

Key Points:
•Paleoecology is the study of the factors which controlled the abundance and distribution of fossil organisms.
•Some paleoecological work examines the life habits of individual species (autecology); others look at the assemblage of organisms that existed at one place and one time (community ecology or synecology).
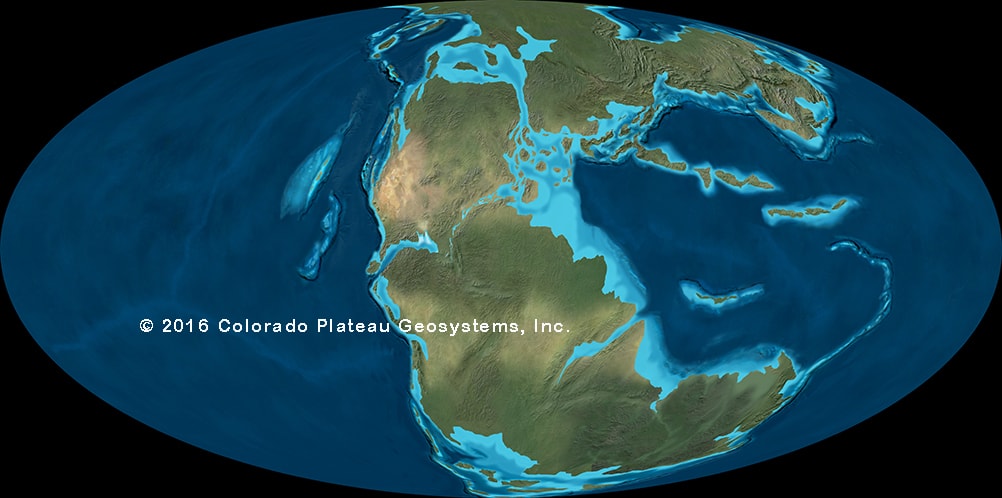
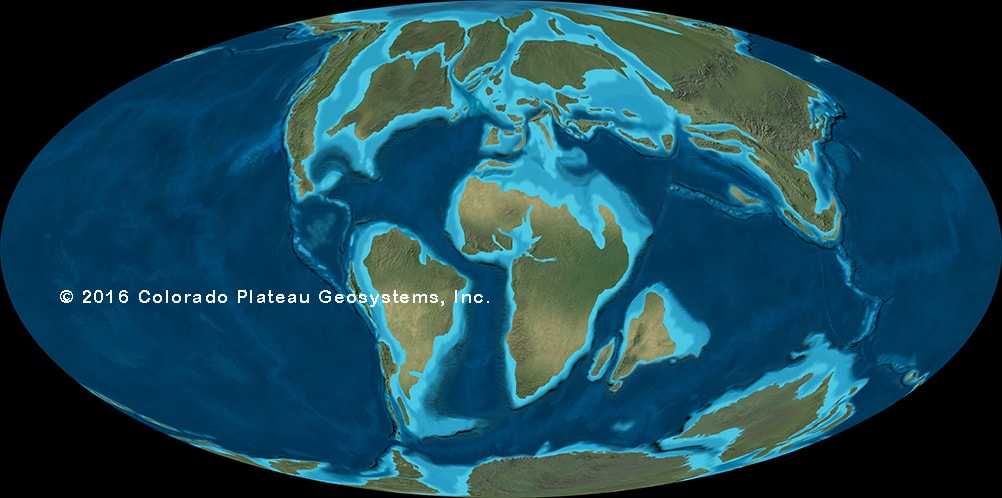
•The large scale pattern of dinosaur history is controlled by the break-up of Pangaea. Earlier dinosaur faunas in different parts of the world are very similar to each other; as time went by, the separation of land masses allowed distinctly provincialized faunas to evolve.
•Different groups of dinosaurs co-existed at different times and places.
Here are some important terms concerning paleoecology:
Example of Autecology: Migration in Camarasaurus
The isotopic composition of water varies from spot to spot due to temperature, evaporation, and other factors. In a study geochemist Henry Fricke and colleagues compared the isotopic composition of clay minerals and the teeth of the macronarian sauropod Camarasaurus from several sites in the Morrison Formation (Late Jurassic, western North America). While the clays only had the isotopic composition of the lowland environment (not surprisingly, since that's where they formed and that's where the teeth were found), but the Camarasaurus teeth show a broader spread of values, including upland signatures. Looking within individual teeth, it was found that some succession of layers show lowland values and then swtich to show upland values. This has been interpreted as indicating migration between uplands and lowlands
within the species.
Another Example of Autecology: Infection in Tyrannosaurus
Many specimens of Tyrannosaurus (and other tyrannosaurids) show lesions in the lower jaw. These were first considered to be
bitemarks from other tyrant dinosaurs, but closer examination reveals that these are not punction marks, but resorbed bone.
In fact, their closest analog in the modern world are the lesions produced by avian trichomonosis (infections in birds from the protozoan Trichomonas gallinae). Modern birds of prey get this infection primarily from eating infected prey, and so it is quite likely that the tyrant infections were similarly picked up from infected food items.
Synecology of a Single Fauna: the Hell Creek (latest Cretaceous, western North America)
How do we determine the animals that make up a paleofauna? Rather simply: the animals whose fossils are found in a formation. But not all formations are equally well-explored, so while we have long faunal lists for some of these, and very few species known in others. Also, there are complicating factors:
In a recent study of the latest Cretaceous Hell Creek Formation of western North America, the distribution of different dinosaur species within different micro-habitats was examined. The researchers compared the relative occurrences of different species in sandstones (representing the rivers and near-river environments) and mudstones (which generally represented the floodplains further from the river). They found that the ceratopsians of the Hell Creek (the chasmosaurines Triceratops and Torosaurus and the leptoceratopsid Leptoceratops were more common in the upland environments; that the ornithopods (the hadrosaurine Edmontosaurus and Thescelosaurus) more common near the rivers; and that other dinosaurs did not show a statistically significant preference between the habitats (a possibility that ornithomimids were more common in the uplands, but not statistically significant). In the case of the apex predator of the environment, Tyrannosaurus, this lack of preference has strong statistical support: this makes sense, because meat is meat, so its food base would be found in both settings. In contrast, however, the particular plants favored by ceratopsids and ornithopods may have been linked to their favored micro-habitats.
(Neither the oviraptorosaur Anzu nor the dromaeosaurids Acheroraptor and Dakotaraptor were known when this study was done.)
Few other formations have been subjected to this level of analysis, but hopefully this will be done in the near future.
Inter-faunal Comparisons: Late Jurassic Sauropod-Dominated Communities
One can compare faunas to faunas just as we can compare micro-habitats to micro-habitats. In a recent study, different sauropod-dominated communities of the Late Jurassic from around the world were compared. These sites were from different formations on different continents representing different habitats of the Late Jurassic.
The researchers found that arid environments were dominated by high-browsing macronarians; humid environments by lower browsing forms (including diplodocoids, stegosaurs, and ornithopods); and no surprisingly semi-arid environments had a mixture of these. Once again, the carnivores were largely independent of the different types of environment. (This isn't really that much a surprise: consider that leopards, tigers, mountain lions, and (in the historic past) jaguars and lions range from tropical jungles through arid plains to cold forests, while the herbivores they feed on are often restricted to just one habitat.)
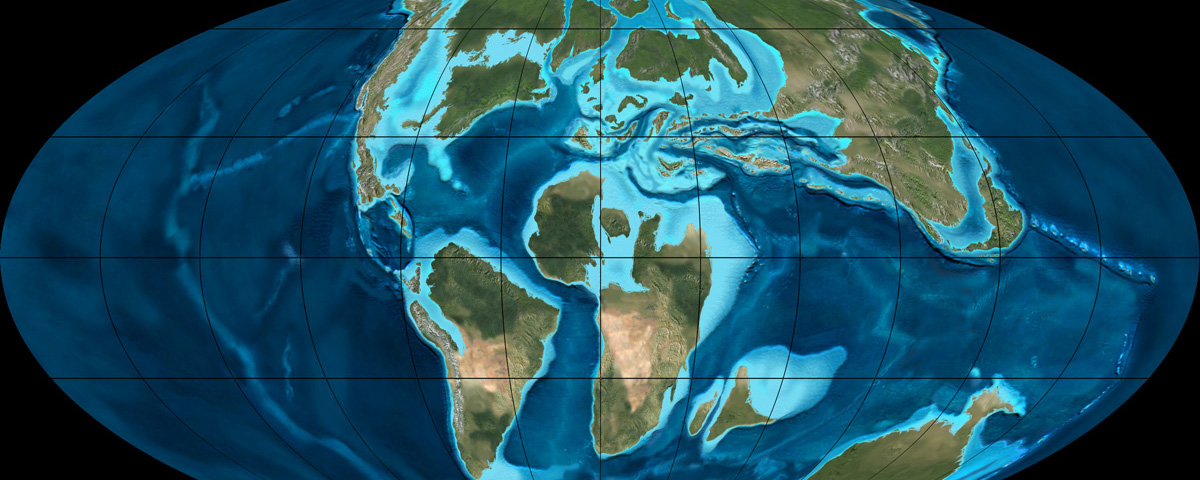
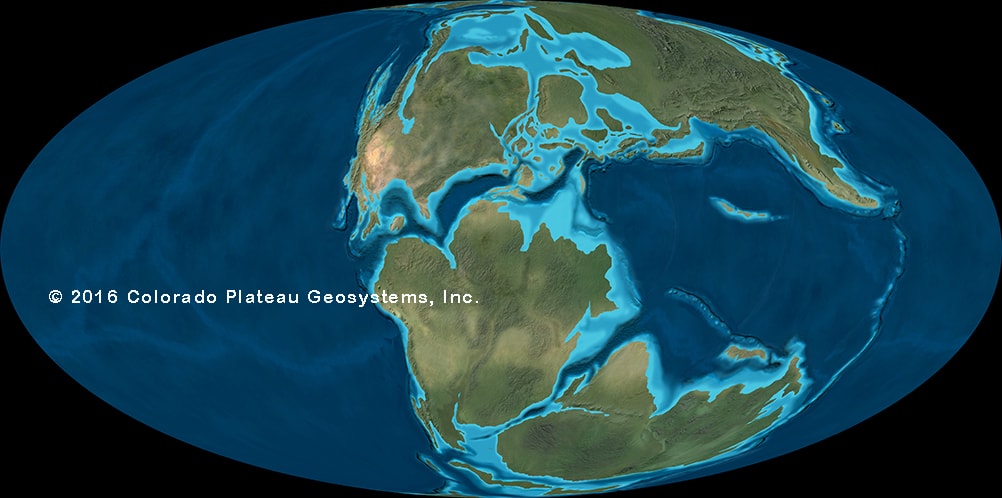
History, Extinction, and Paleobiogeography: the Many Worlds of the Late Cretaceous
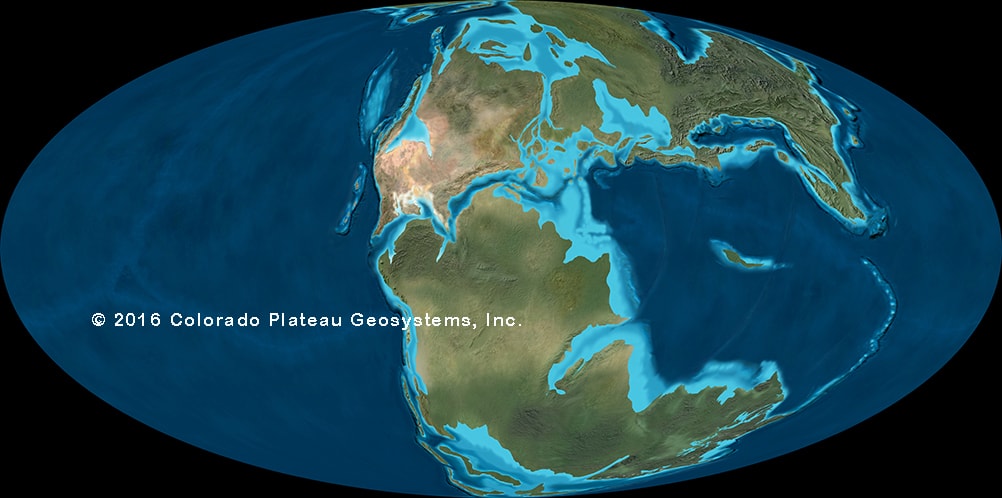
For much of dinosaur history, the same dinosaur groups were found in nearly every continent: that is, the faunas were cosmopolitan. This makes sense for the early part of dinosaur history, when the landmasses were still joined as Pangaea. But even with the birth of the Atlantic at the Triassic-Jurassic boundary, the spreading of the Atlantic during the Middle and Late Jurassic, and the break up of the southern continents (Gondwana: today's South America, Africa, Antarctica, Australia, India, and Madagascar) and the rise of higher sea levels during the early Early Cretaceous and late Early and early Late Cretaceous, there is still a generally shared pattern of the types of dinosaurs found worldwide. Different groups come and go, but they do so around the world:
The later part of the Early Cretaceous and the beginning of the Late Cretaceous (basically, the block of time between about 120 and 85 million years ago) is a time of relatively high oxygen levels, fairly high carbon dioxide values, high sea levels, and high temperatures (even for the Mesozoic). This produced a climate that was nearly tropical from pole-to-pole. Some of the largest dinosaurs of all are from this time (sauropods like Sauroposeidon, Argentinosaurus and Paralititan; theropods like Giganotosaurus, Mapusaurus, Carcharodontosaurus, and Spinosaurus): it may be that the conditions were favorable to high plant productivity (more food made per unit time), which could support more herbivores, which could then support more carnivores.
During this time several equatorial dinosaurs evolved tall fins from their neural spines. This may have served as a means to dump excess heat and cool off.
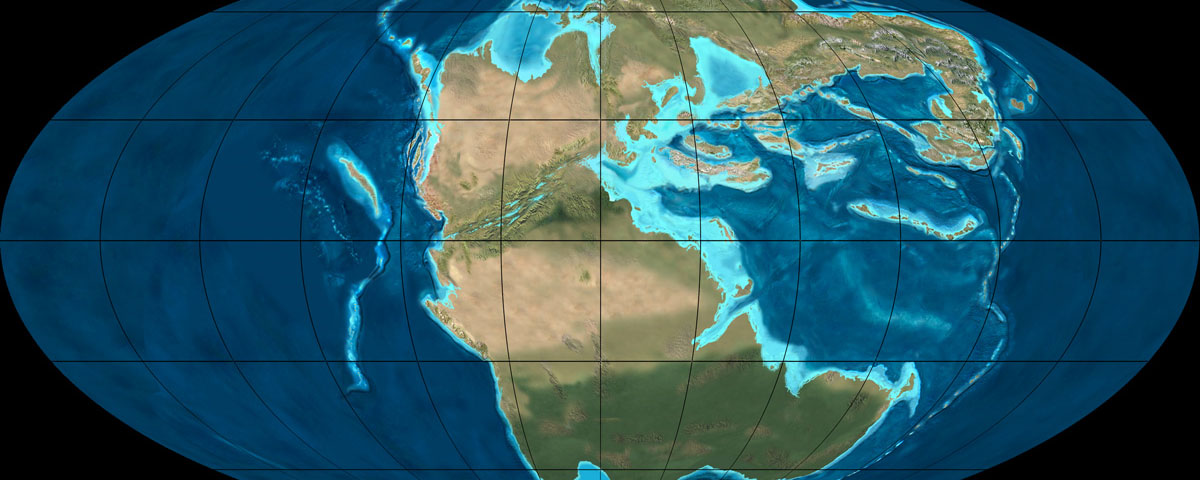
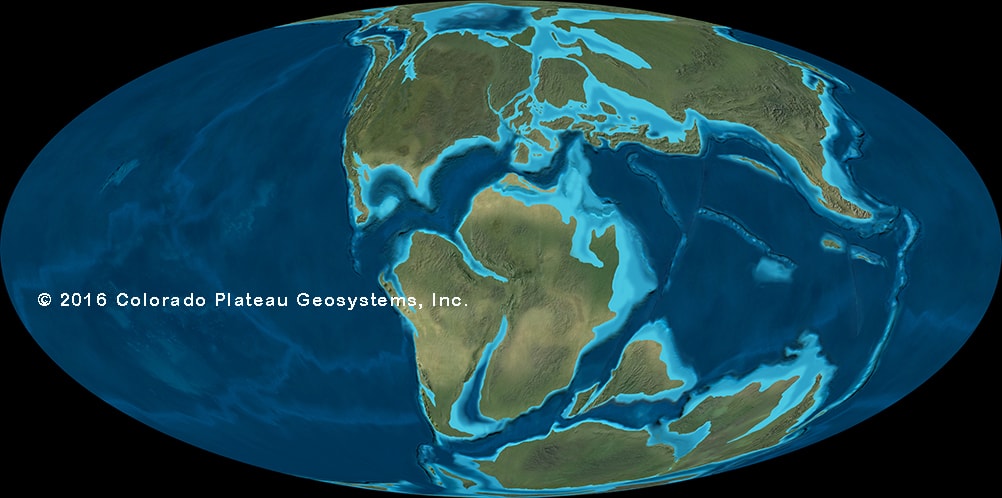
Laurasian (Europe, Asia, North America) and Gondwanan dinosaurs of this interval became even more distinct. While some groups were shared (ankylosaurs; iguanodontians; titanosaurs; spinosaurids; carcharodontosaurids; megaraptorans), others were more provincial. The odd regions out seems to be parts of Laurasia. In North America, ceratosaurs and spinosaurids have yet to be confirmed. In Asia (in particular) the ankylosaurines, the neoceratopsians, and many diverse coelurosaurs (tyrannosauroids, ornithomimosaurs, oviraptorosaurs, therizinosaurs, and deinonychosaurs) flourish and diversify. In Laurasia more generally, iguanodontians undergo "hadrosaurification": that is, we see the correlated progression of hadrosaurid traits within Hadrosauria.
Also at the end of this time western North America's fauna receives a number of immigrants via a landbridge from Asia: ankylosaurines, advanced neoceratopsians, advanced tyrannosauroids, and ornithomimids.
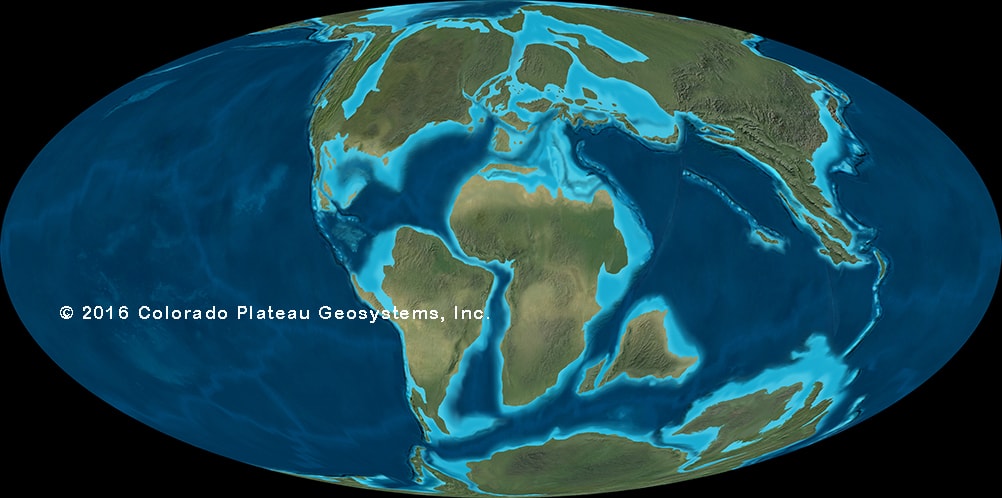
The situation changes, however, with the later part of the Late Cretaceous. Several changes occur during this time:
Why these faunal changes occurred remains to be explained. It may be related to a general cooling of the world, and perhaps a loss of food at the base of the food chain (which supported the largest sauropods, which supported the giant carnivores). For whatever reason, the world left in this faunal change is rather distinct: it is provincial (that is, there are distinct faunas in different region).
There are several distinct biogeographic regions during the late Late Cretaceous (85-66 Ma):
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}