


Key Points:
•Most of metazoan diversity lies within Bilateria, the "worms" (very broadly defined.)
•Bilaterians divide into two major clades: Protostomia (arthropods, annelids, mollusks, brachiopods, bryozoans, and their allies) and Deuterostomia (chordates, echinoderms, hemichordates, and their allies). The division is recognized on the basis of embryology, development, anatomy & genetics.
•There is molecular and some fossil evidence indicating that Bilateria was present and diverged into its subclades within the Ediacaran Period (the end of the Neoproterozoic Era). However, most of the macroscopic fossils of the Ediacaran do not seem to belong to any bilaterian group. Instead, at least some represent body plans (and lineages?) which may have died out at the end of Ediacaran.
•The "Cambrian Explosion"--the appearance of diverse macroscopic biomineralized fossils--was not a sudden event. Biomineralization occurs independently among diverse bilaterians and other metazoans starting in the latest Ediacaran throughout the earlier parts of the Cambrian. It seems that small biomineralized parts (either whole tiny skeletons or individual sclerites [armor pieces] on larger animals) were easier to construct first, and that complete larger skeletons only happened later.
•This pattern is consistent with hypotheses that the driver for the ability to form larger skeletons was extrinsic of the organisms themselves, and had to do with changes in the global marine environment. The exact nature of this geochemical change remains a matter of debate.
•Regardless of the cause, the rise of biomineralized skeletons allowed animals to interact with each other and with the their non-living environment in a great diversity of new ways; once this began, the main driver of the diversification among animals was natural selection itself.
On the sudden appearance of groups of Allied Species in the lowest known fossiliferous strata. "There is another and allied difficulty, which is much graver. I allude to the manner in which numbers of species of the same group, suddenly appear in the lowest known fossiliferous rocks. Most of the arguments which have convinced me that all the existing species of the same group have descended from one progenitor, apply with nearly equal force to the earliest known species. For instance, I cannot doubt that all the Silurian trilobites have descended from some one crustacean, which must have lived long before the Silurian age, and which probably differed greatly from any known animal. Some of the most ancient Silurian animals, as the Nautilus, Lingula, &c., do not differ much from living species; and it cannot on my theory be supposed, that these old species were the progenitors of all the species of the orders to which they belong, for they do not present characters in any degree intermediate between them. If, moreover, they had been the progenitors of these orders, they would almost certainly have been long ago supplanted and exterminated by their numerous and improved descendants.
Consequently, if my theory be true, it is indisputable that before the lowest Silurian stratum was deposited, long periods elapsed, as long as, or probably far longer than, the whole interval from the Silurian age to the present day; and that during these vast, yet quite unknown, periods of time, the world swarmed with living creatures." -- Chapter 9 "On the Imperfection of the Geological Record", On the Origin of Species, Charles Darwin, 1st Edition (1859)
BILATERIA: CONQUEROR WORMS
We have covered the (probably) paraphyletic Porifera (sponges) and the (almost certainly) paraphyletic "Radiata" or "Coelenterata" (Cnidarians and Ctenophora) in previous lectures. We now move up into the vast majority of animal diversity: the Bilateria, or "worms, broadly defined". Bilaterians are ancestrally characterized by:
Evidence in the Ediacaran for Bilateria as a whole is compelling, but the exact nature of which bilaterian groups are present is problematic at present.
Bilaterian Embryology
Historically, an understanding of embryonic development has been key in recognizing the major groups of Bilateria. When we left off with diploblastic animals we saw embryos that underwent cleavage to form a blastula (ball of cells with a hollow center or blastocoel) which underwent gastrulation (invagination) to produce a gastrula with an opening (blastopore) to the ancestral gut archenteron). The cells lining the archenteron are the endoderm, and typically become the digestive tissues in cnidarians, ctenophores, and bilaterians. The lineage of cells that remain on the outside of the gastrula are the ectoderm, and are associated with the external tissues of eumetazoans.
Now for something completely different: cnidarians and ctenophores have only these two cell lineages, and are called diploblastic. But bilaterians produced a new cell line that forms between these two, called mesoderm, which goes on to produce many of the remaining complex 3D organs of the body. Thus, eumetazoans are triploblastic. (Okay, some cnidarians have a mesoderm-like tissue called entocodon, but developmental biology suggests this is convergent rather than synapomorphic with true mesoderm.) Perhaps not coincidentally, this new raw material (mesoderm) shows up in the phylogeny at the same time that a full compliment of regulatory and related genes (HOX) also does.
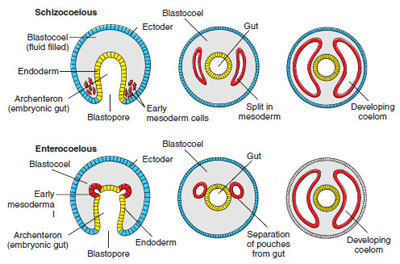
The vast majority of bilaterians have a coelom (or body cavity) in which the guts are suspended and which can act as a hydrostatic skeleton. There are some flatworms which are acoelomate (without coelom), where the body tissues are solid ectoderm --> mesoderm --> endoderm. There are also nematodes which are pseudocoelomate, where a false-coelom forms between the mesoderm and the ectoderm. But the vast majority of bilaterians are eucoelomate, with a true coelom that forms WITHIN the mesoderm (and in which mesodermally-derived mesenteries help to suspend the 3D organs in the chamber.)
Eucoelomy is achieved in two different major pathways, which point to the two-fold division of the major groups of Bilateria:
Classically, though, the names of these two grade clades are based on the fate of blastopore and origin of the mouth:
Some have tried to use nerve cord development as a nice way of dividing up Deuterstomia and Protostomia:
So with all due recognition that the following statements are inaccurate (because there are exceptions to almost every single point!), we classic recognize two major clades within Bilateria with the following features:
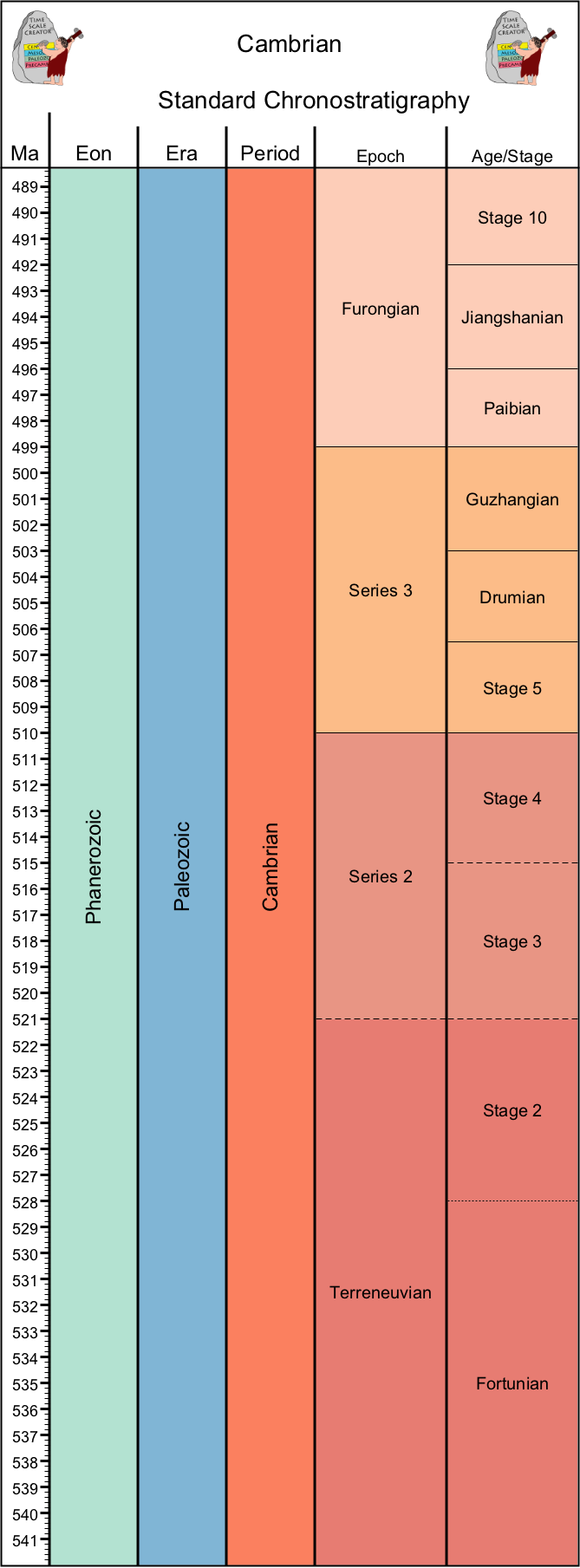
The Cambrian Period (541-486 Ma) is the first period of the Phanerozoic. It was traditionally recognized by the oldest known specimens of fossils with skeletons (of silica, calcium phosphate, or [most commonly] calcium carbonate) that can be seen by the naked eye. More intriguingly, instead of just one species ancestral to all other skeletonized forms, many diverse clades simultaneously show up in the record. This was a useful biostratigraphic marker, but a puzzle to evolutionary biologists.
That is not to say that there were no known Precambrian fossils; just no fossils of macroscopic organisms. One false start was "Eozoon" (which turned out to be a metamorphic rock). However there are similar-looking structures common in the Precambrian (and Cambrian) called stromatolites. These layered structures are the byproducts of biofilms (aka "slime") created by prokaryotes, catching particles of sediment and producing mounds in tidal zones and similar environments. Modern stromatolites only form where the water is too harsh (normally to saline) for animal life, but earlier they formed in a wider variety of environments.
The position for the base of the Cambrian--the stratigraphic marker to be used--was debated during the middle of the 20th Century. It had been recognized that below the Cambrian Explosion proper (first trilobites, first big mollusks, etc.) there were other skeletonized fossils: archaeocyathid sponges and an assortment of tiny calcareous fossils of various shapes nicknamed the "Small Shelly Fossils" (or "Small Shelly Fauna", or "SSF"). And below those there are the first trace fossils that dip down into the sediment and back out again: Treptichnus pedum (once called "Phycodes pedum"). Recent work strongly suggests that Treptichnus pedum was generated by priapulid worms (known to have been fairly common in the Cambrian). If priapulids were present by the time T. pedum, then the diversification among metazoans was well underway by that point and we have to look for the origin of animals in the Proterozoic Eon.
In 1992 it was proposed (and ratified in 1994) to use a location near Fortune, Newfoundland, Canada as the site for the formal base of the Cambrian. The horizon picked was one that was (at the time they ratified this) the lowest occurrence of Treptichnus pedum at that location. Under this model, the SSF appear after this, as do the first archaeocyathids, and then the great diversification that is the Cambrian Explosion.
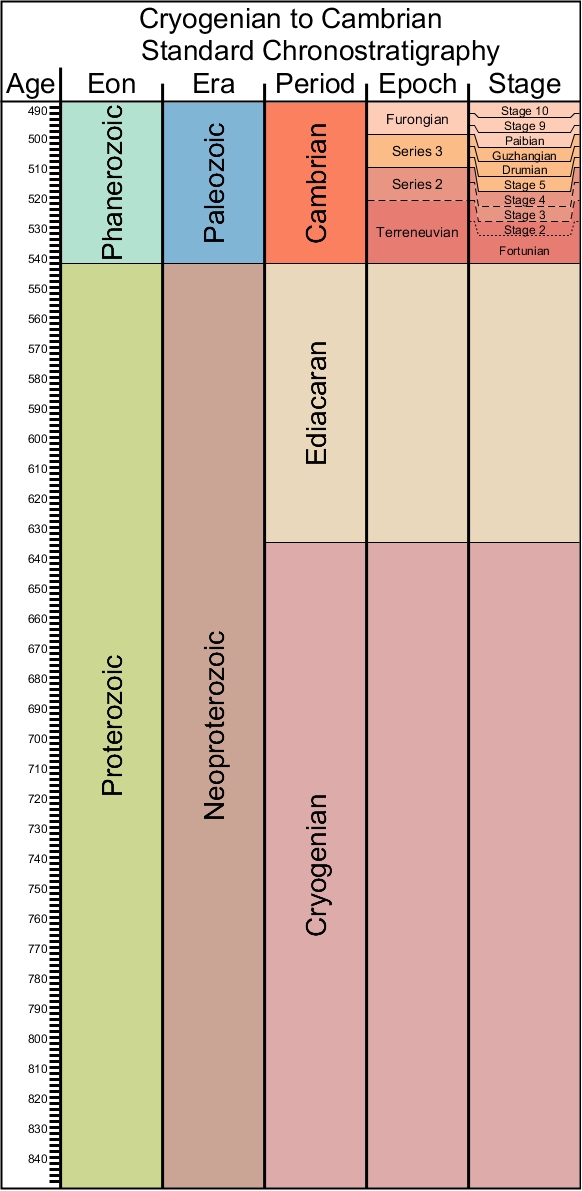
What is the evidence for multicellular organisms in the Proterozoic Eon? Macroscopic colonies of organisms (possibly prokaryotes, more probably eukaryotes, called Grypania) are known from 2.1 Ga. Likely multicellular red algae (Bangiomorpha) are present by 1.6 Ga. Complex communities of multicellular algae are present by the Ediacaran. But what about metazoans?
We should consider the following questions when addressing issues of putative Proterozoic animal fossils:
Below is a simplified cladogram of the Metazoa:
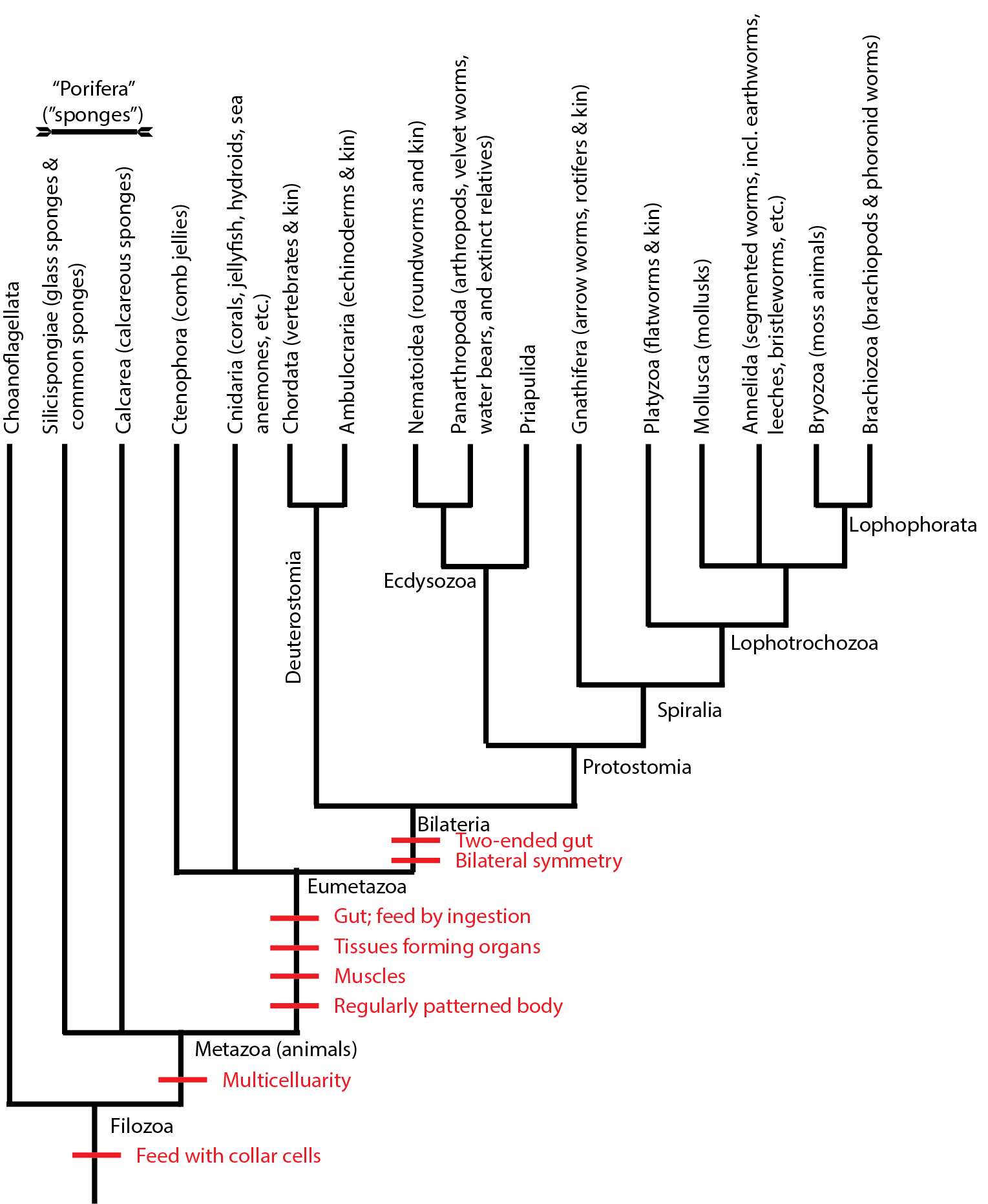
Note that "sponges" (the paraphyletic grade comprised of Silicispongiae and Calcarea) are the basal branches of metazoans. One would expect that they would be the first to show up in the fossil record, as in fact they do:
How about other groups? One intriguing bit of evidence from the 1800 Ma (1.8 Ga) Stirling Range Formation: trace fossils that seem to indicate a macroscopic animal moving along the surface of the sediment, pushing it out of its way. This led to great debates about the age of the Stirling Range Formation, but it does indeed seem to be quite old. However, discovery that the macroscopic multinucleate amoeba Gromia leaves trails as it very slowly moves around which look just like the Stirling Range Formation tracks undermines these ichnofossils as evidence of early worms.
Another intriguing line of evidence that was of great interest over the last 20 years are the Doushantuo "embryo" fossils. This formation (which runs from the base of the Ediacaran at 635 Ma to about 551 Ma at the top: the fossil-bearing layers are concentrated from 600-590 Ma) preserves many microfossils and small macrofossils (including algae, sponges, tubular fossils, and so forth), primarily by a phosphatized coating. Some of the fossils resembled the dividing cells of cnidarian and bilaterian embryos, although neither late-stage embryos nor adult organisms ever showed up. Although some still consider these embryos to be evidence of metazoans, an alternative hypothesis strongly suggests these are actually the dividing stages of unicellular organisms. Whether these belong to the stem-lineage leading to animals, to choanoflagellates, to fungi, to plans, or to any other of the diverse groups of eukaryotes has yet to be established. So like the Stirling Range Formation ichnofossils, the Doushantuo embryos might well be produced by something that aren't animals.
Newly discovered: in November 2020 the first case of these "embryos" has been reported outside the Doushantuo Formation. Extremely similar fossils from the same-aged Portfjeld Formation of Greenland have now been described. Again, they are found without any hint of supposed larval or adult organisms in the same formation!
The Ediacaran Biota: The actual evidence of Proterozoic animals (even macroscopic animals) was actually first discovered some time ago. In 1872 fossil impressions given the name Aspidella terranovica were described from St. Johns, Newfoundland, Canada. These simple sets of concentric rings were not particularly convincing as animal fossils to people at the time (instead, they were quite reasonably dismissed as some kind of abiogenic sedimentary structure), but in hindsight were eventually realized to be the first described Neoproterozoic metazoans.
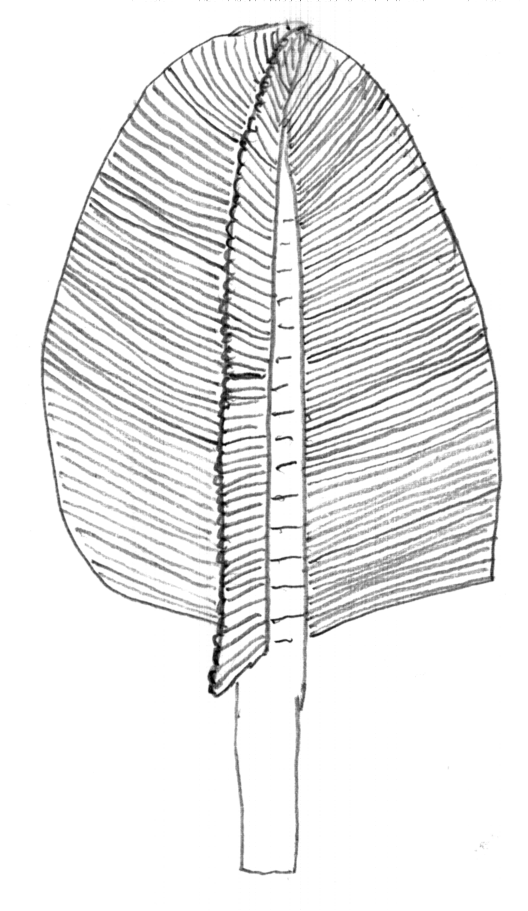
In 1957 a young kid walking in Charnwood Forest, UK discovered a leaf-like fossil in the Precambrian rocks present there. This discovery became recognized as Charnia. Because the geology of the UK was so well-studied, it was definite that these were from below the Cambrian, and the news went worldwide of the discovery of macroscopic Precambrian animals.
As it turns out, however, even better fossils of a diverse community of organisms had been discovered and published back a decade earlier. In 1946 Australian mining geologist R.C. Sprigg was exploring the Ediacaran Hills of Flinders Ranges, in Australia. These only reached international attention, however, in 1961, which is why the Charnwood fossils are generally regarded as the "first discovered" Precambrian animals. The great diversity of forms found at Ediacara led to all fossils of this variety being lumped under the name "Ediacara Biota". Such fossils have been found in many locations around the world (other than those already mentioned, they include the White River localities in Russia and the Nama locality in Namibia).
Preservation of the Ediacaran Biota: The Ediacaran organisms are macroscopic: some up to a meter long or so, but most much smaller. They are primarily preserved as impressions in sediments (sometimes shallow-water coarse sandstones, sometimes deeper water shale). In some cases the outside skin surface is preserved by permineralization or replacement. In many cases a "death mask" of pyrite formed during decay, which aided in preservation.
From the 1960s until the 1980s (and by some researchers even into the 21st Century), these fossils were interpreted as early representatives of well-known Phanerozoic groups like cnidarians, flatworms, segmented worms, arthropods, etc. However, paleontologist Dolf Seilacher and his colleagues pointed out the following aspects about that many of the fossils of the Ediacaran Biota do not match our expectations for members of any of the living groups of animals (or at least eumetazoans):
Seilacher named this quilted organisms the "Vendobionta". It is not at all certain that these form a single group. More about them below.
Not all Ediacaran Biota organisms fit the above profile. We will deal with the Ediacaran stem-mollusk Kimberella and the cone-dwelling cloudinomorphs later. There are some Ediacarans that do match some Phanerozoic groups of non-bilaterian animals:
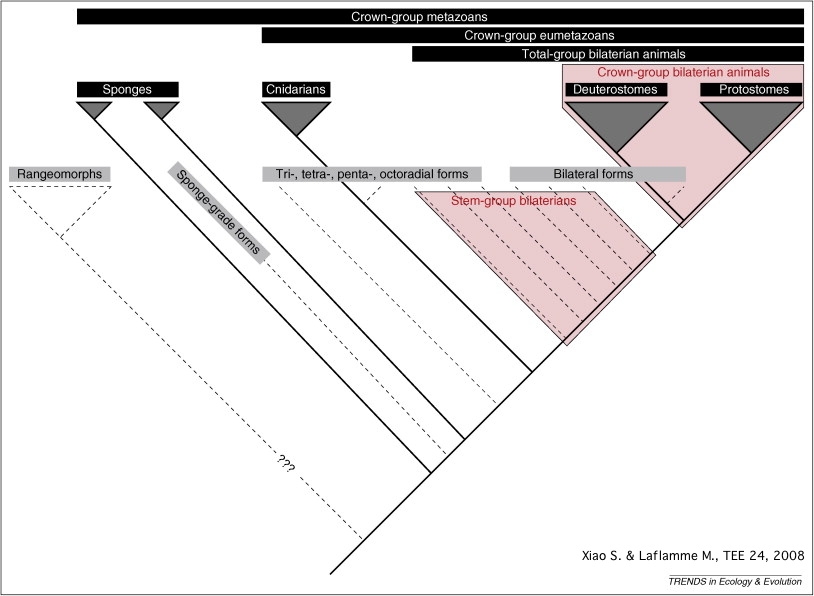
But what about the rest of the Ediacaran critters? Seilacher argued that these organisms formed their own clade (Vendozoa or Vendobionta) separate from Metazoa. However, the patterning of these bodies are more like animal growth than most other multicellular animals: indeed, having a "front" and "back" throughout development in some forms suggests at least some HOX genes were present. Recent work by S. Xiao and M. Laflamme suggests that different Ediacaran forms represent different stem-group parts of the metazoan tree. (In fact, under this model, the ancestors of the bilaterians would have passed through an "Ediacaran biota" phase.)
Recent biomarker studies have demonstrated that the classic "vendobiont" Dickinsonia possessed cholesteroids, and thus belongs to the metazoan clade. (Some other Ediacaran organisms lack traces of this substance; instead, biomarkers indicate that Palaeopascichnus was a giant protist, and Beltanelliformis was a cyanobacteria colony.)
New work suggests the vendobionts have several possible subgroups. (That said, there is no indication any of these are actually monophyletic, nor that "Vendobionta" is actually a clade):
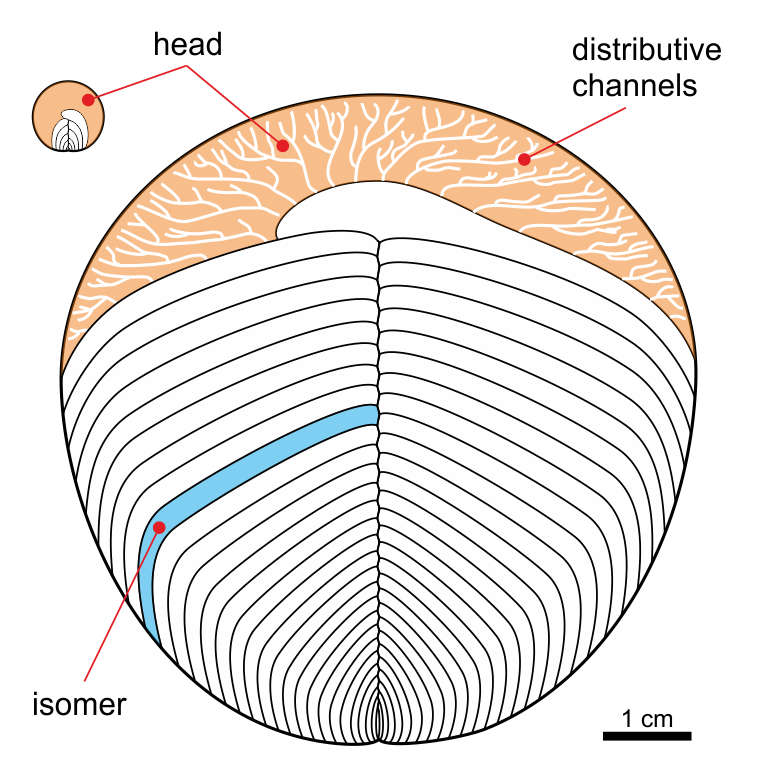
Kimberella: One of the Ediacaran organisms stands out as being neither a cnidarian nor a "vendobiont"-type animal, but a probably crown-group bilaterian. This is Kimberella (commonly preserved as a belly print, but occasionally preserved showing the top surface). It is often associated with scrape marks suggesting a radula scraping up algae from the algal mat. Based on the preserved data, Kimberella appears to be a stem-mollusk, sharing some but not all traits common to the concestor of living Mollusca. In particular, it appears to have:
If Kimberella is indeed a stem-mollusk, then by minimum divergence time it requires that stem-annelids, stem-platyzoans, stem-brachiozoans, stem-bryozoans, stem-chaetognaths, and stem-deuterostomes must also be present in the Ediacaran. And thus, the major divergences among Bilateria occurred long before the Cambrian Explosion.
Some other possible Ediacaran bilaterians have been described in recent years:
Ediacaran Calcifiers: While the vast majority of fossils of the Ediacaran are soft-bodied, there are a few very small organisms with hard parts. Coronacollina (mentioned above) seems to have had siliceous spicules, but these are not preserved. There are a few calcifying organisms as well.
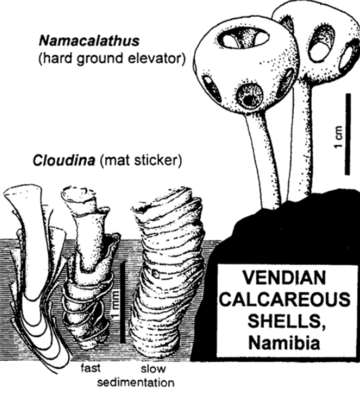
One of these is Namacalathus, tiny spheroids with holes on the side, set atop a small stalk. These seem to be some form of suspension feeder: indeed, they are quite likely some form of calcifying sponge. Larger than Cloudina, they are nonetheless very, very, very small. (Another calcifying organism--Namapoikia--seems to be an encruster, but is not clearly a metazoan: it could be a unicellular form.)
Cloudinomorphs: The most diverse of the Ediacaran calcifiers are the cloudinomorphs. The first discovered of these was Cloudina: small cone-in-cone calcitic tubes, most likely formed by some kind of eumetazoan. These are VERY tiny, and so it isn't a surprise they were missed for so long. The shells show a series of cones in cones.
Until recently all that was known of the cloudinomorphs was their shells. They seemed to be slightly flexible, and were arranged over the algal mats. Indeed, they seem to enter and leave the mat. Whether the animal inside was a suspension feeder or relied on chemosymbiosis is still being examined. New discoveries in early 2020 shows that the animal inside had a gut. Whether this animal was a cnidarian (and thus it was a blind gut) or a worm-like organism (with a flow-through gut) isn't at all certain. If the latter, the animal resembles some of the polychaete annelid worms.
Recent (March 2017) discoveries finds that the calcifiers (including cloudinomorphs) are only found in carbonate rock, and that they all have nearly-identical soft-tissue versions in detrital sedimentary deposits. It appears that the same organisms produce unshelled structures where carbonate minerals are not abundant, but can calcify when limey mud (and carbonate rich water) is around.
If you wish to explore a slab of Cloudina-bearing limestone from Namibia (currently being studied by Dr. Jay Kaufman of the Geology Department and his students), you can look at a very high resolution scan here.
Here is a summary of the attributes of the "vendobiont", Kimberella, and cloudinomorph biology:
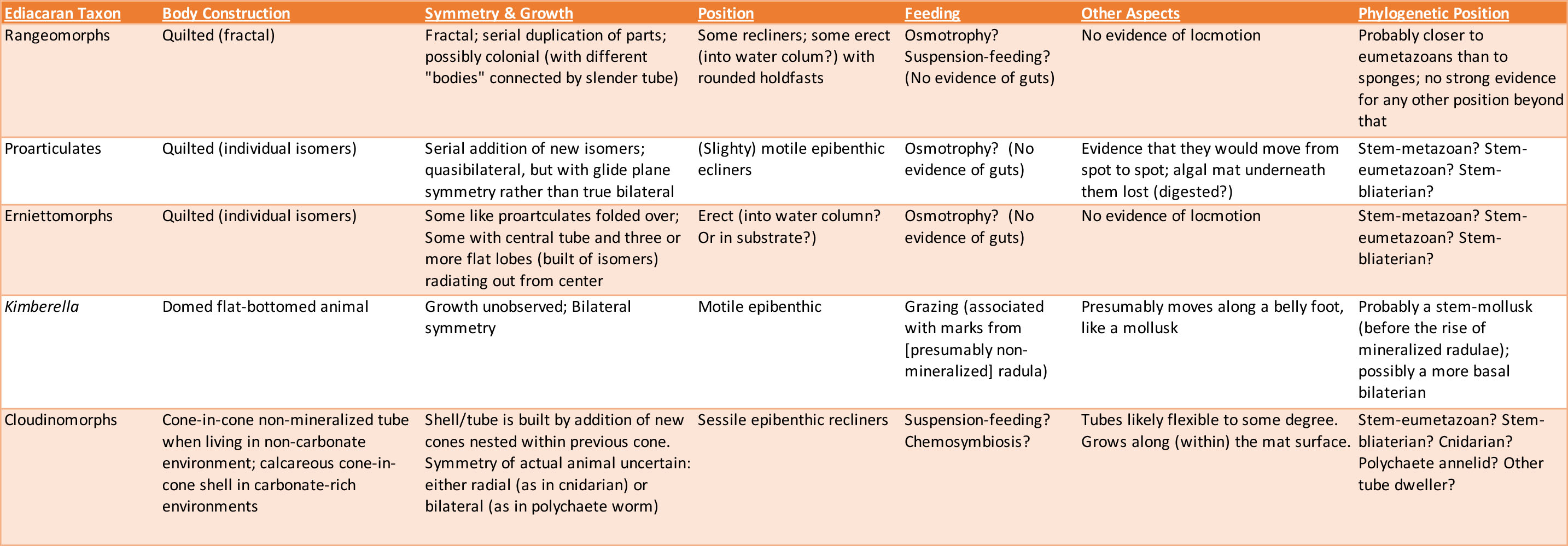
Lifestyles of the Ediacaran Biota: How did the Ediacaran biota make a living? Some are easy to figure out: Kimberella seems to have been an algae-scraper (the ancestral condition for Mollusca), and cnidarianss and sponges as suspension feeders. But what of the other forms?
The Ediacaran was still part of the Time of the Slime: algal and bacterial mats covered much of the surface of the sea floor. This matground provided both a habitat and food source for a number of niches in the Ediacaran community. There are some (often larger) Ediacaran forms that seem to have sat on the mat, moved slightly, sat again, moved again, and so forth. It appears that the mats underneath each of these was digested (although some argue it simply died by being cut off from sunlight & nutrients). This suggests that at least some of these were osmotrophs: feeding by absorbing nutrients over the body surface rather than ingesting into a gut or similarly engulfing food particles.
If osmotrophs were present in many of these Ediacaran forms, this could explain the lack of mouths, guts, anus, etc., in these impressions: they may not have been a digestive tract at all. Osmotrophy is different from the feeding in sponges but also in eumetazoans. (There is a very poorly known group of tiny animals today, the Placozoa, which feed by osmotrophy.)
So the paleoecology of the Ediacaran community consisted of a number of forms:
Predation seems to be absent or at least very, very rare, so some refer to this time as the "Garden of Ediacara" (in analogy to the Paradise of the Garden of Eden).
Let's revisit the phylogeny of Metazoa, and see where they Ediacaran forms might fit:
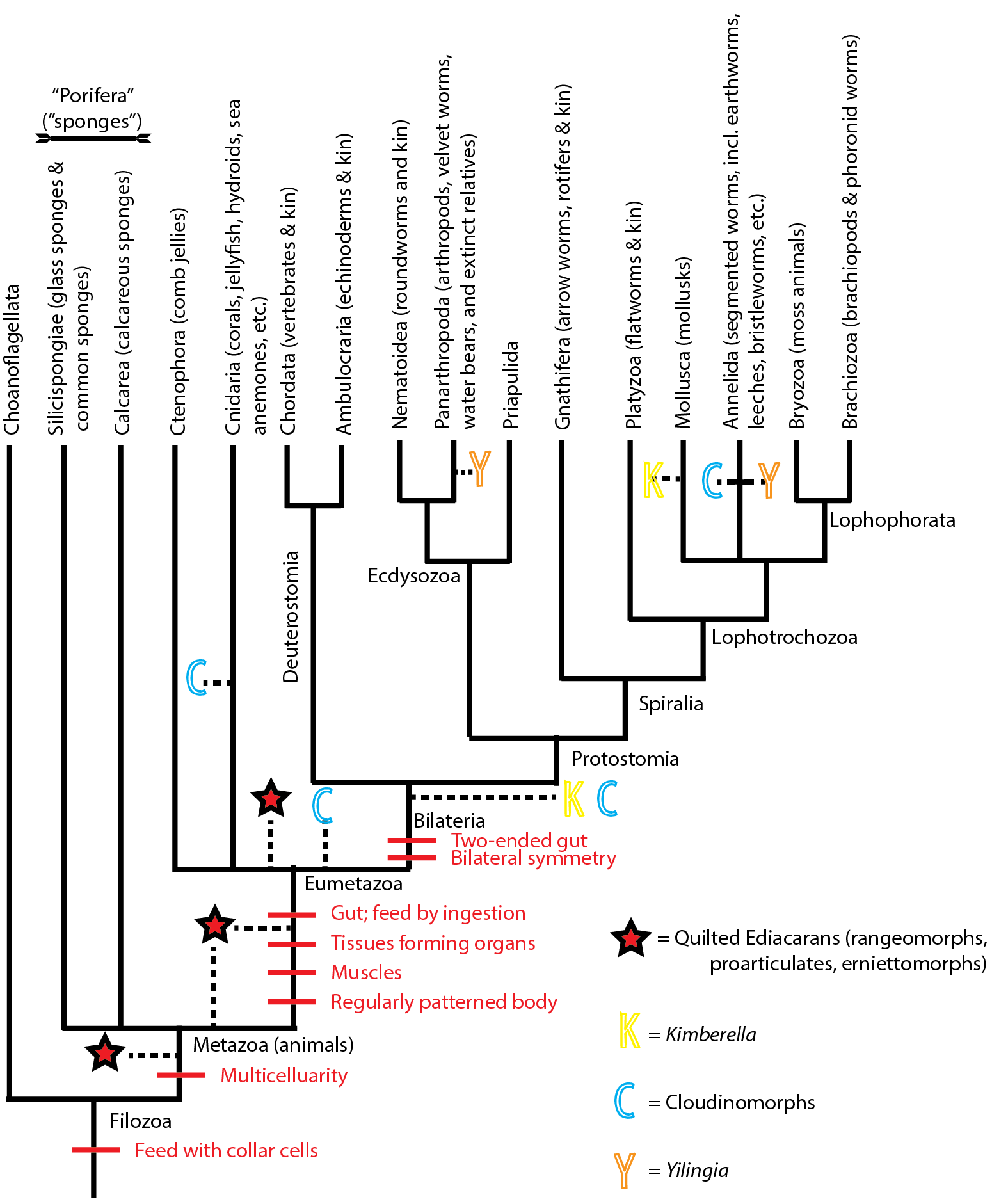
Kimberella might be a stem-member of Mollusca, or a stem-bilaterian, or many positions in between. Yilingia might be a stem-panarthropod or a stem-annelid. Ikaria remains too uncertain to place anywhere, so it is not shown. The vendobionts quilted Ediacaran forms seem to be multicelluar. Some might be stem-metazoans. Since many of them show regularly patterned bodies, and at least some show signs of movement (and thus muscles), but none show guts, they might be stem-eumetazoans. (An alternative is that they are crown-eumetazoans that lost their guts). Cloudinomorphs might be cnidarians, or basal metazoans, or polychaetes, or something else all together.
Proterozoic Oceans and Increasing Oxygen Levels: The oceans of the Neoproterozoic seem to have been considerably different than the oceans of the Phanerozoic. Modern oceans are largely clear, well mixed (with lots of oxygen distributed through out), and contain complex food webs of animals, algae, phytoplankton, and so forth. In contrast, Neoproterozoic oceans seem to have been cloudy with bacteria and organic matter, very stratified (layered with not much mixing) with a lot of anoxic levels, and very simplified ecosystems.
Chemical evidence in the sedimentary record shows that the Neoproterozoic was a time of increasing oxygen levels. This may have been the spark leading to the multicellular algae and metazoans: large, complex eukaryotes require a lot of oxygen to operate. It may be that there is a positive feedback loop involved: increasing oxygen favors diversification of animals (some of which are suspension feeders) --> cleaner water --> success of eukaryote algae --> more oxygen produced than by cyanobacteria --> increased size and diversification of animals --> and so on.
Serpents in the Garden: At the very very end of the Ediacaran, new types of trace fossils show up. The first burrows descending somewhat into the sediment are present. These include Treptichnus and a few other similar forms, and are consistent with tunnels dug by priapulid worms. Priapulids are known to be predators, and so we have a spreading of carnivores at the end of this interval. Given how vulnerable most of the Ediacaran forms seem to have been to predation, this may have been the death knell of the Ediacaran biota. (In fact, given that the Ediacaran biota forms are not present in the Cambrian (one suggested Ediacaran-style frond in the Cambrian turns out to be a cnidarian with mouths), this may have been the first mass extinction for Metazoa.)
If these are priapulids, that means stem-priapulids are present before the start of the Cambrian, which requires stem-members of the Panarthropoda-Nematoidea clade being present.
So here we are at the base of the Cambrian:
But where is everything else?? We get to the base of the Cambrian, and still haven't solved the problem of the Cambrian Explosion! That will be the next topic we get to.
So here we are at the base of the Cambrian:
So where are the missing worms? It is not at all unreasonable to predict that they are part of the meiofauna: organisms barely visible to the naked eye, in about the same size range as grains of sand. Such taxa are exceedingly unlikely to be preserved, and yet today make up a sizable fraction of the marine benthic biomass. The presence of some protostome meiofauna in Cambrian Lagerstätten is consistent with that.
Wonderful Life
Let's review the metazoan phylogeny, and now map onto it the basic mode of feeding present in the basal forms of each
terminal clade, and the most parsimonious changes at the interior nodes when they would have changed:
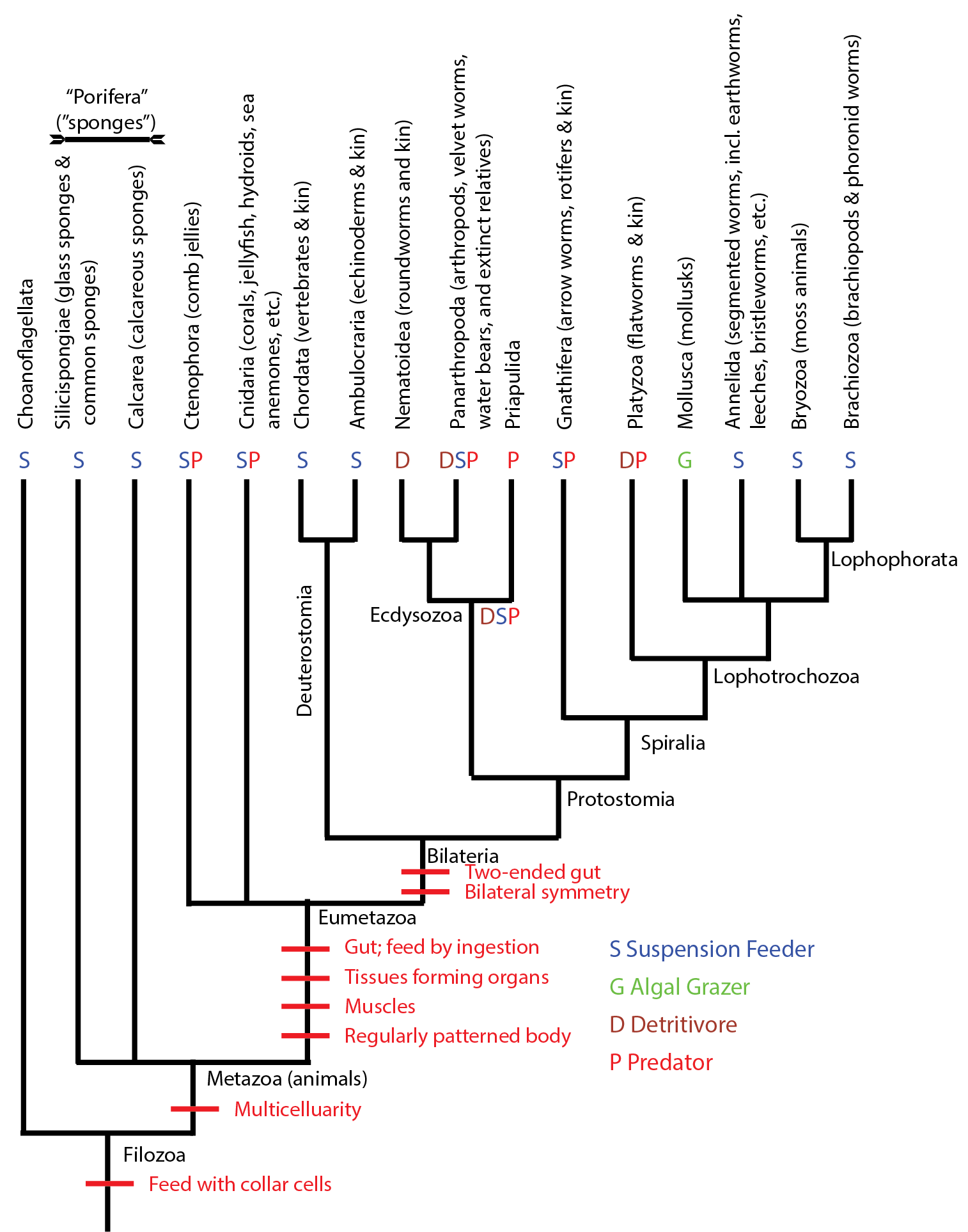
As we can see, suspension feeding is by far the most common mode of life for Metazoa. And the two groups of bilaterians which are clearly present by the end of the Ediacaran are NON-suspension feeders (predatory priapulids; algal-scraping mollusks). The great diversity of suspension-feeding "worms" were present may have been too tiny to be preserved, or lived in environments that are not preserved: neither case is particularly compelling, but that seems to be the situation. (It is true that there are many microscopic soft-bodied bilaterians: rotifers, tadigrades, chaetognaths, etc.)
But as we move into the Cambrian, the record changes.
While some of these organisms are known from hard parts, others (such as radiodonts and stem-vertebrates) were soft-bodied. These are known only from Lagerstätten. Thankfully these are fairly common in the Cambrian. Among these the first discovered and most influential in changing ideas about Cambrian diversity is the Burgess Shale of British Columbia, Canada (about 515 Ma). Subsequently earlier ones have been found: the Chengjiang fauna of the Maotianshan Shale, China (525-520 Ma); Sirius Passet in Greenland (527 Ma); and more.
The Cambrian Explosion Revisited: What is It?: So, with more data at hand, what is the Cambrian Explosion, really? Is it:
Let's take a look at these.
We can reject the literal phylogenetic model already. The Ediacaran evidence shows that crown-group Bilateria was already well diverged 10s of millions of years prior to the Explosion. Furthermore, molecular phylogenetic divergence dates point to Ediacaran (or even Cryogenian) branching events among the major divisions of Bilateria. So the lineages containing the members of the Explosion were present 10s of millions of years before they began to mineralize.
Ongoing genetic studies strongly suggest that key genetic complexes lead to increasing morphological and taxonomic diversity. But this is still ultimately a phylogenetic issue: the origin of these complexes must have occurred prior to the divergence between the different lineages, which we have already seen were Precambrian rather than Cambrian events. We can thus reject the developmental model as the cause of the Cambrian Explosion per se: however, the existence of these gene complexes were likely exapted during the Explosion to allow for rapid anatomical changes during that event.
We have already seen that the development and scale of biomineralization increases from the Ediacaran (millimeter-scale) to the earliest Cambrian (sub-cm-scale) to "Cambrian Explosion" proper (cm-scale), and across the Tree of (Metazoan) Life from sponges and eumetazoans (Ediacaran) to tiny elements in mollusks, brachiopods, sponges, etc. in the SSF to complete macroscopic skeletons. This is fully consistent with some form of change external to the organisms allowing first the very smallest shells, then armor made of individual tiny elements, and only later in complete large armor, all occurring across the phylogeny. Thus the geochemical model is supported, at least provisionally. But what might be changing? Nearly every mineral or element has been suggested by some researcher, but two in particular seem most likely:
Stem-Mollusks Support the Extrinsic Model: We actually see a pattern supporting this transition by looking at stem-mollusk history:
So that pattern is strongly consistent with a changing external situation allowing for increased levels of mineralization. But this leads to another question, namely…
Why Biomineralize at all?: We might naturally assume this was for defense (that is, shells as armor), and that might well be part of this. But there are other reasons for this:
Study of the Cambrian faunas show that they are far more diverse and complex than Ediacaran ones, with the presence of multiple levels of predation, pelagic communities (with nekton and plankton), and diverse infauna being major changes. (These faunas are not as complex as Ordovician or later ones, however).
These changes are consistent with the ecological model as a driver for the Cambrian Explosion. Indeed, these different models need not be mutually exclusive.
The Cambrian Substrate Revolution (aka the Agronomic Revolution): Increasing grazing and increasing burrowing doomed the algal mat community. (Indeed, thick biofilms only grow today in places where metazoans are excluded). While the sediment-water interface was once firm and the nutrients in the sediment once trapped and excluded from the water, the Cambrian Substrate Revolution churned up the sediment, allowing for soupier sediment-water interface and more availability of the sediment nutrients into the water and vice versa.
Putting it All Together: Rather than a single simple answer, it looks like there are a number of factors contributing to the Cambrian Explosion, extended over the Ediacaran into the early part of the Cambrian (and hence the "long fuse" of the Explosion):
To Syllabus.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}