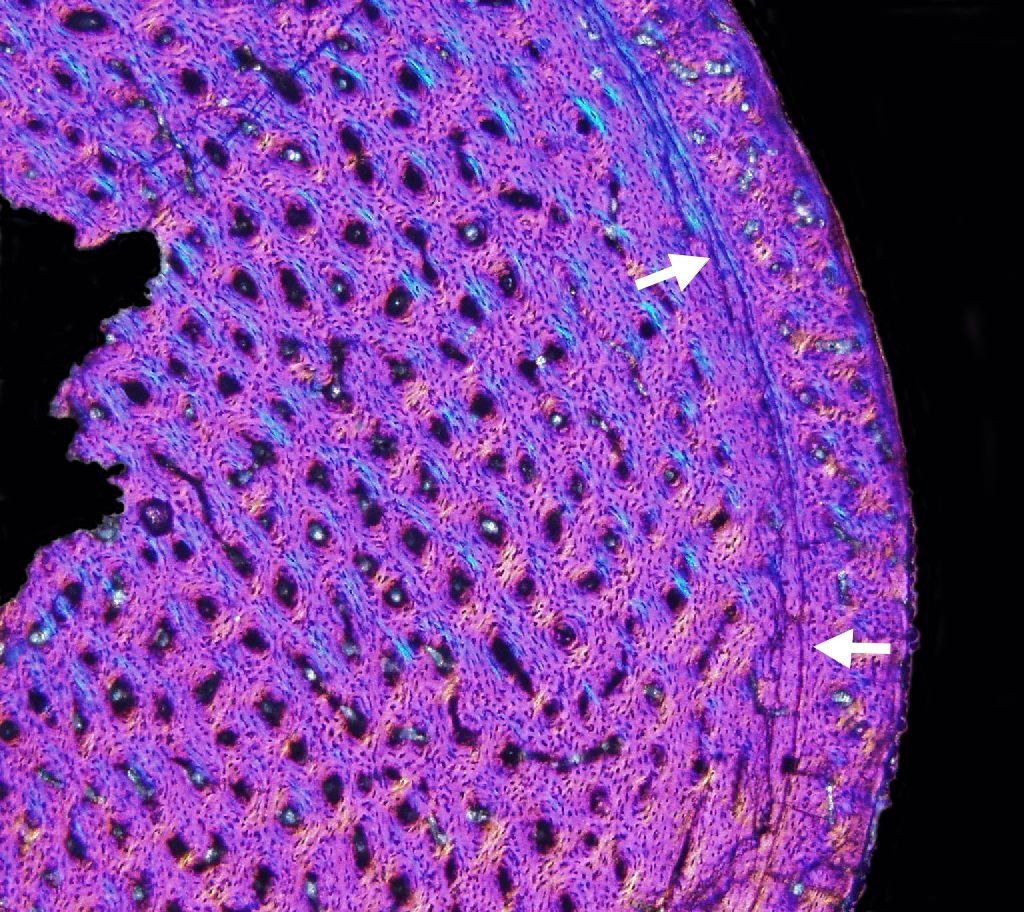
Like many animals, dinosaurs deposited Lines of Arrested Growth (LAGs for short). These are basically growth rings like tree trunks, one per year. (Okay, technically speaking the broad term is cyclical growth marks (or CGMs). This includes rings (which show temporary cessation of growth) as well as more diffuse annulus marks (which simply show slow downs.) In practice, we typically just use the acronym "LAG".) Various paleontologists use that information to "age" individual dinosaur fossils. By comparing the age of the dinosaur to its size, and plotting all individuals of the species on a graph, you can figure out the growth pattern and growth rate of that dinosaur species: that is, we can develop a skeletochronology for some species. (Sauropods are particularly difficult to deal with, as excessive reworking of their bones obscures their CGMs.)
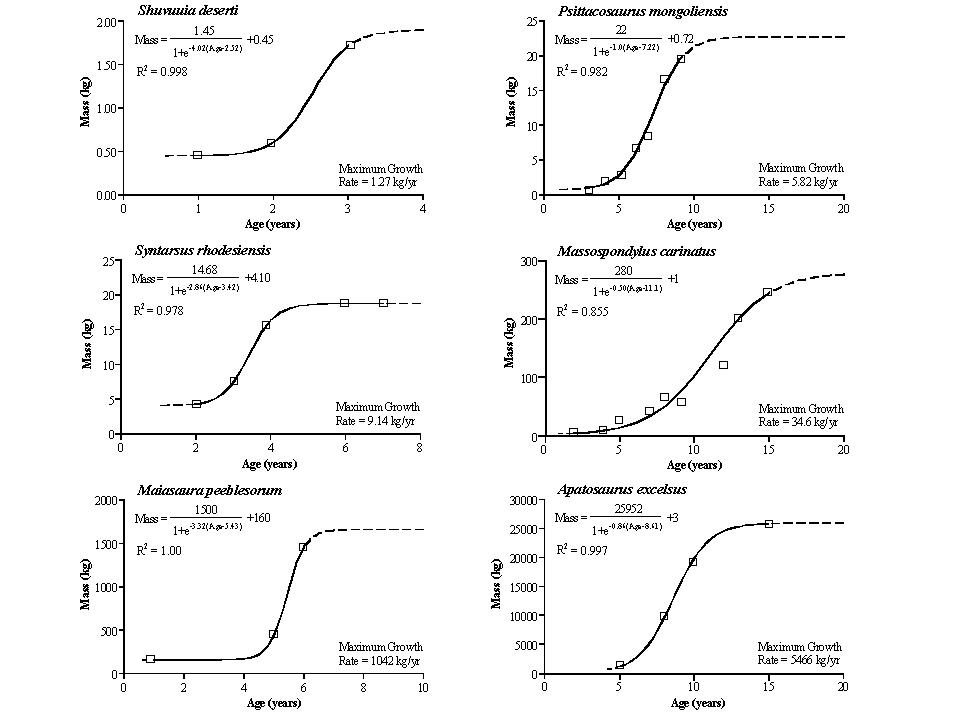
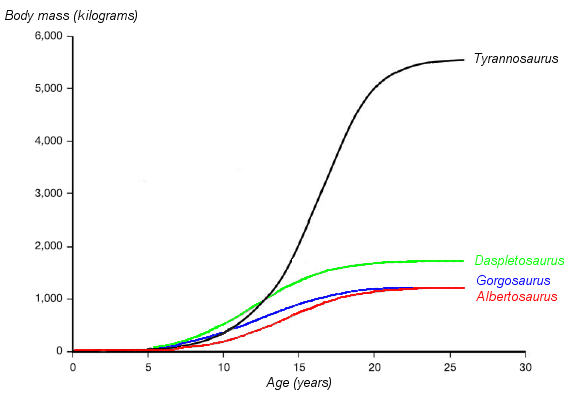
Actual rate was VERY HIGH compared to typical reptiles, particular in big dinosaurs. Estimates based on bone "growth rings" (and other features) indicate small dinosaurs were full grown at only a couple of years, hadrosaurids (e.g. Maiasaura) at only 7 years for, and only 15-20 years for some big sauropods (e.g., Apatosaurus) and some big theropods (e.g., Tyrannosaurus) to reach adult size. In other cases it might be longer: the carcharodontosaurid theropod Meraxes may have taken about 30 years to reach adult body size, and the largest specimen giant diplodocoid sauropod Diplodocus hallorum seems to have died at about age 60 but it is uncertain how long it was at full body size.
(In contrast, big crocodilians from the Late Cretaceous seem to have taken 50 years or so to reach the same size as big hadrosaurs).
Since most animal populations stay generally stable over time, more baby dinosaurs died before reaching adult size than in typical populations of modern birds or mammals (imagine herd of antelope where every female produced a litter of 12 every year!).
Unlike typical non-avian reptiles, dinosaurs seem to have determinate growth: reaching a fully adult size, then stopping growth (or at least slowing it WAY down: all the LAGs bunch up together to form an external fundamental system (or EFS).
Lifespans for dinosaurs mostly seem to be shorter than those of similar sized mammals. For instance, the oldest known Tyrannosaurus individual is only 28-33 and the oldest Camarasaurus about 38, but elephants can live to 70 or more. There are dinosaurs with elephantine lifespans, though: the oldest carcharodontosaurid known died around age 50 or so, the oldest Diplodocus at about age 60, and the oldest Supersaurus was so old at time of death that the traces of its growth are obscured.
A Note of Caution: in at least one living bird (the North Island brown kiwi Apteryx mantelli) the number of LAGs found in individuals of known age underestimated the actual age at time of death. It is noteworthy, though, that kiwis are rather peculiar birds, having undergone some substantial size and metabolic rate decreases in their ancestry from the ancestral modern bird condition, and this might be reflected in this unpredicted situation. Additionally, studies of some small living amphibians, lizards, tuataras, and mammals found that the LAG count and the age didn't always match. So counts of LAGs can be used as a minimum age of the animal, but it might not be the actual age.
Life-History Strategies
Different types of organisms have different life history strategies: different ways of dealing with how many young are born at a time; how many survive to adulthood; how long they live; etc. There are two general types of strategies that we see commonly in nature:
Non-avian dinosaurs are definitely more r-selected than typical larger placental mammals, but more K-selected than sea turtles.
To Next Lecture.
To Previous Lecture.
To Lecture Schedule.

{kind=link}
{kind=link}
{kind=link}
{kind=link}