


Key Points:
•Ecology is the study of the factors which control the abundance and distribution of organisms. It examines the flow of energy in living systems, and their interactions with the environment.
•A primary aspect of ecology is trophic interactions: who eats whom. Energy enters ecosystems from sunlight captured by photosynthesis; herbivores eat the plants; carnivores eat herbivores (and other carnivores); and decomposers break down dead tissue into its component parts.
•Since organisms respond to changes in temperature, humidity, etc., their fossils are excellent proxies for ancient environments.
•The isotopic composition of fossil hardparts can be used to examine paleoenvironment, paleoecology, physiology, and more.
"Naturalists try to arrange the species, genera, and families in each class, on what is called the Natural System. But what is meant by this system? Some authors look at it merely as a scheme for arranging together those living objects which are most alike, and for separating those which are most unlike; or as an artificial means for enunciating, as briefly as possible, general propositions, that is, by one sentence to give the characters common, for instance, to all mammals, by another those common to all carnivora, by another those common to the dog-genus, and then by adding a single sentence, a full description is given of each kind of dog. The ingenuity and utility of this system are indisputable. But many naturalists think that something more is meant by the Natural System; they believe that it reveals the plan of the Creator; but unless it be specified whether order in time or space, or what else is meant by the plan of the Creator, it seems to me that nothing is thus added to our knowledge. Such expressions as that famous one of Linnaeus, and which we often meet with in a more or less concealed form, that the characters do not make the genus, but that the genus gives the characters, seem to imply that something more is included in our classification, than mere resemblance. I believe that something more is included; and that propinquity of descent, the only known cause of the similarity of organic beings, is the bond, hidden as it is by various degrees of modification, which is partially revealed to us by our classifications." -- Chapter 13, On the Origin of Species by Means of Natural Selection (1859), Charles Darwin
"The affinities of all the beings of the same class have sometimes been represented by a great tree. I believe this simile largely speaks the truth. The green and budding twigs may represent existing species; and those produced during each former year may represent the long succession of extinct species. At each period of growth all the growing twigs have tried to branch out on all sides, and to overtop and kill the surrounding twigs and branches, in the same manner as species and groups of species have tried to overmaster other species in the great battle for life. The limbs divided into great branches, and these into lesser and lesser branches, were themselves once, when the tree was small, budding twigs; and this connexion of the former and present buds by ramifying branches may well represent the classification of all extinct and living species in groups subordinate to groups. Of the many twigs which flourished when the tree was a mere bush, only two or three, now grown into great branches, yet survive and bear all the other branches; so with the species which lived during long-past geological periods, very few now have living and modified descendants. From the first growth of the tree, many a limb and branch has decayed and dropped off; and these lost branches of various sizes may represent those whole orders, families, and genera which have now no living representatives, and which are known to us only from having been found in a fossil state. As we here and there see a thin straggling branch springing from a fork low down in a tree, and which by some chance has been favoured and is still alive on its summit, so we occasionally see an animal like the Ornithorhynchus or Lepidosiren, which in some small degree connects by its affinities two large branches of life, and which has apparently been saved from fatal competition by having inhabited a protected station. As buds give rise by growth to fresh buds, and these, if vigorous, branch out and overtop on all sides many a feebler branch, so by generation I believe it has been with the great Tree of Life, which fills with its dead and broken branches the crust of the earth, and covers the surface with its ever branching and beautiful ramifications." -- Chapter 4, On the Origin of Species by Means of Natural Selection (1859), Charles Darwin
Ecology
First up, some definitions:
How does energy go from the non-living environment into the living world? One major way is photosynthesis. Here is the process greatly simplified:
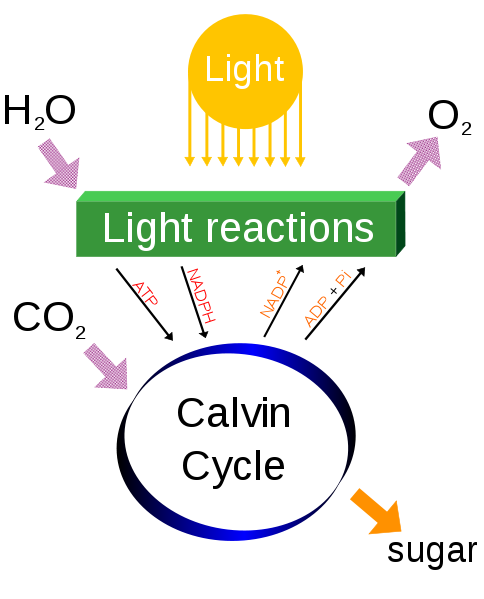
And here is the photosynthesis equation, simplified:
6CO2 + 6H2O + Sunlight yields C6H12O6 + 6O2
Or, in other words, photosynthesizers (plants and algae) take in water and carbon dioxide and using sunlight convert these into sugars and waste oxygen gas. Photosynthesizers are the major form of autotroph: organisms that don't eat other organisms.
In contrast, most other life forms (animals, fungi, most "protists", etc.) are heterotrophs: we get nutrients from digesting other life forms, primarily via the aerobic respiration equation:
C6H12O6 + 6O2 yields 6CO2 + 6H2O + Energy
(That is, food (glucose) plus oxygen yields waste carbon dioxide and waste water, plus energy).
In this reaction, we take in sugars and oxygen and release waste water and carbon dioxide.
So plants, algae, and other photosynthesizes are basically made up of air (and a little bit of other matter drawn from soil and/or water), while animals and other heterotrophs are made of food (ultimately plants) and air. This most important aspect of ecology is thus trophic relationships: who eats whom. These mark out relationships in a food chain of organisms that eat the organisms below them. However, since in nearly every case any species eats more than one type of food, trophic relationships are better displayed as a food web, comprised of many interlocking food chains.
The trophic level is the position an organism occupies of a food chain. Producers do not consume other animals: in terrestrial ecosystems, these are plants (which "eat" sunlight). Decomposers break down the bodies of any kind of organisms. Consumers are those that eat other organisms: 1st order consumers are herbivores (plant-eaters); 2nd order consumers eat 1st order consumers; 3rd order consumers eat 2nd and 1st order consumers; and so on, up to the apex predator (consumers that eat lower order consumers but are not typically eaten by any species but themselves).
As you move up a food chain, there is less and less energy available. Plants receive much (but by no means all) of the energy of incoming sunlight; some of that energy becomes stored as their tissues, but the rest of it is used for them to run their bodies. Herbivores consume plants, and gain some (but not all) of that stored energy, some of which they used to build their own tissues but the majority being used for their own operation (running, breathing, pumping blood, etc., etc., etc.). Predators eat the herbivores, gaining some (but not all) the stored energy in the tissues of the herbivores, using some to build their own bodies, but the rest for their operations. Thus, the amount of energy available at each trophic level becomes less and less: this is described as the energy pyramid.
Investigation of individuals, populations, and communities of ancient organisms and their interactions with and dynamic responses to changing environments. On the one hand, the data is hampered by the various taphonomic filters and our inability to directly observe interactions. On the other, the temporal scope of paleoecology greatly exceeds "neo-ecology" and consequently may be better able to capture the "big picture".
Niche Theory
As defined by British ecologist C.S. Elton, a species' niche is "its place in the biotic environment, its relations to food and enemies". This zoo-centric concept was expanded by G. Evelyn Hutchinson to represent an n-dimensional hypervolume where the dimensions are environmental conditions and resources that define the requirements of the species to practice its way of life (i.e., to persist and propagate).
We can now recognize several aspects of niche theory:
Niches are subject to change. Some niche variables (such as bedrock) don't move in ecological time scales (but of course do move on geologic time scales!) Others (e.g., distribution of sunlight over the year) move following various cycles. Still others (e.g., climate; presence/absence of other taxa) may move following far more complex patterns. As environmental variables shift in real space, organisms may move with them (the actual individuals for motile species; the propagules for plants and sessile animals), so long as the new region is within the potential niche and dispersion is possible from one space to another. (If these conditions are not met, it is likely that the population goes extinct!)
Communities: Are They Real>
A long, on-going debate in ecology is whether the communities we see in nature are real entities as such:
Paleontology's contribution is to see if there are patterns of assemblages in the past that share the same taxa (or their immediate ancestors) as the present, or if the associations change over time; to examine if there were communities that were widespread in time and space in the past that are no longer extant; and to examine the long-term trends in ecology (macroecology).
One major test is for autochthony. Are the fossils in life position? Articulated? Are there low levels of breakage and abrasion? Are they poorly sorted in terms of size? Are there low levels of shared orientations (and thus little sign of current, and thus possible transport)? If so, than the animals were almost certainly living next to each other prior to burial.
(It is worth noting that some allochthony can be good!! For instance, we would like to know the nekton and plankton in a region, since they do interact with the benthos. And since most terrestrial sedimentary deposits are aquatic (lacustrine or fluvial), we would like to have some transport of the land animals and plants into these water bodies to know they were in the region.)
The best test is to find multiple stratigraphic and/or geographic occurrences of the same assemblage: more commonality indicates a better chance it is not just a random taphonomic assemblage.
Paleoecology shows that some communities do persist for long periods of time, both extant styles or extinct ones (like the mammoth steppe of the Pleistocene Arctic). On the flip side, there are non-analogue biomes: combinations of taxa (some extant) that are found together in the past which are never found together today.
Assessing Trophic Relationships
One of the most important aspects of ecology (and paleoecology) is determining who ate whom: that is, reconstructing the food web. Sometimes there is direct trace fossil evidence: for example, this humerus of the Late Cretaceous Asian duckbilled dinosaur Saurolophus: the arrows point to bite marks generated by a tyrannosaurid (almost certainly Tarbosaurus, which is known from the same fauna).
However, sometimes we do not have such direct evidence. In these cases, we can make relatively reasonable assessments. For example, it is unlikely that small carnivores habitually killed the largest herbivores in their environment (while the latter were adults, at least: babies wouldbe fair game), and it is unlikely that there were species that were not eaten by SOMETHING.
In the case of the herbivores, however, it becomes somewhat more difficult. For one thing, it is often the case that the conditions that are best to preserve plant fossils (especially of leaves and fruits) are not good for preserving bone, and vice versa. So we often lack good knowledge of the flora that co-existed with any fauna. That said, we have the digested plant remains in coprolites as positive evidence that herbivores ate particular plant species (although referring a coprolite to a particular herbivore species can be difficult.)
Fossils represent one of our major clues to restoring paleoclimate. One primary technique is the modern analog approach. Some aspect of the community (species composition, for instance) is compared from a modern environment (whose parameters are known) to a fossil one.
One classic example is leaf margin analysis. It is observed in modern floras that proportion of entire (that is, smooth) leaves in a flora has a rather strikingly accurate correlation with mean annual temperature. So looking at fossil leaf populations, the ratio of entire vs. jagged leaves in the fossil horizon is used to predict what the ancient mean annual temperature was at the time of deposition.
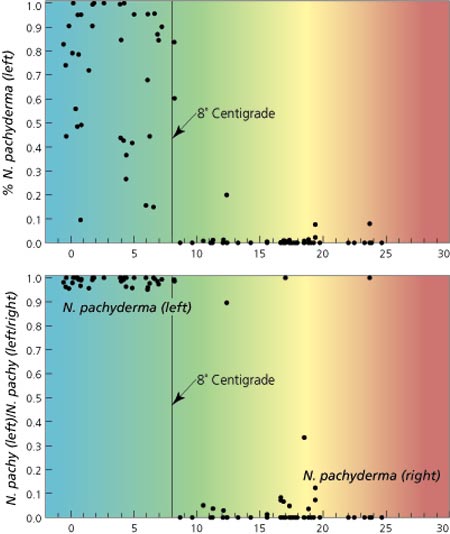
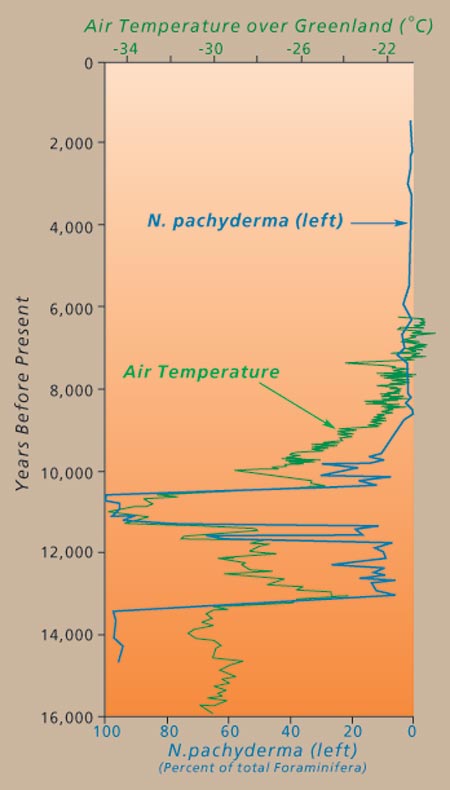
Similarly, the distribution of some species (for instance, some foraminiferans) are proxies of ancient water temperature. Looking at the the changing distribution of fossils of these species through time and space allows paleoclimatologists to track changes in ancient temperatures. One particular paleothermometer is Neogloboquadrina pachyderma, whose direction of coiling changes at 8°C. Tracking changes in coiling direction down stratigraphic section allows the recognition of when the local temperature passed this treshold.
The hard parts of organisms retain stable isotopic ratios of the time of formation so long as diagenesis does not otherwise alter these. Some of the more commonly utilized isotopic elemental systems in paleontology are C, O, N, P.
These have been used to reconstruct:
To Syllabus.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}