Fall Semester 2018 Fossil Species and Alpha Taxonomy
Many skulls of the dire wolf Canis dirus discovered at the La Brea Tar Pits, California
Key Points:
•Species delimitation remains problematic. Several different methods have been proposed, but all have some problems in certain situations.
•Nevertheless, the idea of "species" represents a useful concept for describing biodiversity.
•A set of rules, initially established in the 1700s but modified over subsequent centuries, governs the formal taxonomic nomenclature.
•Alpha taxonomy is the core of this work: the description of specimens, identification of the same, and coining of new names if it appears to be a new taxon.
"Of what use are the great number of petrifactions, of different species, shape and form which are dug up by naturalists? Perhaps the collection of such specimens is sheer vanity and inquisitiveness. I do not presume to say; but we find in our mountains the rarest animals, shells, mussels, and corals embalmed in stone, as it were, living specimens of which are now being sought in vain throughout Europe. These stones alone whisper in the midst of general silence." -- Aphorism 132, Philosophia Botanica (1751), Carolus Linnaeus
"I am fully convinced that species are not immutable; but that those belonging to what are called the same genera are lineal descendants of some other and generally extinct species, in the same manner as the acknowledged varieties of any one species are the descendants of that species." -- Introduction, On the Origin of Species by Means of Natural Selection (1859), Charles Darwin
""The usual concept of species can be stated as follows (Mayr 1970): "Species are groups of interbreeding natural populations that are reproductively isolated from other such groups." This concept is grandly called "the biological species concept." But that is an arbitrary appropriation of a term with a more general and earlier meaning. I will instead use the term "reproductive species concept."" -- "Ecological species, multispecies, and oaks" (1991), Leigh Van Valen
"An important aspect of any species definition whether in neontology or palaeontology is that any statement that particular individuals (or fragmentary specimens) belong to a certain species is an hypothesis (not a fact). -- "Cladistic classification as applied to vertebrates" (1977), Niels Bonde.
The Species Problem, Continued
So here are things most people agree with about species:
There exist clusters of individuals closely related by inheritance...
...with a contiguous connection of propagation through space and time...
...characterized by a unique suite of traits (morphological, genetic, behavioral, etc.) distinct and diagnosable from similar clusters...
...and occupying a particular niche (of whatever breadth) in their ecosystem
The previous set of notes includes a look at various proposed species definition concepts (or more precisely, criteria: that is, the means by which species would be delimited). Each of these has two major aspects: some similarity criterion (shared aspects of members of the species) and some difference criterion (features that make the species distinct from other species.) Different species criteria emphasize these to different degrees, but each includes both. For instance, the BSC considers interbreeding (a similarity criterion) within the species, and reproductive isolation (a difference criterion) as the boundary of the species.
The previous notes also indicates some of the problems with each of the distinct species criteria. None entirely encompass the range of biological possibilities, or are either too restrictive or too inclusive compared to the species as actually recognized and used by field naturalist and other working biologists. But just to add to the difficulty, here are some additional problematic issues:
Species originating within another species: This is a particular problem for the phylogenetic species concept. We recognize that some species (indeed, likely many!) originate from subpopulations of previously existing species; the REMAINING subpopulations are free to persist under the original morphology, range, habitat, gene exchange, etc. A notable example includes a number of extinct Hawaiian geese and the surviving Branta sandvicensis (nene), which represent a cluster of new species arising from isolated populations of Canada geese (Branta canadensis), the rest of which's populations remained in the original morphology and habitat. Similarly, polar bears (Ursus maritimus) arose as isolates within the cluster of populations of brown bear (Ursus arctos).
Introgression: This is a problem for the BSC. The rise of genomic analysis has revealed that not only is hybridization in the wild not a rare phenomenon, it does not (as some BSC advocates hoped and claimed) only produce rare individuals of reduced fitness, and thus do not contribute to the adaptive success of variants within the population. Instead, it has been discovered that introgression (the repeated backcross of hybrids with their parent species, resulting in significant gene transfer from one species to another) is actually disturbingly common. Indeed, it is appearing time and time again in many, many groups of animals (it had long been recognized to be common in plants.) In just the last several years it has been documented among many types of mammals, including various species of bears, of big cats, of mysticetes (baleen whales), of proboscideans (including the survival of genes from extinct elephantids in the surviving species), and in the genus Homo. Note that in all of these but whales the studies have included recent (Pleistocene) fossil DNA, showing this phenomenon is on-going. And in at least the human case, the Neanderthal genes introduced into some modern human populations code for traits with positive adaptive significance: that is to say, individuals that are descendants of these hybrids have the potential for increased selective success compared to other individuals.
Species originating by hybridization: This introgression pattern achieves greater expression in entire species which arise by hybridization between closely related taxa. Note: this is a distinct phenomenon from the hybridogens mentioned last time: those are species where every individual has to be produced by a hybrid breeding event. These cases were initially established by hybridization, but later contained populations of interbreeding individuals with distinct morphologies and genomes from either ancestral lineage. These cases remain controversial, but two examples that seem to be reasonably well-supported are: the red wolf (Canis rufus) of Southeastern North America, with ancestry in the grey wolf (C. lupus) and either (or BOTH) the eastern wolf (C. lycaeon) and the coyote (C. latrans); and the Jefferson mammoth (Mammuthus jeffersonii), a morphologically-distinct form genetically nested among the Columbian mammoth (M. columbi) but with a substantial amount of woolly mammoth (M. primigenius) genes.
So where does that leave us?
There really are clusters of biodiversity with shared history, interbreeding, ecology, etc.
In general (barring ring species, etc.), these variation within each cluster is less than the the distance to the next such cluster
At lower levels of analysis, reticulation (interbreeding) dominates inheritance over divergence (branching)
As you move to higher level branching tends to dominate, but (as we now know) reticulations do not cut off at some arbitrary point. (Even though they are essentially absent [so far as we know] above the old-style family level, reticulations are present by rare at very high levels in the form of various sorts of endosymbiosis, as in the origins of lichens, eukaryotes, plants, etc.)
So the word of advice is that at present no species concept/criterion is perfect, so hold off on accepting them as Holy Writ!
Keep in mind, the goal of Science is to describe Nature. Modify rules to fit Nature, not Nature to fit rules.
And specifically with regards to fossils: in the end--with the rare exception of fossil genomes--all fossil species are morphospecies, since we really can't see other aspects of them. And since there is the time factor that neontologists don't have to deal with, we see stratigraphic variation which segue into chronospecies. (We'll talk more about this issue and rates of change, later.)
Alpha Taxonomy and Species Grammar
Alpha taxonomy: the discovery, description, and classification of species.
Linnaean taxonomy has its own special set of grammatical rules:
Genera have one word names (e.g., Panthera, Homo, Ginkgo, Tyrannosaurus);
The genus name is always Capitalized and italicized (or underlined if you don't have access to italics);
Species have two word names, the first part of which is the same as the genus name (e.g., Panthera leo, Homo sapiens, Ginkgo biloba, Tyrannosaurus rex)
The genus name is ALWAYS Capitalized, the second part ("trivial nomen" or "epithet") is ALWAYS in lower case, and the name is ALWAYSitalicized or underlined;
The complete trinomial for a species includes the full species name PLUS the family names of the original describers of the species, and the year of description.
Example: Tyrannosaurus rex Osborn 1905
If the species was originally in a different genus, but was lumped into a previously named species OR split into an entirely new genus, the author name and date of the original species is put in parentheses:
Example: Pithecanthropus erectus DuBois 1892 is typically lumped into the same genus as Homo sapiens Linnaeus 1758. So the first species would now be written out as: Homo erectus (DuBois 1892).
Species names can be abbreviated by using only the first letter of the genus name, followed by a period (NEVER by a hyphen): H. sapiens and T. rex are correct; H. Sapiens or T-Rex are WRONG!!;
Each genus has a type species: the one species uniquely associated with that genus;
All taxon names other than species have one word names, which are Capitalized; all taxon names other than genera and species are in roman letters (i.e., they are never italicized/underlined): Dinosauria, Tyrannosauridae, Animalia; not Dinosauria, tyrannosauridae, or animalia.
Because there is disagreement about the features used to define a particular species or genus, different biologists and paleontologists will sometimes disagree about which specimens belong in a particular species, and which species belong in a particular genus (and so forth).
Taxonomists who consider a particular set of specimens to represent many taxa are called splitters; those who consider a particular set to represent few taxa are called lumpers;
If a taxonomist feels that some specimens of a genus belong to an as-yet unnamed species, they can split these specimens off as a new species (which a new type specimen);
On the other hand, if a taxonomist considers that two previously named species are not distinct enough from each other to truly be distinct species (that is, the taxonomist regards the two names as synonyms), they may lump them together:
In these cases, the Rule of Priority is used: whichever of the names was published first (the senior synonym), even if only by days, is the name that must be used;
The junior synonym is then abandoned
The same case applies to genera: if two genera are thought to represent the same genus, the first named genus name is the one that is used.
For those interested in a website concerning some unusual Linnaean species names, click here.
Types and Type Specimens
Another aspect of Linnaean taxonomy is that each species must have a particular type specimen. This is a particular individual preserved specimen (extant animal) or fossil (extinct animal) that is the "name holder" for that species. A type specimen is specifically referred to in the original description and diagnosis of the species. It need not be the most complete specimen known at the time (although that helps, as the more complete it is, the better the chance a less-complete individual can be compared to it!). The type specimen plus all the additional (referred specimens) are collectively called the hypodigm. Ultimately, if a species is regarded as being "valid" (that is, representing a real species in Nature), the type specimen is the only individual that is absolutely certain to belong that that species.
Similarly, each genus has a particular type species. This is the particular species to which the genus name is linked. If a genus is valid, the type species is the only species that is absolutely guaranteed to be within that genus.
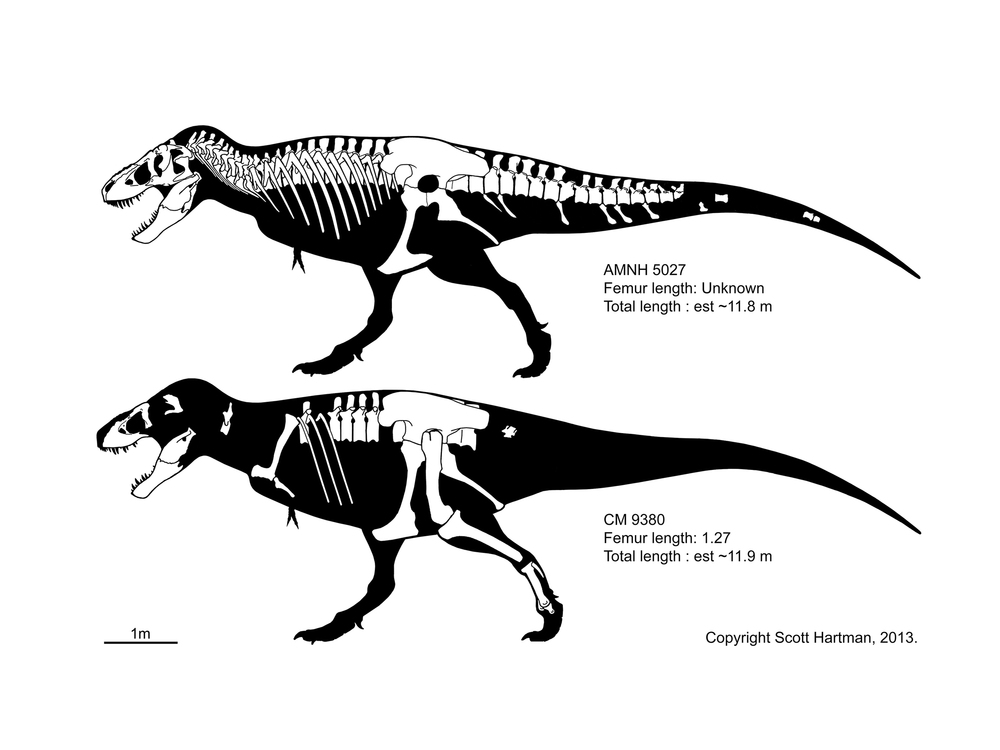
As an example, CM 9380 (in the collections of the Carnegie Museum of Natural History) is the type specimen of Tyrannosaurus rex, and Tyrannosaurus rex is the type species of the genus Tyrannosaurus
There are several types of type specimens:
Holotype: Single specimen designated as type bearer
The type of the type species of a genus is a genoholotype
Syntype: Several specimens designated as type bearers. Should be from same locality!
Paratype: An additional specimen other than a holotype used in the species description
Lectotype: One of the syntypes later chosen as a single holotype. Often done so if it is problematic whether all the syntypes belong to the same species
Neotype: Replacement for a lost or destroyed type
Plesiotype: Specimen used in redescribing an existing species. A plastotype is a variation on the neotype: it is a cast of the original type specimen that replaces a lost type
Objective and Subjective Synonyms
If two synonyms are based on the same type specimen, they are said to be objective synonyms. If they are based on separate type specimens, they are subjective synonyms.
Problematic Names
Obsolete Names: as of 1 January 2000, if a name has not been used as a valid name for over 100 years, it is considered obsolete
and is not considered to have priority over a more recently-coined name
Homonyms: Two names referring to different taxa but with same spelling
E.g., Microtia Bates 1864 (a modern butterfly) and Microtia Freudenthal 1976 (a fossil rodent)
The name is said to be occupied, requiring a name change for the junior honomym
The senior homonym retains its name; the junior one uses the next most senior synonym (if any) or needs a new name
Invalid Names: Names for which the type specimen is found to not have diagnostic features when more relevant specimens are
recovered
E.g., Deinodon horridus Leidy 1856, teeth of carnivorous dinosaur from Judith River Fm of Montana. Later discoveries show two
distinct skeletal taxa (Gorgosaurus libratus and Daspletosaurus torosus) with "Deinodon" teeth. Since we can't
determine which of the two the type of Deinodon belongs to, it is considered invalid
When You're Not Sure of the Species Identification:
Genus sp.: You recognize that it belongs to a particular genus, but you don’t know which species
Do not confuse with the format Genus sp. 1, sp. 2, etc.: that implies you recognize new species but haven't gotten around to
naming them yet
Genus cf. G. trivial: the "cf." stands for confer, compare. Probably the same as, but not yet demonstrated to
belong to, the known species G. trivial
Genus aff. G. trivial: the "aff." stands for affinis, has affinities with. Similar to, but does not belong to, the
known species G. trivial
Difficulties of Paleo-taxonomy
Combining this lecture with the last, there are some inherent difficulties with the taxonomy of fossils that are not (generally) problems with living species:
Incompleteness: No fossil is fresh! Even a frozen Pleistocene mammal has degraded somewhat, and the fast majority of fossil taxa are known only from hard parts or impressions. (And in the case of many taxa, those skeletons are far from complete.) So it becomes very difficult to assess the shared identity (or lack thereof) of two specimens from different parts of the body, unless there are other specimens which link the two. And, of course, we lack the evidence for the most part of features like soft tissue, color, behavior, etc., that are used to distinguish closely-related species today
Conspecifics vs. Sympatric species: Two individuals that are members of the same species are said to be conspecific. (The comparable word for genera is congeneric). When we find two related specimens we might consider them members of the same species. But there is another possibility: they are from different species who simply inhabited the same place and time (sympatric). Differences between conspecific individuals (as we saw before) can be from numerous sources:
Individual variation
Sexual dimorphism
Taphonomic differences
Ecomorphic differences: even individuals with essentially the same genome will have different phenotypic expression depending on the environment in which they grow up
Ontogenetic differences: while taxa with continuous or modular growth (many mollusks, brachiopods, forams, etc.) still have their earlier morphologies present in their later forms, those that undergo ecdysis (arthropods & kin) can have radically different forms at each growth stage (instar). Without directly seeing the living animal metamorphosis, we have to infer what series of instars are from which species. Similarly, although many vertebrates lack true metamorphosis, different life stages can have significantly different body proportions, tooth types, muscle scar development, and more. These different ontogimorphs might be mistaken as separate taxa rather than growth series. (Collectively, instars and ontogimorphs and other distinct expressions of morphology at different life stages are called semaphorants.)
Parataxonomies: alternative taxonomic systems parallel to the primary one. Ootaxonomy (taxonomic system for vertebrate egg
fossils) is one such set. Ichnotaxonomy SHOULD be one, but is considered under the zoological code.
The Peculiarities of Paleobotanical Taxonomy
Different plant organs often have very different taphonomic potentials and so are rarely preserved together. As a consequence, each
paleoplant organ system (leaves; trunks; roots; stems; pollen/spores; etc.) has its own taxonomic system. WITHIN each system, the principles
of priority apply. But discovery that (for example) leaf A, seed B, and trunk C belonged to the same paleoplant does NOT result in synonomy
of A, B & C. This is in gross violation of ANY species concept/criterion, and of the neontological botanical code, but it is ultimately very pragmatic.
Pronouncing Taxon Names: How should one pronounce a taxon name? Short answer first: it really doesn't matter too much, so long as people know what taxon you are referring to. One reasonable approach is to use the pronunciation preferred by the person who coined the name (if you can find out what that pronunciation is) or at least preferred by the specialists in that taxon (if there is a consensus; sometimes there is, sometimes there isn't).
If you want something more rigorous, one preference is to use late northern Continental Latin pronunciation. This is the language of Kepler, Copernicus, and (most importantly) Linnaeus. Alternatively, you might consult this site for suggestions. (There are some differences between these: in the former "-idae" the vowel at the end is like the "a" in "plate"; in the latter it is like the "e" in "we".) In any case, please do NOT use either Church Latin or Classical Latin: those pronunciations represent language forms many centuries before the rise of scientific Latin.




{kind=link}