

Key Points:
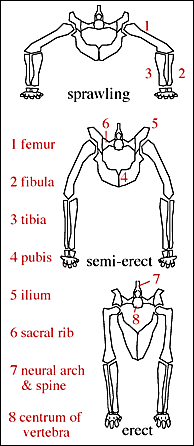
•There is a shift towards obligate bipedalism, parasagittal (upright) posture, digitigrade stance, a grasping manus, and a perforate (open) acetabulum on the lineage up to Dinosauria.
•Dinosauria is comprised of three major clades: Ornithischia, Sauropodomorpha, and Theropoda. Traditionally, sauropodomorphs and theropods were recognized to form a clade Saurischia. However, recent discoveries have reduced the support for this hypothesis, and alternative relationships are possible.
•Dinosauria first appears by early in the Late Triassic, and may have actually originated even earlier. Many (although not all) of the oldest dinosaurs are from the southern parts of Pangaea, but this may be due to a sampling bias.
•There are several Triassic (Herrerasauria; Eodromaeus; Tawa + Chindesaurus; etc.) and even Jurassic (Chilesaurus) dinosaurs whose position within the clade are presently very uncertain.
I. Origin of the Dinosaurs
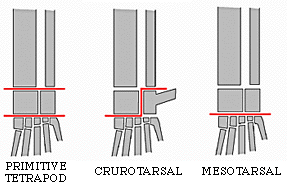
Modern birds and all archosaurs closer to them than to crocodilians are the Avemetatarsalia (bird metatarsals). Just recently the most basal branch of avemetatarsalians was discovered: the Aphanosauria ("hidden reptiles"). Only recognized in 2017, these were medium-sized (2-3 m long or so) quadrupedal, long-necked Middle Triassic archosaurs. Best known of these is Teleocrater of Tanzania.
The remaining avemetatarsalians belong to the Ornithoidira ("bird necks"). These were generally small-bodied animals that evolved under the shadow of their pseudosuchian cousins, differing from typical diapsids by having elongate tibiae and metatarsi (suggesting that they were even faster than typical sauropsids.) Their name is derived from the fact that their cervical vertebrae were greatly modified so that they look very different from dorsal vertebrae, but instead could allow the neck to form an S-shaped curve.
One branch of the ornithodirans that appeared in the Late Triassic are the Pterosauria: the first group of powered fliers among the vertebrates. We'll look more at the pterosaurs toward the end of the course.
The other major branch of ornithodirans are the dinosauromorphs. The oldest dinosauromorphs are known from the Middle Triassic in terms of body fossils, but footprints show that some very small dinosauromorphs were present as early as the Early Triassic. Dinosauromorphs modified the hindlimbs further than the ancestral ornithodirans to evolve:
The combination of these traits allowed the little dinosauromorphs to run and accelerate in a bipedal mode all the time, not just at top speeds like typical diapsids. The parasagittal gait and hinge-like ankle also allowed dinosauromorphs the ability to move more actively and constantly rather than only in short bursts of speeds; thus, they were striders. Additionally, although early dinosauromorphs were small (~30 cm long for Marasuchus, 1-3 m long for "silesaurs" and basal dinosaurs, etc.), the presence of limbs directly underneath the body meant that this lineage to grow to much larger size than any previous clade while still remaining terrestrial and mobile (sprawlers relegated to a semi-aquatic life if they became too big to support their weight).
Early dinosauromorph lineages were typically a meter or less long. These include the lagerpetids, such as Ixalerpeton, Dromomeron, and Lagerpeton; and the lagosuchids, such as Marasuchus and (possibly) Saltopus.
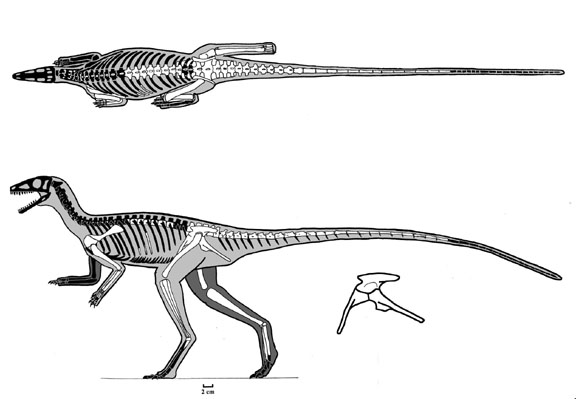
The largest dinosauromorphs other than dinosaurs are the Silesauridae. These were quadrupedal animals typically about 1-2 m long (although fragmentary remains of one at least 3 m long has been found in Tanzania). They and dinosaurs share the specialization of elongate pubes and ischia (which may have aided in increased respiratory ability: more air per breath). Primitive forms look to have been carnivorous, but most silesaurids were herbivores. Silesaurids had been known from teeth and bones of the Middle and Late Triassic from around the world, but had been thought to be from primitive dinosauromorphs, ornithischians, sauropodomorphs, and/or theropods until the more complete specimens of Asilisaurus and Silesaurus itself let people know what this group looked like. At least some have a toothless portion of the front end of the dentary, likely covered by a rhamphotheca (beak). (As will be discussed later on, an alternative hypothesis is that silesaurids ARE dinosaurs!)
Also occurring in the Middle Triassic (of Tanzania, in this case), is Nyasasaurus. Incompletely known, what data is available shows that it is either a dinosauromorph closer to Dinosauria than are silesaurids, or (possibly) the oldest known dinosaur. (At least one study provisionally put it quite far up the dinosaur tree, as a massospondylid "core prosauropod". If true, this would mean a tremendous part of the diversification of Dinosauria had occurred by the Middle Triassic that is not yet documented in the fossil record.)
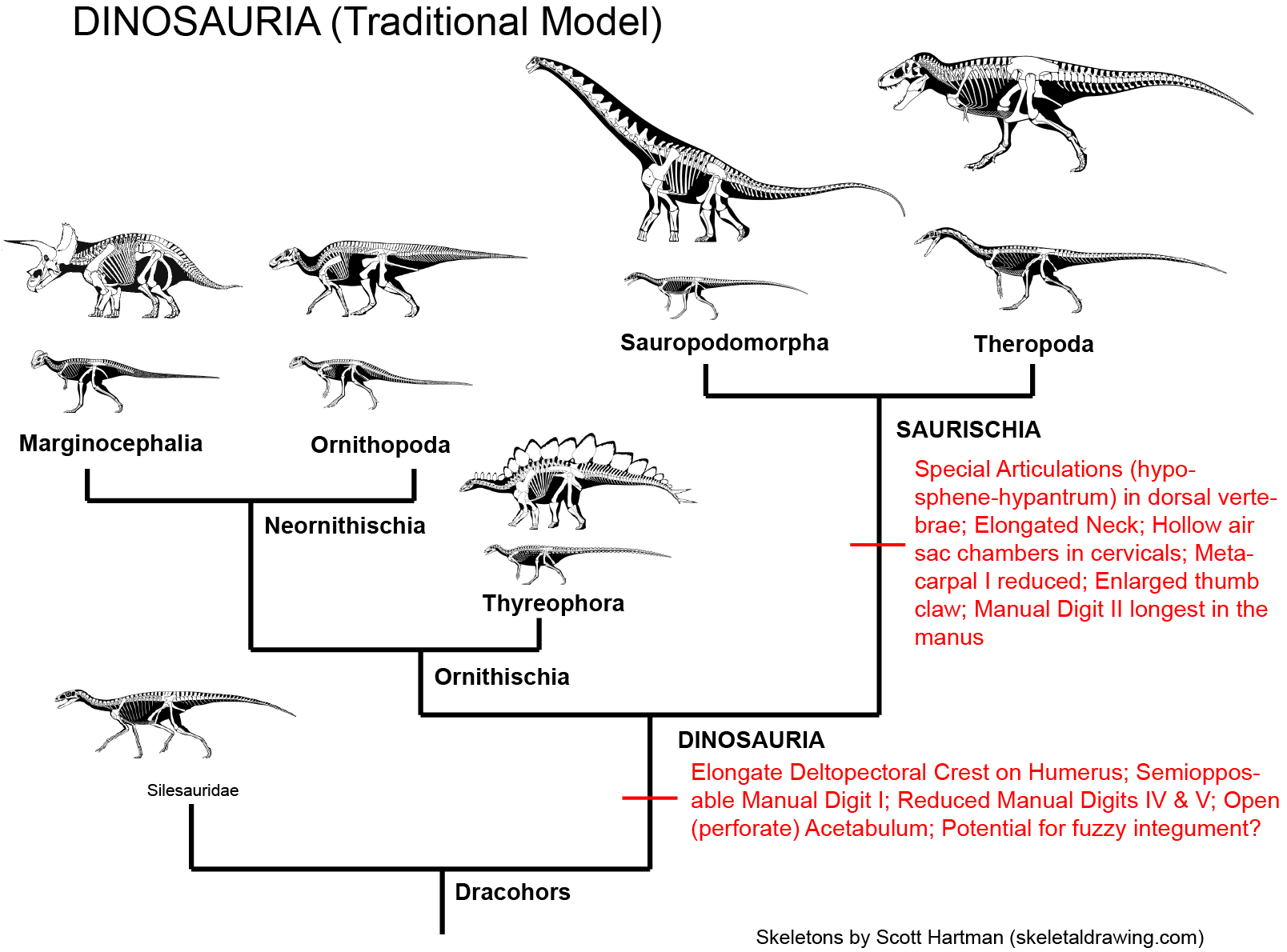
Silesaurids and Nyasasaurus are the closest relatives to Dinosauria, but they do not seem to be true dinosaurs: that is, they lack the shared derived features expected in the concestor of Iguanodon, Diplodocus, and Megalosaurus and its descendants. (But see below.) Those features (the shared derived features of Dinosauria) are:
A possible shared derived feature of Dinosauria is the presence of at least some fuzz rather than scales. This rather startling concept is because basal members of one lineage and more derived members of the other show the presence of such structures. While many dinosaurs are preserved with impressions of scales, there are several with other forms of integument. These include:
At present we have no definite evidence of fuzz or other non-scales in sauropodomorphs. But it is possible that a dinosaurian evolutionary novelty is the ability to generate at least some quill, tuft, or branching integument rather than just scales. However, different clades of dinosaurs express this ability in different fashion.
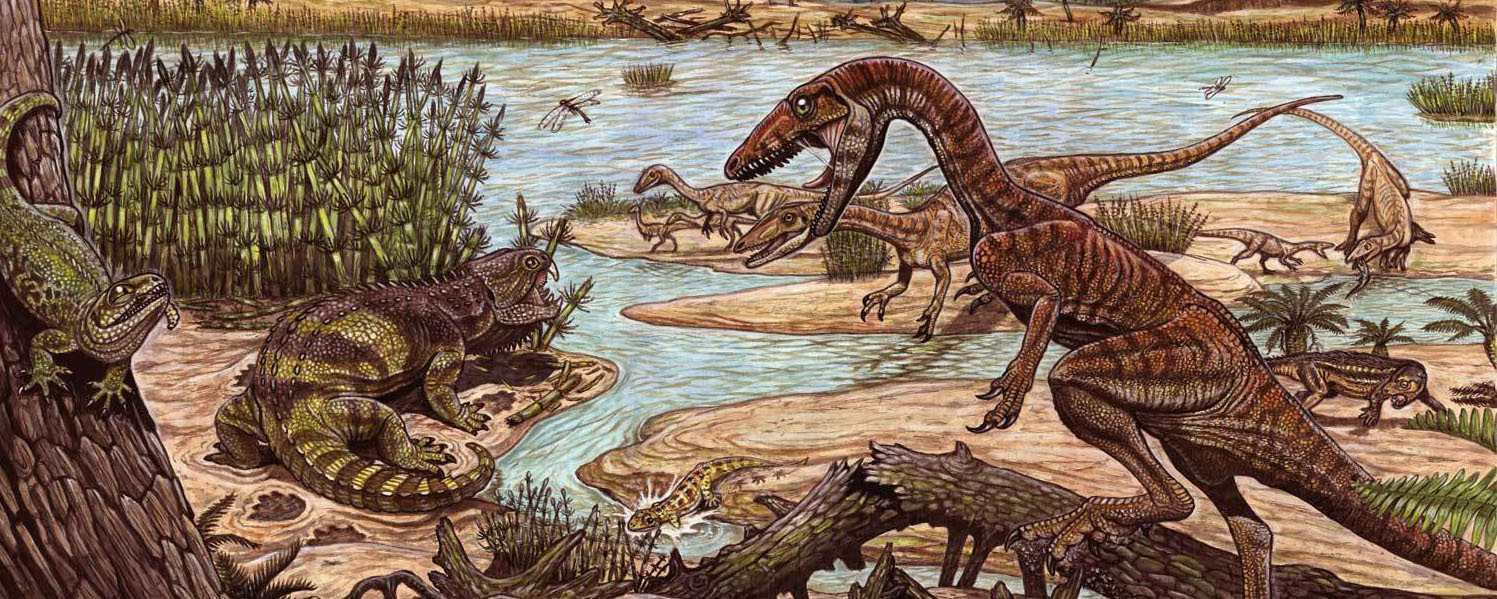
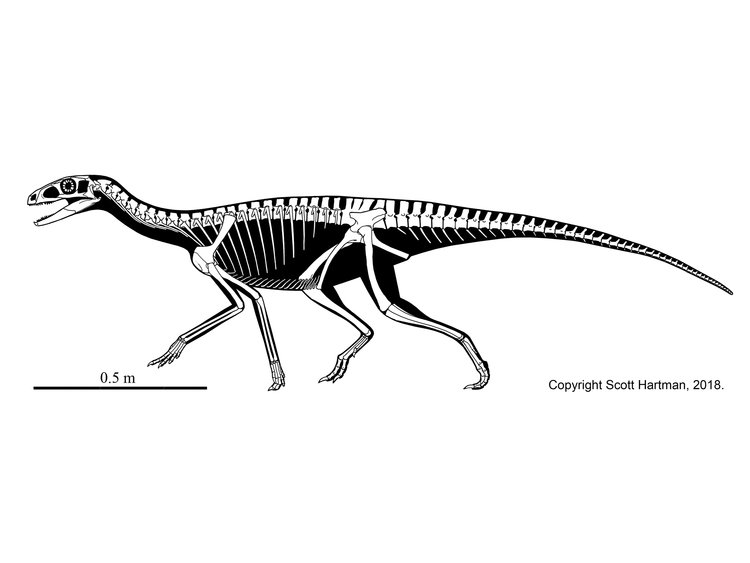
Early dinosaurs were all around 1-2 m long bipeds with grasping hands. Footprints that might come from dinosaurs are found in the Middle Triassic of Argentina, but these may be from early members of the dinosaur lineage that evolved before the origin of Dinosauria proper.
The oldest known definite dinosaurs are from about 230 Ma, at the very beginning of the Late Triassic Epoch. The very oldest dinosaurs are from Argentina (the Ischigualasto Formation) and Brazil (the Santa Maria Formation), and from India; fossils from the same age from Europe, North America, and elsewhere have turned out to be non-dinosaurian.
Dinosauria contains three major clades:
Until recently essentially all dinosaur researchers considered the most basal split in Dinosauria to be between Ornithischia and a clade named Saurischia ("lizard hips"), the latter defined as Megalosaurus, Diplodocus, and all taxa closer to these taxa than to Iguanodon. However, recent studies call into question that relationship: we'll go into that in a little more detail shortly.
Well look at the shared derived characters of Sauropodomorpha and Theropoda in later lectures. But let's look a bit at the base of Ornithischia:
Simplified cladogram of Ornithischia:
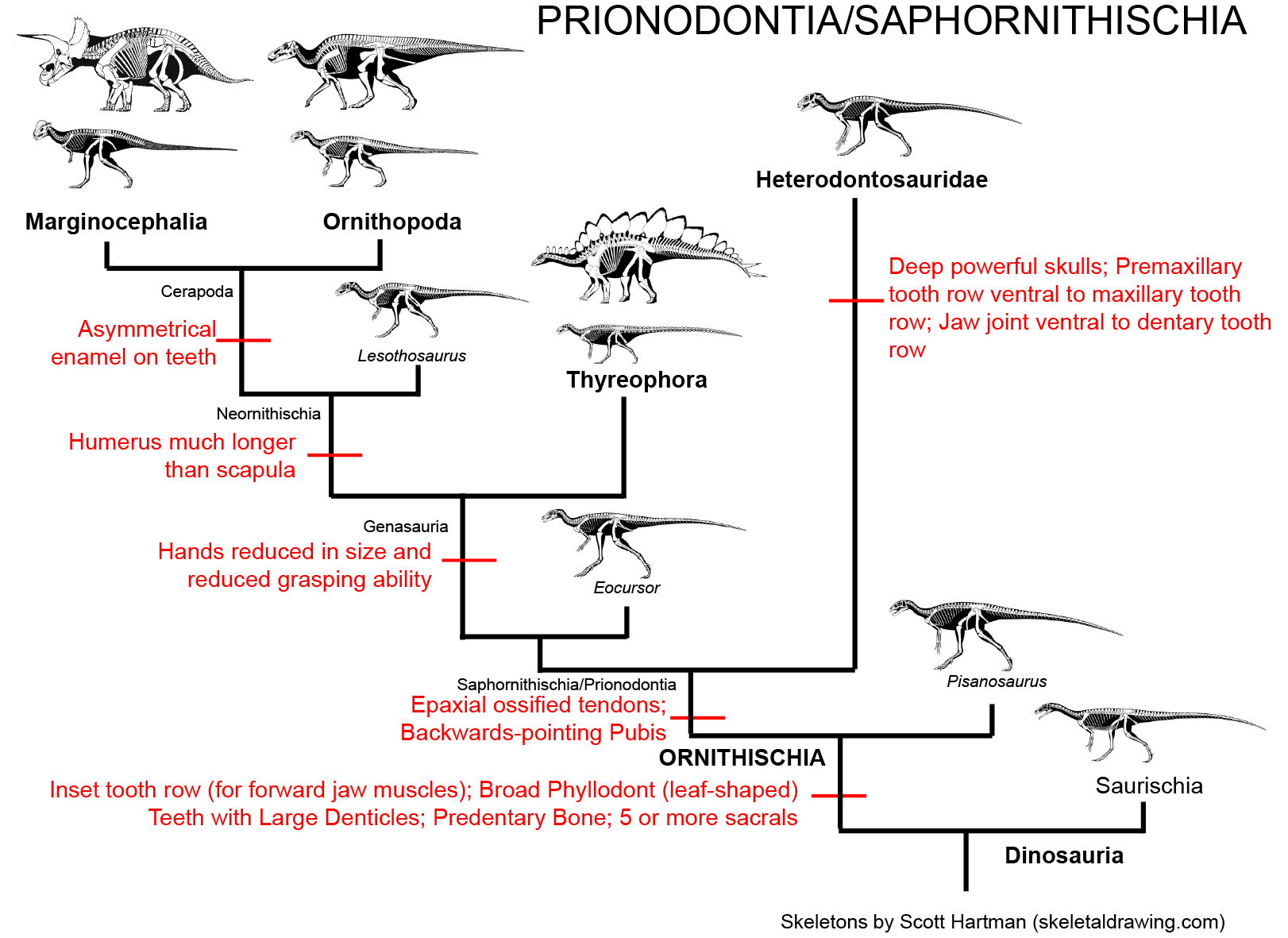
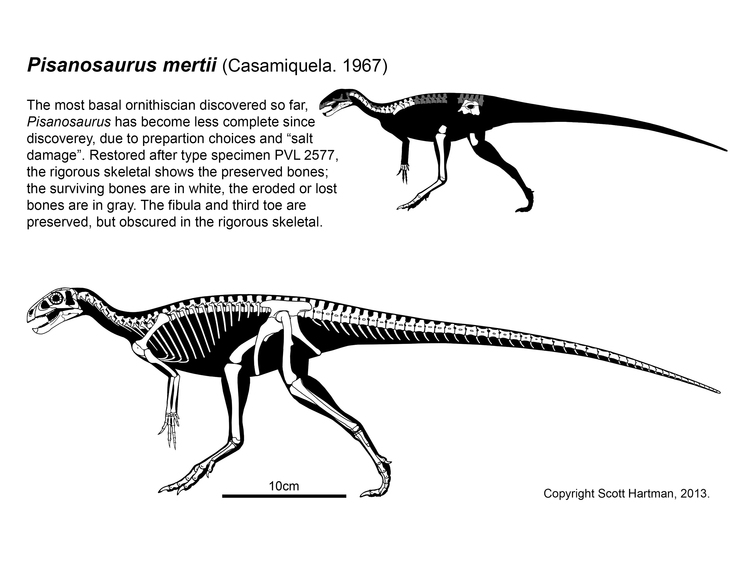
Ornithischians are, at present, not definitely present in the Triassic. Since the 1970s the oldest and most primitive ornithischian was thought to be Pisanosaurus of the early Late Triassic Argentine Ischigualasto Formation. The fossil is incomplete, so many aspects of its anatomy are uncertain. But some new analyses in 2016 and 2017 have shown this is probably a silesaurid, not a dinosaur (although others show it is is indeed one.) (As we will see, there is an hypothesis where it is BOTH a silesaurid and an ornithischian!)
Ornithischia is characterized by the following traits:
Based on their tooth form and the retroverted pubis, ornithischians were herbivorous. (That doesn't mean that they were exclusively plant eaters, of course! In the modern world, many "herbivorous" sauropsids and mammals eat some meat.)
Major Alternative Dinosauria Arrangement I: Silesaurids are the Triassic Ornithischians: There are a number of derived features of the teeth and jaws shared by Ornithischia and Silesauridae. The traditional interpretation is that these are convergences, since silesaurids like the shared derived characters of Dinosauria. That said, a few analyses actually place Silesauridae as a basal branch in Ornithischia. The latest version finds that silesaurids are a paraphyletic group relative to the "classic" Ornithischia, with Pisanosaurus as the base of the latter. An advantage of this hypothesis is that it explains where the Triassic ornithischians are. However, if true then the features regarded above as shared derived dinosaurian traits (such as the modified hand and the perforated acetabulum) either evolved independently in derived ornithischians and saurischians, or were lost in Silesauridae. We'll see how this proceeds.
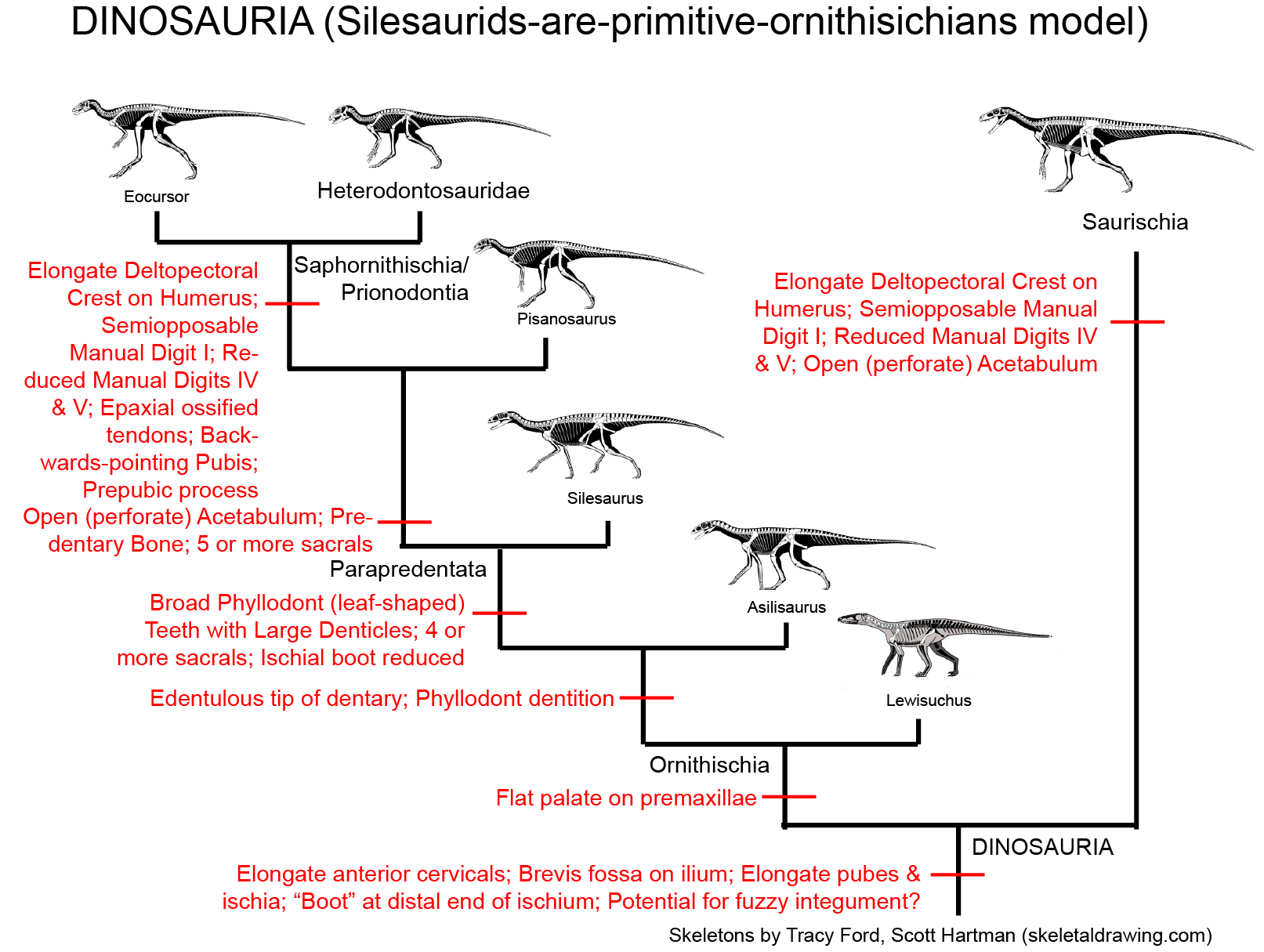
Simplified cladogram of Saurischia:
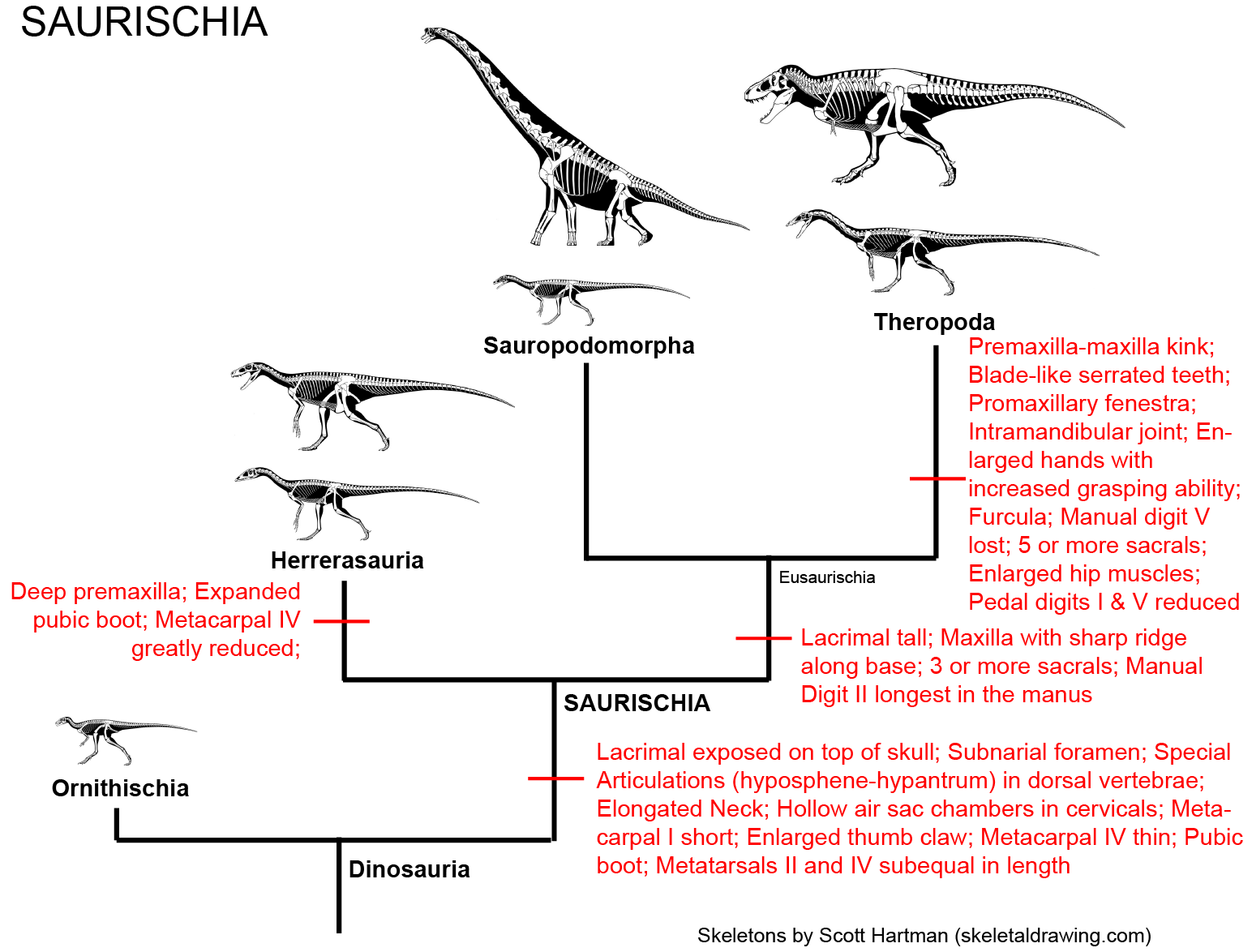
Saurischians are named after a primitive trait ("lizard hips" = "forwards-pointing pubis"; the ancestral condition), but they are in fact united by some shared derived traits. These include modifications of the snout and of the vertebrae: however, these are rather technical features and we're not go into them in detail in this course. Additionally, some of the traits long thought to be basal saurischian traits are now popping up in early ornithischians and in silesaurids, and so are not actually saurischian shared derived characters.
The saurischians were more common in the Late Triassic than their ornithischian sisters. The theropods include the long-necked herbivorous Sauropodomorpha and the (ancestrally carnivorous) Theropoda. Among other traits shared among the saurischians are:
(A special note: Owen's Dinosauria traits re-examined: If we look at some of the traits that Owen used to recognize Dinosauria, we now find that some were actually inherited from pre-dinosaurian ancestors (e.g., parasagittal stance, from the early dinosauromorphs) and others evolved convergently between Iguanodon, Hylaeosaurus, and Megalosaurus (giant size; more than two sacrals). Still, even though the traits to recognize them have changed, we still use the name "Dinosauria" to unite these reptiles.)
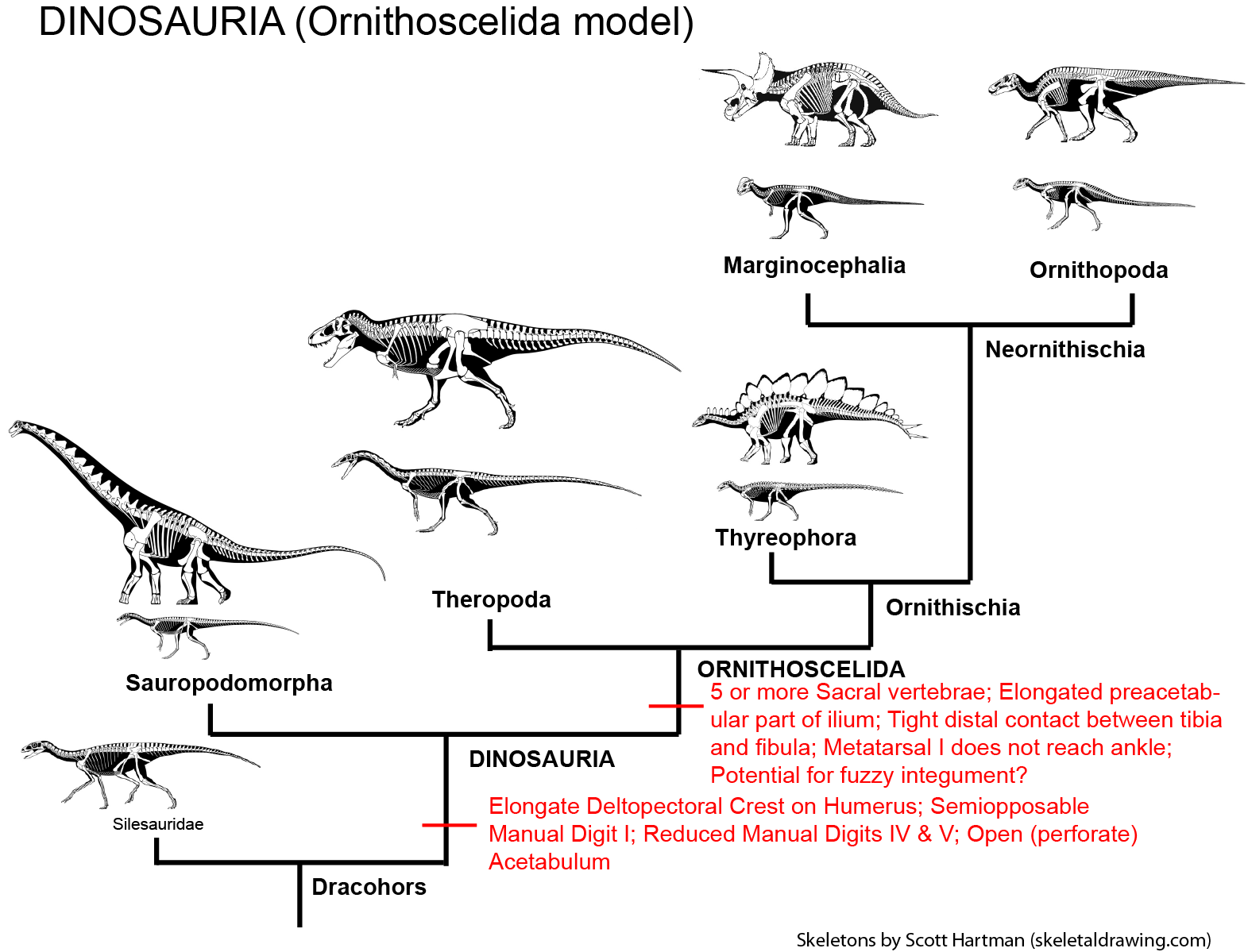
Major Alternative Dinosauria Arrangement II: Ornithoscelida: The dinosaur research community was rocked in March 2017 by a paper by graduate student Matthew Baron and his advisors. They presented evidence that Sauropodomorpha was not the sister group to Theropoda, and instead that theropods and ornithischians were closer to each other than either was to sauropodomorphs. Their analysis had a greater sample of early dinosaurs and dinosauromorphs than previous ones. The resurrected an old name, Ornithoscelida ("bird legs"), for the clade comprised of Megalosaurus, Iguanodon, and all taxa closer to them than to Sauropodomorpha. Ornithoscelida is supported by numerous (if sometimes subtle) traits of the skull, vertebrae, humerus, and hindlimb. (Among these are a metatarsal I that does not reach the ankle joint.) In this scenario Herrerasauria seems to be on the sauropodomorph line, or possibly more basal.
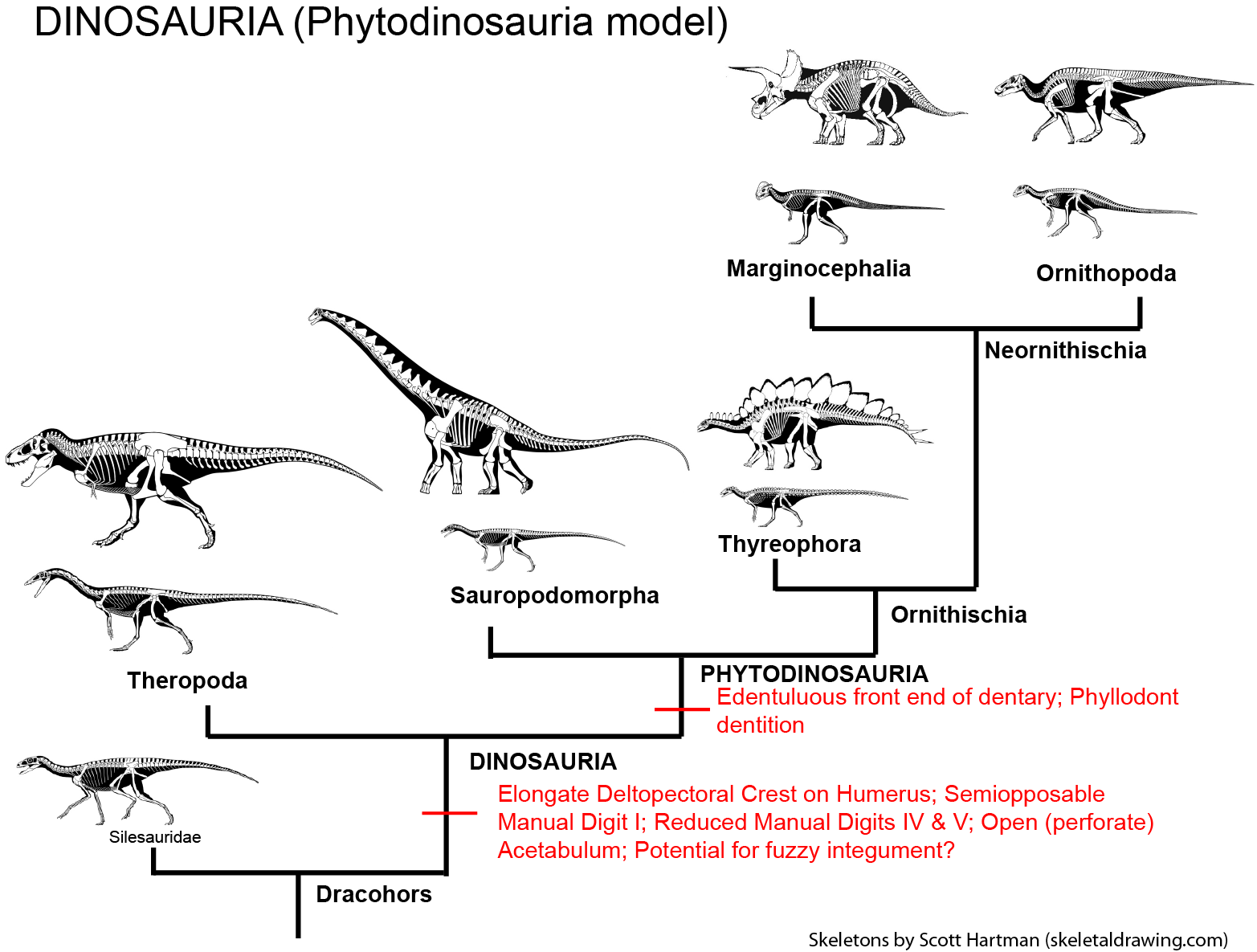
Major Alternative Dinosauria Arrangement III: Phytodinosauria: As they used to say in the old commercials, "but wait, there's more!" Back in the 1980s a possible phylogenetic hypothesis put forth by several researchers was the union of Sauropodomorpha and Ornithischia to the exclusion of Theropoda. This was given the alternative names "Ornithischiformes" or (more commonly) "Phytodinosauria" ("plant [eating] dinosaurs"). This hypothesis is based on the shared presence of phyllodont dentition and several other skull features (including a possible cheeks and beaks), associated with herbivory. A working definition for the clade would be "Diplodocus, Iguanodon, and all taxa sharing a more recent common ancestor with them than with Megalosaurus." No recent computer-generated phylogenetic analysis has supported, but possible support for this arrangement may exist in the enigmatic Chilesaurus (more about it below). In one possible variation of this hypothesis, animals currently considered basal sauropodomorphs may actually be ancestral to ornithischians, explaining the apparent absence of bird-hipped dinosaurs in the Triassic.
Chilesaurus's Relationship Status: It's Complicated: In 2015 a new dinosaur from the Late Jurassic of Chile was described. Several individuals had been found, both juveniles and adults up to 3.2 m long. Given the name Chilesaurus, this dinosaur has a suite of traits associated with different dinosaur clades. Its short skull and long neck resemble those of basal sauropodomorphs; its backwards-pointing pubis and possible beak at the end of the dentary are similar to those of ornithischians; and various vertebral, scapular, pelvic, and hindlimb traits are comparable to theropods (and especially primitive tetanurines.) Its teeth are blunt and have fine denticles, strongly suggesting a herbivorous diet: however, they do not resemble the teeth of either basal sauropodomorphs or ornithischians. The initial analysis found little Chilesaurus to be a bizarre herbivorous tetanurine theropod; alternatively, a paper in August 2017 found it to be the most primitive ornithischian (branching off before the origin of the predentary). Just throwing this out there: it would also make a great amount of sense as a basal ornithischian derived from sauropodomorph-like ancestors under the "Phytodinosauria" hypothesis. Given its position in time, either of these latter ideas require it to be a late survivor retaining the attributes of much older Triassic ancestors. In any case, an earlier representative of the Chilesaurus-lineage would be very helpful in pinning down its place in the (already destabilized) dinosaur family tree.
III. Late Triassic Dinosaurs Of Uncertain Position
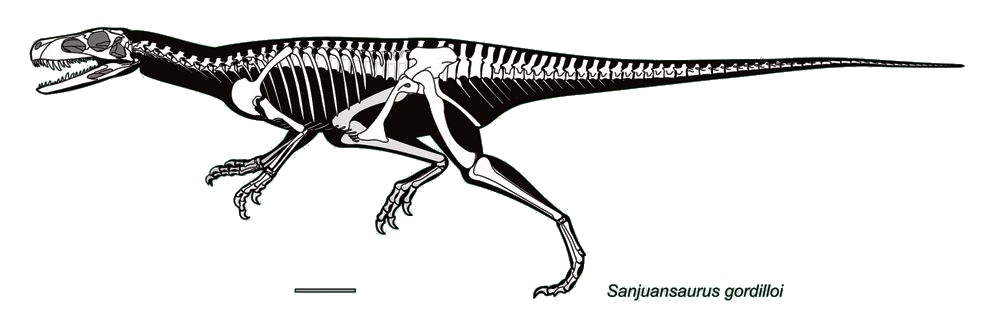
As you might notice from the further information boxes above, our knowledge of the basal relationships among dinosaurs is somewhat destabilized at present. One consequence of this is that a number of Triassic taxa cannot at present be securely placed in any of the three major clades. These include:
When our knowledge of the basal relationships within Dinosauria are better established, the positions of these Triassic basal saurischians (probably) should settle down.
Although dinosaurs appeared in the early part of the Late Triassic ecosystems, they were just a minor part. For example, other animals were far more common in the Ischigualasto Formation than Eoraptor, Herrerasaurus, and Panphagia. Theropods were not the top predators in any known Late Triassic assemblage: that distinction goes to big pseudosuchians. By the end of the Late Triassic, sauropodomorphs HAD become the largest and most common herbivores in some ecosystems (away from the equator).
Recent work shows the earliest dinosaurs and the earliest members of most dinosaur lineages showing up in the southern continents (especially South America and southern Africa) first. New discoveries from the lower part of the Chinle Formation of the American Southwest shows that diverse non-dinosaurian dinosauromorphs were still a major part of the assemblage well into the later part of the Late Triassic.
So most of the the Late Triassic is the time WHEN DINOSAURS SHARED THE EARTH!. Pseudosuchians remained ecologically diverse and important, and (especially in the early Late Triassic) non-archosaurian archosauromorphs and therapsids were also significant. In the later Late Triassic (in Gondwana and Europe, at least) things shift: pseudosuchians remain important (especially in the form of paracrocodylomorphs, aetosaurs, and phytosaurs) as did dicynodont therapsids, but sauropodomorphs expand in diversity (and size!). In recent years it has been suggested that a major environmental shift was the cause of this. Sedimentological analysis shows that the world got considerably wetter from 234-232 Ma. Its particular trigger isn't well understood, but it affected life on both land and sea. This event occurred in the Carnian Stage, and so is called the Carnian Pluvial Event ("pluvial" = "of or concerning rain"). One of the changes on land is the diversification and spread of groups of tree-sized plants (especially conifers and the now-extinct cycadeoids). It has been argued that the accumulation of this new leafy biomass may have promoted the expansion of the sauropodomorphs, and thus of Dinosauria.
(That said, the phenomenon is NOT apparent in North America! Both the excellent body fossil record of the Southwest and the trackway record of the East show no spread of sauropodomorphs, or dinosaurs in general, in North America in the later Carnian and subsequent Norian and Rhaetian stages.)
At the end of the Late Triassic, though, there was a mass extinction that affected the entire planet: the Triassic-Jurassic Extinction. The primary trigger seems to have been a period of tremendous volcanism in the middle of Pangaea. 201 Ma saw the beginnings of the rupture of the supercontinent of Pangaea, between the landmasses that are today North America, Europe, and Asia (on the north) and South America, Africa, and the other continents (to the south). This produced the Atlantic Ocean in the long term, but for the inhabitants of the Earth at the time suffered.
A vast amount of greenhouse gasses (both carbon dioxide and methane) were released, producing catastrophic global warming. But this was preceded by a burst of sulfur dioxide, which produced a rapid phase of cooling. There is additional evidence that poisons like mercury were released into the environment.
These massive pulses of environmental change were too much for many groups of organisms to survive. This is one of the five greatest mass extinctions known, with many terrestrial and marine taxa wiped out. Among terrestrial vertebrates, some of the main victims were:
With no serious competition remaining, dinosaurs radiated into numerous forms. From the beginning of the Early Jurassic onward, DINOSAURS RULED THE EARTH.
To Next Lecture.
To Previous Lecture.
To Lecture Notes.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}