


Key Points:
•Understanding the locomotion and other activity of dinosaurs is improved by studying biomechanics: reconstructing their actions using the principles of engineering and physics.
•Ichnology is the study of trace fossils.
•Ichnotaxa are named trace fossils: they technically are names for the objects themselves, not for the biological species which made them.
•The most commonly studied dinosaur trace fossils are footprints. Although it is often difficult to assign an ichnofossil to a particular biological species, we can often tell the larger clade to which it belonged.
•Trackways allow us to estimate the speed at which a dinosaur was moving.
•Land animals range from more graviportal ones (adapted to weight support) toward cursorial ones (adapted to running). Some adaptations of limb and girdle features can be used to recognize trends to one or the other of these behaviors.
•As animals grow, their proportions do not necessarily remain the same. More often, there is some form of allometry: differential growth in proportion.
DINOSAURIAN TERRESTRIAL LOCOMOTION
How can we figure out how dinosaurs walked, ran, fed, etc?
Fossils are our primary line of approach, both body fossils and trace fossils.
In the case of body fossils, a few traits (skeletal ones) can be read directly from fossils. Others (muscles, tendons, sense organs, and so forth) might have osteological correlates (that is, direct traces on the bone, even if they are not made of bone themselves.) Still others might be determined by biomechanical or other functional studies.
There are four main methods of inference in dinosaur biology:
Let's consider the issue of dinosaur locomotion. Dinosaurs are essentially striders: their hindlimbs were restricted to antero-posterior movement.
Ichnology: the study of trace fossils.
Most dinosaur ichnologists concentrate on footprints and trackway analysis.
An individual footprint represents:
Footprints give direct information about the soft tissues of the bottom of the foot, and about the natural position of the toes.
Trackways, however, give even more data. By measuring the stride length, and estimating hip height, the speed of the dinosaur at the time of that trackway can be calculated. These data tend to show dinosaurs walking around at speeds comparable to modern large-bodied mammals.
However, trackways do have some problems:
Footprints and trackways can, however, reveal the presence of dinosaurs not yet known by body fossils (such as Middle Jurassic North American dinosaurs).
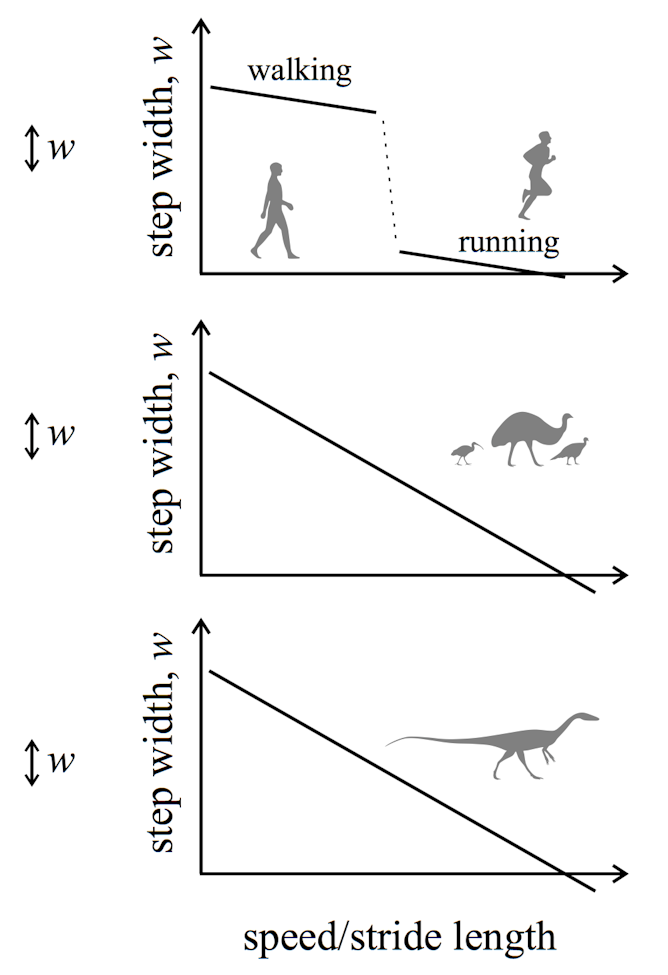
An interesting new discovery in ichnology is that Mesozoic dinosaurs (like extant birds) did not have discrete gaits like walk-jog-run or walk-trot-canter-gallop. Instead, they had a continuous transition of speed, stride length, and step width.
There is a whole discipline of ichnotaxonomy: the naming of trace fossils. However, it must be remembered that these are sedimentological entities, not biological entities: the same animal can produce tracks given entirely different ichnotaxonomic names if it is walking slowly or running; on soft mud or hard mud; if adult or juvenile; etc.
One interesting note: almost no dinosaur trace fossil shows tail drag marks: this was some of the first evidence that dinosaurs held their tails up above the ground.
Trace fossils can help us understand something about group behavior. A recent study of Alaksan hadrosaurs showed many individuals moving in the same direction at the same time (based on the similar quality of footprint preservation.) Additionally, by counting up the trackways of different sizes, it helped to give an estimate of the relative fraction of the herd of different growth stages.
Deciding whether an animal is running or not is a trickier problem that it first sounds. The classic definition of running is "a gait in which there is a suspended phase" (that is, all feet of momentarily off the ground). We can generally approximate when a trackway has reached that point. However, the modern (or "kinetic") definition of running is "a gait in which the leg is at its most compressed at the middle of its contact with the ground" (like the spring on a pogo stick).
We can apply principles of functional morphology to understanding how dinosaurs operated: that is, work out the function of organisms by the shapes and structures of their body parts.
For instance, we recognize from their parasagittal limbs and digitigrade feet that dinosaurs were striders: they walked one foot directly in front of the other rather than with splayed limbs. Study of modern striders (mammals and birds) show a continuum from cursorial (running specialists) to graviportal (support specialist) forms. Cursorial animals tend to have:
while graviportal animals tend to have:
Examining these traits in dinosaurs, we find that some small ornithopods but especially many coelurosaurian theropods show cursorial adaptations relative to other dinosaurs, while stegosaur and ankylosaur thyreophorans and eusauropod sauropodomorphs show greater graviportal adaptations.
Of course, there are also issues of scaling: as body size increases, the mass supported by any surface (like the bottom of the foot) or cross-section (like the cross-section of the femur) increases faster, so the relative mechanical strength of the animal decreases. Body parts do not all grow at the same rate: that is, they often show allometry:
To Next Lecture.
To Previous Lecture.
To Lecture Notes.

{kind=link}