

Key Points:
•Dinosauria first appears by early in the Middle Triassic. Many (although not all) of the oldest dinosaurs are from the southern parts of Pangaea, but this may be due to a sampling bias.
•Early dinosaurs were relatively rare in the Middle and early Late Triassic. But around 234-232 Ma a major environmental event--the Carnian Pluvial Event--seems to have favored dinosaurs over their competitors
•The Triassic ends with a major mass extinction--the Triassic/Jurassic Extinction--triggered by the eruption of the Central Atlantic Magmatic Province and the breakup of Pangaea. This event wiped out most of the dinosaur's rivals, and allowed them to inherit the Earth
I. Dinosaurs in the Triassic
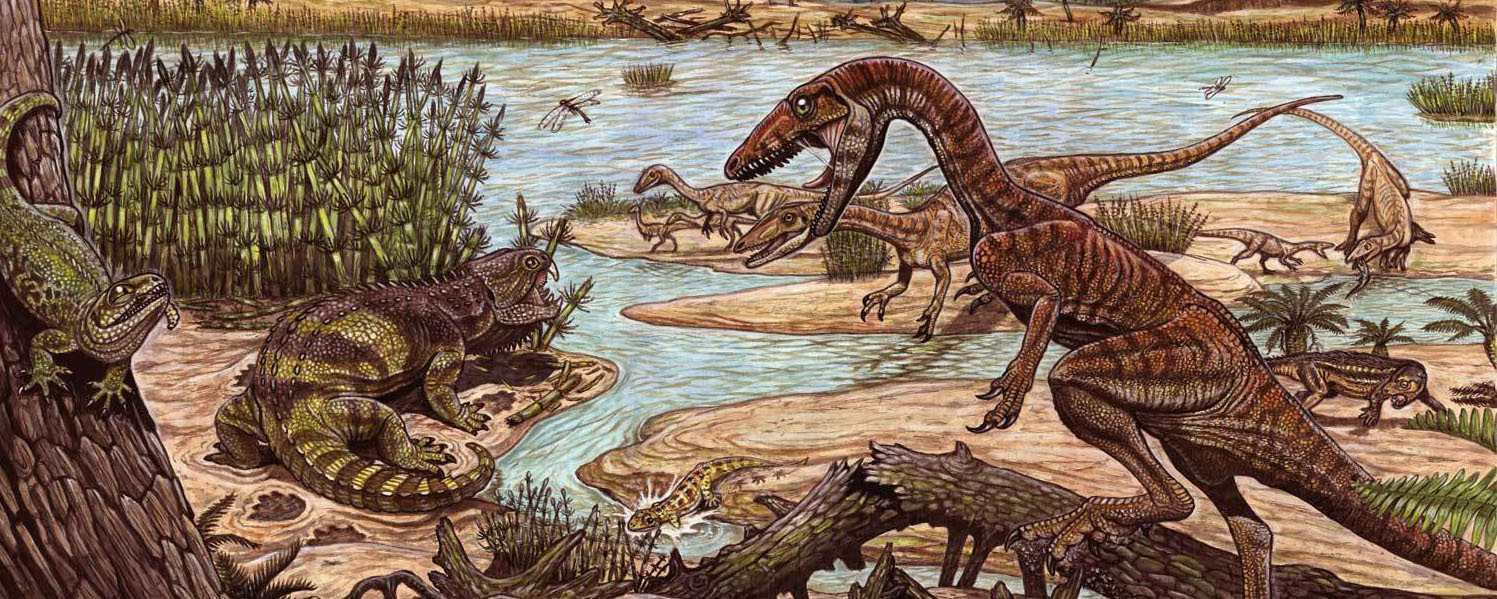
If "silesaurs" are indeed diniosaurs, the oldest known definite dinosaurs are from about 247 Ma (and just appeared shortly after the Permo/Triassic Mass Extinction), and show up first in South American and Africa. However, even by the very beginning of the Late Triassic Epoch (when non-silesaur dinosuars are definitely present) they were rare. The very oldest dinosaurs are from Argentina (the Ischigualasto Formation), Brazil (the Santa Maria Formation), Zimbabwe (the Pebbly Arkose Formation), and from India (the Maleri Formation); fossils from the same age from Europe, North America, and elsewhere have turned out to be non-dinosaurian. Because of this, it has been argued that southern Pangaea was the ancient homeland of Dinosauria, and that the group spread northern (as the arid zones decreased in wetter times) in the later Late Triassic.
Although dinosaurs appeared in the early part of the Late Triassic ecosystems, they were just a minor part. For example, other animals were far more common in the Ischigualasto Formation than Eoraptor, Herrerasaurus, Panphagia, Pisanosaurus and the like. Theropods were not the top predators in any known Late Triassic assemblage: that distinction goes to big pseudosuchians. By the end of the Late Triassic, sauropodomorphs HAD become the largest and most common herbivores in some ecosystems (away from the equator). And by the lower part of the Chinle Formation of the American Southwest shows that silesaur-grade ornithischians and primitive saurischians a major part of the assemblage well into the later part of the Late Triassic.
So most of the Late Triassic is the time WHEN DINOSAURS SHARED THE EARTH!. Pseudosuchians remained ecologically diverse and important, and (especially in the early Late Triassic) non-archosaurian archosauromorphs and therapsids were also significant.
But by the end of the Late Triassic dinosaurs were far more common, and many of the earlier diapsid reptiles were rare or extinct. Was this due to dinosaurs out-competing them ("good genes") or was it due to extrinsic changes in the external environment ("good luck")? Or maybe a bit of both?
We will explore in later parts of the course the evidence that dinosaurs had elevated metabolisms relative to most of their contemporaries. This would allow greater and more sustained activity during the day, but would come at the cost of requiring much more food. Traditionally the fully upright posture of dinosaurs was used as evidence of their locomotor superiority to their contemporaries, but it is noteworthy that many of the most successful late Late Triassic pseudosuchians (such as paracrocodylomorphs) ALSO had a parasagittal stance.
Studies of the morphological diversity shows that pseudosuchians actually remain the most diverse group of medium-to-large terrestrial amniotes all the way until the end of the Triassic. So even though dinosaurs were diversifying, they were still relatively less important parts of the ecosystem. So an alternative to the superiority model is that environmental changes may have resulted in dinosaur success.
II. The Carnian Pluvial Event & the Triassic/Jurassic Mass Extinction: Dinosaurs Inherit the Earth
In the later Late Triassic (in Gondwana and Europe, at least) things shift: pseudosuchians remain important (especially in the form of paracrocodylomorphs, aetosaurs, and phytosaurs) as did dicynodont therapsids, but sauropodomorphs expand in diversity (and size!). In recent years it has been suggested that a major environmental shift was the cause of this. Sedimentological analysis shows that the world got considerably wetter from 234-232 Ma. Its particular trigger isn't well understood, but it affected life on both land and sea. This event occurred in the Carnian Stage, and so is called the Carnian Pluvial Event ("pluvial" = "of or concerning rain"). One of the changes on land is the diversification and spread of groups of tree-sized plants (especially conifers and the now-extinct cycadeoids). It has been argued that the accumulation of this new leafy biomass may have promoted the expansion of the sauropodomorphs, and thus of Dinosauria.
(That said, the phenomenon is NOT apparent in North America! Both the excellent body fossil record of the Southwest and the trackway record of the East show no spread of sauropodomorphs, or dinosaurs in general, in North America in the later Carnian and subsequent Norian and Rhaetian stages.)
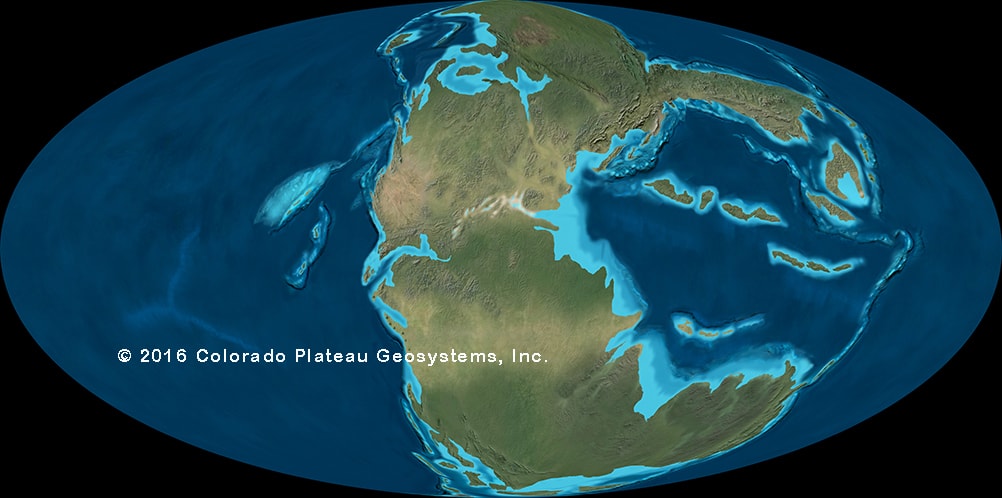
At the end of the Late Triassic, though, there was a mass extinction that affected the entire planet: the Triassic-Jurassic Extinction. The primary trigger seems to have been a period of tremendous volcanism in the middle of Pangaea. 201 Ma saw the beginnings of the rupture of the supercontinent of Pangaea, between the landmasses that are today North America, Europe, and Asia (on the north) and South America, Africa, and the other continents (to the south). This produced the Atlantic Ocean in the long term, but for the inhabitants of the Earth at the time suffered.
A vast amount of greenhouse gasses (both carbon dioxide and methane) were released, producing catastrophic global warming. But this was preceded by a burst of sulfur dioxide, which produced a rapid phase of cooling. There is additional evidence that poisons like mercury were released into the environment.
These massive pulses of environmental change were too much for many groups of organisms to survive. This is one of the five greatest mass extinctions known, with many terrestrial and marine taxa wiped out. Among terrestrial vertebrates, some of the main victims were:
A recent (2022) study postulated that it was the intense cold which was the primary factor influencing which groups survived and which thrived at the extinction among terrestrial amniotes. In particular, the authors speculate that the higher metabolic rates and possible insulation of dinosaurs and pterosaurs (relative to pseudosuchians) allowed them to make it through this event.
With no serious competition remaining, dinosaurs radiated into numerous forms. From the beginning of the Early Jurassic onward, DINOSAURS RULED THE EARTH

{kind=link}
{kind=link}
{kind=link}