

Key Points
- Homo sapiens is characterized by many behaviors which are not definitely present in other extinct human species nor in other extant primates.
- One of our primary traits is the tendency to roam: we have by far the largest geographic range of any primate.
- While Africa and Eurasia had been populated by multiple human species, but only Homo sapiens made it to Sahul (Australia plus New Guinea) and the Americas.
- Tracing humanity's spread combines paleontological (fossil), archaeological (tools), and genetic data.
- Humans have arrived in Sahul by 60-45 ka and the Americas by 14.8 ka (definitely) and possibly as early as 22 ka.
The Biblical story of Eden talks of how the first two humans Adam and Eve lived in the Garden of Eden without having to work for a living, but after the Fall lived in a world with suffering (including having to labor to feed themselves). From this couple all populations of the world descended. In fact, Genesis 10 explains how the children of Noah were the founders of the various populations around the known world of Biblical times (i.e., the Near and Middle East, and the parts of Europe, Central Asia, and Africa with which these peoples had contact in the Bronze Age.) As it turns out, however, humans were even more widespread than Genesis implied: from the Arctic to the Tropics, from rainforests to deserts. Yet everywhere, these people represent the same species.
All peoples of the world share certain behavioral and cultural traits in common, which were thus almost certainly present in the common ancestral population of crown-group humans. These are beyond Linnaeus's description of our most important trait being our intelligence (i.e., Homo sapiens, "thinking humans"), and I have tongue-in-cheek described alternative names that are probably equally good for our species:
How do we trace the spread of humans over the world? There are three primary lines of evidence:
Introduction
Behavioral/Cultural Traits Shared by All Living Human
Genetic Diversity of Modern Humans
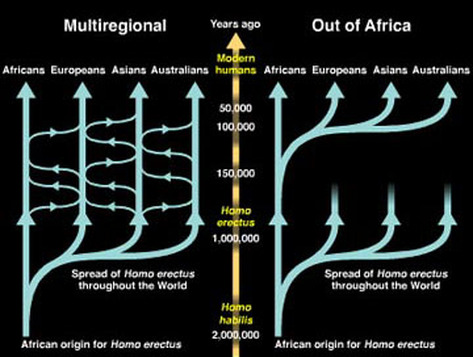
Multiregional vs. Recent Out-of-Africa Origins
In anthropological circles there have been two primary models for the peopling of the world. The first, called the multiregional hypothesis, suggests that the H. sapiens populations of Asia, Africa, and Europe were independently derived from local populations of H. erectus: Asians from classic H. erectus, Africans from H. ergaster through H. rhodesiensis, and Europeans through H. antecessor to H. heidelbergensis to H. neanderthalensis to H. sapiens. (Note that multiregionalists lump H. erectus, H. ergaster, and H. antecessor into H. erectus, and the others all as "archaic H. sapiens."
{kind=link}
This idea flies in the face of our understanding of how speciation works. If true, then "H. sapiens" would be polyphyletic! Furthermore, it suggests that the majority of the genetic heritage at each of these regions was established locally nearly 2 million years ago (although admittedly with some intermixing ever since.) For this reason, the multiregional hypothesis is also called the regional continuity hypothesis.
In contrast, the Recent Out-of-Africa hypothesis considers all living humans as descending from a relatively recent (perhaps only 50-60 kyr old) ancestral population from Africa, spreading out around the world and displacing the previous local inhabitants. (Again, there is no reason to assume that there wasn't limited intermixing.) This is much more in line with the fossil, archaeological, genetic, and other data, and with basic evolutionary biology.
For example, it has been observed that the genetic distance between members of the same troop of chimpanzees in Africa is 4-6 times the genetic distance among any given members of the 7.4 billion humans! So for all our apparent diversity, all humans are extremely closely related with a very recent common ancestral population. Some analyses suggest that we are the descendants of a genetic "bottleneck" where the effective population size of humans some 70 ka or so was a mere 2000 or so individuals, and that this small population size existed for hundreds or thousands of years.
Mitochondrial Eve, Y-Chromosome Adam, and the Genetic Phylogeny of Modern People
Geneticists have examined the genomes of people from all over the world to map out various aspects of our genetic diversity and our genetic interrelatedness. One of the important early discoveries was by looking at the mitochondria. Mitochondria are passed on only from the mother, so they can be used as markers to look down the maternal line. It turns out that the most recent common mother of all living humans seems to have been a woman who lived about 200 ka (152-234 ka), probably in Africa and long after the split between the H. heidelbergensis-neanderthalensis and H. rhodesiensis-sapiens lines. NOTE: although named "mitochondrial Eve" this would NOT have been the very first H. sapiens nor the only woman around at the time! It is simply the last mother through whom all mothers can be traced. She would have been a member of a larger population, some of whom only gave birth to sons (and the same is true of many other mothers at points in between.)
A similar discovery using the Y-Chromosome (which is passed down only from father to son) helps us find the last patrilineal ancestor of all men. Again, this is NOT the oldest H. sapiens nor the only man in his environment. The original analyses placed the so-called Y-Chromosome Adam at around 142-60 ka (and thus thousands of years younger than mitochondrial Eve!), and still most likely Africa. A report from early 2013 suggests that an individual (whose ancestry is from the Mbo people of Cameroon) pulls Y-Chromosome Adam back to 338 ka (long before the oldest evidence of H. sapiens or H. rhodesiensis!! Future work will hopefully sort this out.
Furthermore, geneticists have been able to map out phylogenies of peoples from around the world. Some of the main discoveries:
- The basalmost split among living peoples is between the Khoisan peoples of southernmost Africa and the rest of humanity
- Populations of sub-Saharan Africa (who, incidentally, represent collectively the greatest part of the genetic diversity of humans) represent a paraphyletic series to the remainder
- Similarly, peoples of North Africa are a paraphyletic series to everyone else
- Europeans represent a clade distinct from the remaining populations, which collectively represent an independent "Out-of-Africa" migration dating to about 50 ka. The other parts of this distribution are:
- The Central Asian populations are a paraphyletic series with respect to
- A cluster comprised of peoples from southern India, southeast Asia, Indonesia, Polynesia, and Australasia [which in some genographies represents a cluster closer to East Asians and the Native Americans]. [This cluster once showed up genetically as an early "Out-of-Africa" branch of Homo sapiens, breaking out at 130 ka, but now doesn't show this earlier breakout] and
- The remaining cluster: the peoples of East Asia and the peoples of the Americas
This genetic phylogeny strongly conforms to the Recent Out-of-Africa model. But examination of fossil genomes added some interesting additions.
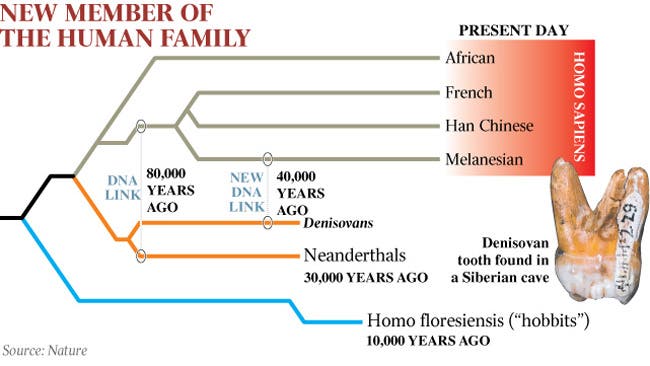
Neanderthals and Denisovan "Mystery Men"
Despite the classic "BSC" model of species definitions, it is becoming apparent that there is widespread successful hybridization among animals in the wild, including even species of mammals. Indeed, this phenomenon is common enough to produce introgression: the exchange of genes and alleles that arose in one species into part of the population of a different species. Introgression has now been documented among many types of mammals, including various species of bears, of big cats, of mysticetes, of proboscideans (including the survival of genes from extinct elephantids in the surviving species), and in the genus Homo.
Neanderthal genetic material has been recovered from numerous fossil throughout their history. Some interesting discoveries have been found:
- Overall, Neanderthal DNA falls well outside the clade of all living H. sapiens, confirming that they were a separate species from us
- Some Neanderthal alleles are distinctly different from their H. sapiens equivalent. For instance, their version of MCR1 (which produces red hair and pale skin) is very different than the vast majority of the same one found in modern humans.
- Earlier Neanderthal DNA are more distinct from H. sapiens than later Neanderthal DNA is, suggesting some interbreeding between both species after their encounter
- Genetic markers for Neanderthal genes show up in SOME but not ALL modern humans. In fact, they show up in all populations outside of Africa. This again is very consistent with the Recent Out-of-Africa model, since the ancestral population of non-Africans would pass through a region inhabited by Neanderthals at that time.
- The greatest frequency of Neanderthal genes is in Europe
- Neanderthal genetic contribution to modern humans include several variants of genes associated with skin color, hair, and some affecting genetic diseases (such as type 2 diabetes and Crohn's disease).
{kind=link}
A very recent discovery, based on 2016 analyses of the Neanderthal genome, is that there were significant differences between the Y-chromosome of Neanderthals and modern humans. This could explain why the Y-chromosome of modern humans lacks Neanderthal genes, and would mean that nearly all successful Neanderthal-modern hybrids would be between male moderns and female Neanderthals: crosses the other direction might have been fairly inviable.
Perhaps more unusual was the discovery at the Denisova site in south central Siberia, between Kazakhstan and Mongolia. Here a handful of fossils (a toe bone, a finger bone, some teeth, lots of fragments, and as of Spring 2019 a bit of skull) have been found, dated to mostly around 41 ka (but some as old or older than 51 ka and as young as 28.9 ka.) At first, nothing was known of their anatomy, but their genetic material was preserved and discovered to be closer to H. neanderthalensis than to us.
{kind=link}
Curiously, Denisovan DNA markers show up in modern humans, too, but ONLY in peoples of Melanesia, Polynesia, and Australasia. So there must have been some interbreeding between these "mystery people" and the ancestors of the southeastern branch of modern humans.
An April 2014 study of the genomes and epigenomes of modern humans, Neanderthals, and Denisovans reveals that we differ from the other species in a few traits associated with the digestive system, but especially with genes activated in higher brain function. (In fact, some of these genes, in a malfunctioning form, contribute to Alzheimer's or to schizophrenia!) These may be the genes associated with the "Great Leap Forward".
And a mystery within a mystery: there is evidence that Denisovans interbred with a population that Neanderthals and moderns never directly interbred with, and that this other group (presumably in eastern Asia) diverged long before the modern vs. Neanderthal-Denisovans split. Might this mystery population be the Dali, Jinniushan, or Homo luzonensis peoples?
Alternatively, the robust Pleistocene Asian advanced Homo species has been proposed by some researchers to BE the southern range of the Denisovan people (in which case the name might wind up being the already-existing species name Homo mabaensis or Homo daliensis or Homo longi, assuming derived features can be found in these type specimens.) This would be consistent with the widespread presence of Denisovan genes in populations in Oceania; at least this is far closer than south central Siberia!! Also, this would extend the range of Denisovan fossils down to about 300 ka.The latest data suggests that all these East Asian forms are a clade that includes Denisovans, but how many species they represent remains uncertain.
But what about the fossil evidence of the spread of humans?
Brave New Worlds
Finding the oldest humans of a region is often difficult. After all, much of the world is not in D-World (depositional world) all the time. Furthermore, the first arrivals are probably of small populations. Additionally, the ability to date sites to the very fine scale to demonstrate the "REAL" first can be difficult, particularly for terrestrial depositional environments. So we face a reverse Signor-Lipps effect: the first fossils and artifacts of humans in a region are almost certainly NOT from the first people to arrive there.
Here is a traditional map for the timing of the spread of Homo sapiens. So in general, we see:
{kind=link}
- Australia colonized perhaps as early as 50 ka, and definitely by 46 ka
- Europe colonized relatively later, at 45-40 ka (except for the recently recognized skull from Greece at 210 ka!)
- East Asia around 80 ka (South China) and maybe around 40-36 ka (Northern China)
- Boreal Asia (northern Siberia, requiring the ability to make heavy clothes is developed) at 45 ka
- Alaska (north of the ice) between 26-14 ka
- And finally the Americas south of the ice around 13.5 ka definitely, but now mounting evidence for even earlier (see below).
But let's take at the finer scale details:
Sahul: Sahul is a name given to the landmass comprised of New Guinea, Australia, and Tasmania (but NOT New Zealand: thus, it is a subset of "Australasia"). During glacial maxima, there are direct land connections between all three landmasses, although the Sahul is still isolated from the Malay Peninsula (the fully-exposed region of Indonesia). The oldest well-dated human remains in the Sahul are 46 ka (previously considered 41-40 ka) specimens from Lake Mungo. There are other fossils that have been dated as old as 68 ka, but the uncertainty of these measurements are very high. However, arrival of humans in the 60-45 ka range seems most likely. A newly discovered chip of an axe at around 49-45 ka is consistent with the Lake Mungo body fossil date.
East Asia: Northern China by 40-36 ka, but some teeth from South China from over 42 ka and probably as old as 80 ka.
Boreal Siberia: 45 ka.
Alaska: possibly as old as 28 ka, but definitely by 14 ka. Note that during the Last Glacial Maximum the conjoined ice sheets from the Rockies (the Cordillera ice sheet) and eastern Canada (the Laurentide ice sheet) blocked Alaska from the rest of the America. In contrast, though, the low sea level meant that there was a continuous land bridge between Siberia and Alaska: the land of Beringia.
{kind=link}
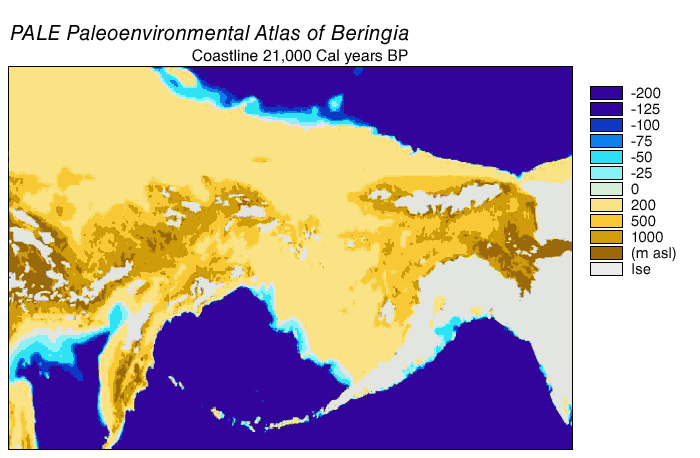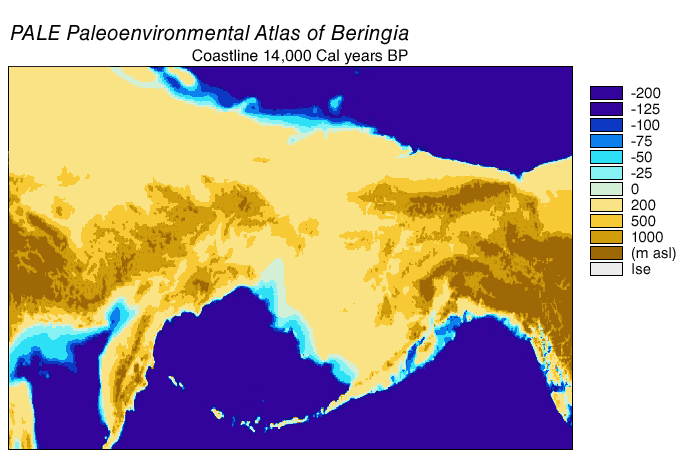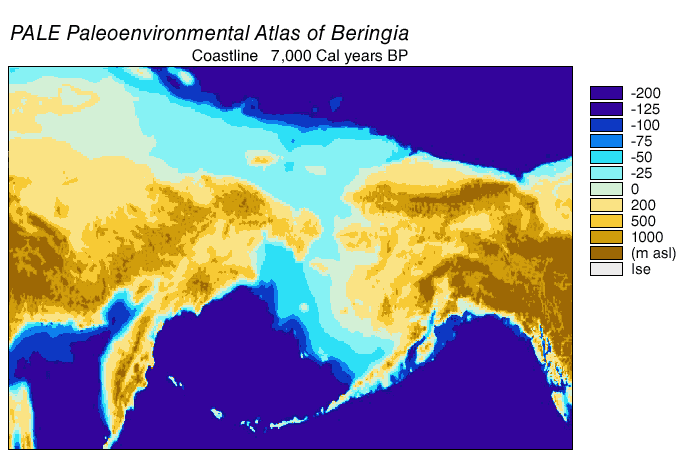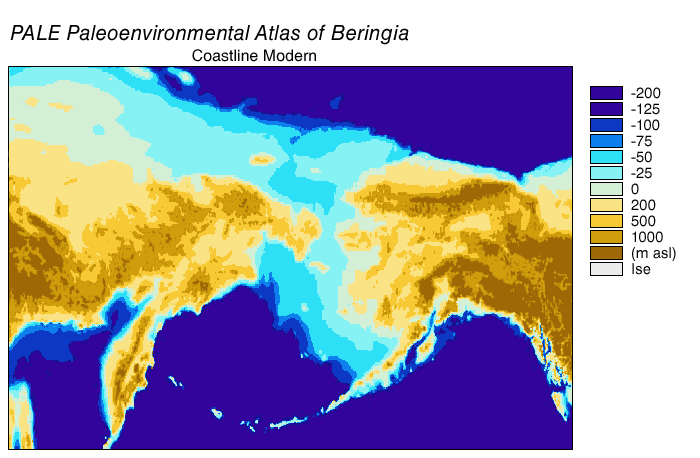{kind=link}
Americas South of the Ice: Here is place of great debate. The traditional model was that the oldest presence of humans south of the ice was around 13.5-13 ka, when an "Ice-Free Corridor" opened up between the Cordilleran and Laurentide ice sheets through which the Paleo-Indians (as the ancestors of the Native Americans have been called) arrived (somewhere near present day Calgary, Alberta). Furthermore, the traditional model considered that these people quickly developed the Clovis tool kit with it's distinctive spearpoint for heavy spears developed to take down very large prey. This Clovis toolkit was widespread in central North America for a flash-in-the-pan: 13.2 to 12.7 ka. Afterwards, it was replaced by the Folsom toolkit (once thought to be as young as 10.5-8 ka, but brand new relating shows lasted merely from 12.61 to 12.17 ka) with thinner spear points for javelins and darts, thrown with atlatls. These were adapted for smaller (bison or smaller) prey. When Folsom disappears, it is replaced by many different diverse cultures.
{kind=link}
{kind=link}
{kind=link}
But over the last few decades a number of sites have been discovered which put doubt on traditional "Clovis first". Among these are the Manis site in Washington State, well-dated to 13.8 ka (and thus pre-Clovis), where a a hunting point was found in the vertebra of a specimen of Mammut, and the newly-dated Page-Ladson site dating from 14.55 ka in the Aucilla River, Florida, which also has butchered mastodons. Other sites with good dates in the 14.8-14.1 ka range are found in the Americas; moderate levels of support for dates in 15-16.8 ka; and possible dates as old as 22 ka. (Some problematic dates older than 30 ka, but there are concerns that the material being dated is non-human [for example, forest fires rather than charcoal pits].)
{kind=link}
In 2021 exciting new evidence strongly supports the presence of humans in North America during the Last Glacial Maximum. A series of tracksites in White Sands National Park, New Mexico, are radiocarbon dated to between 21 and 23 ka. Interestingly, most of these tracks are of small individuals, mostly teens and younger.
But if the ice blocked the way, how would these pre-Clovis people arrive? They could have arrived prior to the coalescence of the Laurentide and Cordilleran ice sheets (which might have been as late as 24 ka). Recall that we are "Homo maritimus": it may be that coastal fishers rather than walking hunters were the First Americans. In fact, if the latter, some of the First Americans might not have been from Asia along the Pacific Northwest, but from Europe to Greenland to eastern North America. (In fact, there is a European toolkit called the Solutrean from 22-17 ka which resembles the Clovis, although this could very well be convergences for hunting similar animals in a similar style.) Furthermore, the genomes of a human skeleton associated with Clovis tools shows the characteristics of Native Americans (and more broadly Asians) rather than Europeans.
{kind=link}
A new model is that coastal fishers moved down the Pacific seacoast south of Beringia before 22-23 ka or so, and reaching as far south as Monte Verde, Chile by 14.6 ka. These populations began to move inland and encountering big game, eventually developing Clovis tools by 13.2 ka. There might have been a second wave of immigration at or just before this time as the Ice-Free Corridor opened up.
So, regardless of when they arrived, what happened as humans first arrived in the Brave New Worlds of Sahul and the Americas? We'll see that in the next lecture.
Some Relevant Videos