


Key Points:
•One of the most useful ways of examining the large-scale patterns of life history in the oceans is Sepkoski's Evolutionary Faunas (EF): broad coalitions of taxa whose fates tend to match each other
•The history of the Ediacaran and Phanerozoic has seen a series of major changes, both originations and mass extinctions
"All these facts, consistent among themselves, and not opposed by any report, seem to me to prove the existence of a world previous to ours, destroyed by some kind of catastrophe. But what was this primitive earth? What was his nature that was not subject to man's dominion? And what revolution was able to wipe it out to the point of leaving no trace of it except some half-decomposed bones?" "Espèces d'éléphans" (1796), Georges Cuvier

Sepkoski's Evolutionary Faunas and Bocout/Sheehan's Ecologic Evolutionary Units, drawn by Ashley Dineen, from Roopnarine's Food Weblog
In 1981 Jack Sepkoski used factor analysis of Phanerozoic marine communities. He discovered that three major associations of taxa had similar evolutionary fate: they tended to increase together and tended to decrease together. On top of that (and almost certainly not coincidentally) they tended to have shared ecological properties. He named these the Evolutionary Faunas (or EFs), and gave each two names: one name refers to the time period in which they were the most common, and another by a key taxon of that community.
These evolutionary faunas are:
(It is worth noting that there were also some non-aligned taxa as well, that didn't statistically sort with the three EFs.)
Note that all three EFs are present in the Cambrian, and all are still present today (the Cambrian EF represented mostly by lingulates and hexactinellids, neither of which are speciose but both still present). The names refer to their heydays, not their limits.
The EFs divide up biological diversity in the seas. An alternative was to break up the macroecological history was initially developed by A.J. Boucot in 1983 and modified into their modern form by P. Sheehan in 1991. These are the Ecologic Evolutionary Units (EEUs): divisions of time characterized by suites of taxa, bounded by extinctions or replacement. A current look at these gives the following succession:
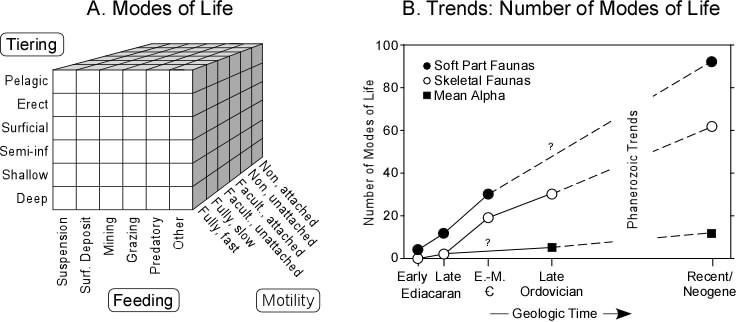
The ecospace model and trends in ecospace filling, from Bambach et al. (2007) doi: 10.1111/j.1475-4983.2006.00611.x
R.K. Bambach, A.M. Bush, and D.H. Erwin developed a useful scheme for describing the complexity of ancient ecosystems. They suggested a 3D ecospace, with the axes based on feeding style, tiering relative to sediment, and motility. Any taxon can be placed in one cube within this larger cube; even more, different communities could be compared by the amount of ecospace filling.
Their work supports the idea that Ediacaran communities really had less ecospace filling than the Phanerozoic; that as the Cambrian and Ordovician moved along the amount of ecospace filling increased; and that Mesozoic and Cenozoic ecosystems really are more "full" than what we know of Paleozoic ones.
Work by P. Wagner and colleagues independently suggests this. They found that the total diversity of life post-Paleozoic was much higher than the standing Paleozoic levels. Comparisons of the communities suggests that any given Paleozoic community had fewer but more common species, while post-Paleozoic ones tended to have a few common but many more rare species.
Cuvier demonstrated the existence of mass extinctions (or "revolutions" as he called them): the loss of great parts of the diversity of life, to be replaced by other forms. Furthermore, Cuvier assumed that these revolutions were the products of catastrophes of some sort.
Regardless of their cause, geologist John Phillips (in 1841) used the pattern of fossil diversity, and especially the crash of diversity to define the Eras of the Phanerozoic: thus the boundaries between the Paleozoic and Mesozoic Eras and between the Mesozoic and Cenozoic Eras were mass extinctions.
But not everyone was convinced that mass extinctions were real phenomena. 19th Century geologist Charles Lyell and Charles Darwin thought that there were no mass extinctions: rather, what we thought were Cuvier's "revolutions" were no more than extended gaps in the rock record, making the gradual loss and addition of species at the ordinary rate appear to be the sudden loss and appearance. In Darwin's words (from The Origin):
In the early 20th Century at least some paleontologists resurrected Cuvier's ideas of 'revolutions' under the name of global diastrophism ("diastrophism" being an old world for "geological uplift, mountain building"). Leading scientists such as the American Museum's H.F. Osborn thought that periods of mountain building occurred worldwide, resulting in major climate shifts and causing intense droughts, climate changes, etc.
As it turns out, both the Lyell/Darwin and Osborn ideas were testable by the same observation. Both required that the extinction boundaries would occur at unconformities (erosional surfaces): in the former case, because all the missing record was lost; in the latter, because global diastrophism would cause uplift and erosion. But when many extinction sites were sampled, no such unconformities were found. Something else must be going on.
In the mid-20th Century paleontologist Norman Newell finally coined the term "mass extinction" and recognized these as "crises in the history of life". Because of advances in understanding geologic time, it was recognized that Lyell and Darwin's "gap" argument did not work: there simply wasn't enough missing time to turn these boundaries into "simply business as usual, with missing rock". As for causes, Newell (who worked primarily on marine invertebrates) argued they were mostly due to rapid sea level changes and/or changes in atmospheric composition. Around the same time, German paleontologist Otto Schindewolf argued that some mass extinctions might have been caused by cosmic causes (such as comets colliding with Earth.)
In the later 20th Century paleontologist Jack Sepkoski finally gave us a good working definition for mass extinctions:
Agents of Destruction
When talking about "causes" of mass extinction, we have to recognize we are talking about several different things:
When identifying reasonable causal agents for any given mass extinction, we have to evaluate if that agent:
Some paleontologists have described a distinction between pulse and press extinctions. Pulse extinctions are rapid catastrophic events, which do not allow for adaptive changes during the episode itself. These are the "Game of Thrones" events: you either win or you die. In contrast, press extinctions would be series of events spread out for 100s of thousands of years. In principle, it allows for evolution to select variations throughout this extended interval, so a taxon which is doing poorly early in the event might become progressively better at surviving it over time. (That said, no one has made a good case for any pulse aspect to mass extinctions.)
A related set of terms are proximate vs. ultimate causes. Proximate causes are the events that directly brings about the change. Ultimate causes are the phenomena that sparks the proximate causes. For example, the proximate cause for the sinking of the Titanic was the flooding of the decks through the hole ripped in its side by the iceberg; the ultimate causes were (among other things) failure to spot the iceberg sufficiently in advance to turn it. For any given extinction (or mass extinction), we have varying levels of success in identifying the proximate or ultimate cause.
A problem with all mass extinction studies is the issue of stratigraphic resolution: that is, how closely does our observed record match the actual event itself. The same set of observed data of species disappearing one-by-one running up to a stratigraphic boundary might potentially be explained as a gradual extinction OR it might be explained by the fact that we are statistically unlikely to recover the last population of any given species. This is the Signor-Lipps Effect: the observation that the stochastic nature of fossil preservation and recovery will tend to "smear" out an instantaneous extinction to make it look gradual.
In order to counteract the Signor-Lipps effect we need to have very high sample sizes of fossils and of stratigraphic horizons running up to the possible event. But nature doesn't always provide us with this, so we always have to remember that our ability to definitively say when a particular taxon went extinct is limited.
The Big Five
The 1970s and 1980s saw renewed interest in mass extinction studies, in response to research of the K/Pg event 66 Ma (the one that ended the giant dinosaurs): first, the Russell and Tucker supernova hypothesis, and then the Alvarez et al. discovery of the iridium spike (more about these specifics in a few lectures). While this was going on, invertebrate paleontologist Jack Sepkoski was assembling a huge data base of the diversity of marine life through time. This data (like Phillip's in the 1840s, and Newell's in the mid-20th Century) helped reveal the presence of mass extinctions. In this case, however, Sepkoski worked with fellow paleontologist David Raup in 1982 to reveal statistically a difference between "background extinctions" and "mass extinctions".
In this study, Raup and Sepkoski idenfitied what came to be called "The Big Five": mass extinctions which were statistically quite different from the ordinary level of diversity collapses. Raup and Sepkoski identified the Big Five mass extinctions: the Ordovician/Silurian (O/S, 443.8 Ma), the two-phase Late Devonian Mass Extinctions (372.2 and the D/C boundary 358.9 Ma), the Permian/Triassic (P/Tr, 252.17 Ma), the Triassic/Jurassic (Tr/J, 201.3 Ma), and the Cretaceous/Paleogene (K/Pg, 66.0 Ma).
Various studies may lose some of these mass extinctions as barely larger than background (especially the D/C), and some mass extinctions may represent multiple events (dividing up P/Tr into two; turning the D/C into two; etc.) the count maybe be more than 5. But we'll stick with the Big Five for this course. (This doesn't meant that there aren't minor mass extinctions: some moderate mass extinctions do seem to exist.)
General Patterns of Mass Extinctions
In general it is a truism that mass extinctions would occur when the rate of extinction greatly exceeds the rate of origination of new species. But are extinction and origination events independent in time, space, and cause? And are these happening gradually throughout the history of a lineage? After all, evidence points towards a general pattern being turnover-pulses: both extinctions and originations/divergences driven at the same time by environmental shifts.
Mass extinction is thus a "Game of Thrones": you only survive or you die. It happens when environmental change happens with such severity that organisms cannot adapt to the shifting conditions. The resultant pattern of life history is long periods of relative stability in terms of groups represented, punctuated by mass extinctions and resorting of the players of the game.
It increasingly looks like most of the major mass extinctions have at least two strong environmental factors, often working in opposite directions. For instance, many have powerful deep freezes accompanying major global warming episodes. These "one-two punches" may be necessary to truly destabilize global ecosystems.
There are some recurring patterns after many mass extinctions. These include:
Cambrian Substrate Revolution (throughout Cambrian): Also called the Agronomic Revolution, it saw the rise of grazers and shallow infauna disrupted the matgrounds that dominated Precambrian marine biomes. Loss of the matgrounds resulted in the extinction of taxa that relied on these surfaces as a habitat, and a migration away from them by same taxa that once lived them (for instance, polyplacophorans moved from soft muddy/sandy surfaces to rocky surfaces.) Additionally, the lack of the matgrounds allows for easier move of biogeochemicals from the sediment into the water column, where they were once more isolated.
The Great Ordovician Biodiversification Event (GOBE) (throughout Ordovician): During the Ordovician the Cambrian EF remains high, but also saw the vast increase in the amount of the Paleozoic EF taxa. This saw the migration of animals away from the sediment-water interface, and the rise of far more predation.
A side effect of this was the migration of some of the EF into deeper water from the shallows.
Ordovician/Silurian (443.8 Ma) Mass Extinction: The Ordovician Period saw a dramatic increase in biodiversity, but crashes at the end. Perhaps 85% of species are lost (although the total number is less than later ones: there were fewer total species known for the Ordovician than for later in the Phanerozoic.) There is devastation within brachiopods, bryozoans, graptolites, trilobites, and conodonts, but more major clades were lost. A fair amount of evidence suggests there were two pulses of extinction, separated by about 1 Myr.
Despite some news items in the 2000s suggesting a "hypernova" or "gamma ray burster" (extremely huge supernova) explosion being the cause, there is zero (zip, nada, no) evidence for this. Instead, this event coincides with a major period of glaciation. It has been suggested that this might have been responsible for the extinction by perhaps reducing the available habitat area in the shallow marine realm (because of a major sea-level drop), and/or a major period of anoxia from global eutrophication, due to the influx of considerable amount of nutrients from material scraped off the continents from glaciers.
The Devonian Nekton Revolution (throughout the Devonian): During the Devonian there is a great increase in the amount of fully nektonic forms, especially fish and ammonoids. Previously the nektonic taxa were mostly demersal: that is, they swam, but mostly lived and fed right above the sediment-water interface. But from the Devonian onward animals began to occupy more and more of the water column, all they way up to the water-air interface.
A side effect of the increase of nekton was the greater transfer of fecal material and carcasses into the deep sea, as swimmers a) move up and down in the water column during diurnal migration, and b) they live farther and farther from shore.
The Greening of Land (Silurian and Devonian): The rise and spread of land plants (especially seed plants) binds up more sediment, allowing for more meandering streams. Additionally, the first paleosols and coals begin to sequester more carbon from the atmosphere. These cause a rise in the amount of atmospheric oxygen (more of that to come) and a cooling of the world (due to reduced greenhouse warming.)
Late Devonian Mass Extinctions (372.2 & 358.9 Ma): A two-phase event: a first strong one between the Frasnian and Famennian Stages of the Late Devonian Epoch (also called the "Kellwasser Event"), and the final Devonian/Carboniferous event (also called the "Hangenberg Event"). Some studies once supported a single major terminal Devonian event, with the Frasnian/Famennian event as a result of the Signor-Lipps effect, but more recent work shows the double phase. This is characterized by a collapse of major reef community (the tabulate-stromatoporoid reef community); as today, reefs in the mid-Paleozoic were MAJOR centers of biodiversity, so collapse of the reef community dragged numerous other species with it. Additionally, there are losses of primitive grades and clades of fish ("ostracoderms", "placoderms", and acanthoidians). There are also major extinctions within trilobites (again), eurypterids, echinoderms (with some major clade losses), conodonts (again), bryozoans (again), and ammonoids (for the first but not the last time). Many of these survived with only a few handful of species to repopulate the Carboniferous.
Although Schindewolf proposed an asteroid impact as the cause for this (without any direct evidence), this is currently best explained as a result of the spread of vascular land plants! With the first rain forest and the first seed plants in the continental interior, there was considerable burial of excess carbon (leading to cooler temperatures and reduced continental shelf space). More importantly, increased mechanical and chemical weathering on the continents leads to increased nutrients into the sea leading to marine phytoplankton booms leading to eutrophication and anoxia. (Indeed, modern coral reefs are suffering from these effects locally because of increased runoff due to human activities in these regions.)
Permo-Triassic (252 Ma) Mass Extinction:A The "Mother of All Mass Extinctions" (so named by Doug Erwin of the Smithsonian), this is the greatest diversity crisis known. If this was the single terminal Permian event, then it was an event with 55.7-82% of the marine genera went extinct (which corresponds to an 80-96% species level extinction). Or, to put it another way, there was only 4-12% survivorship at a species level, and given that a species could survive with very few individuals, it was much greater than 96% of individuals lost!). In comparison, the K/Pg had a 40-47% genus loss.
However, some models suggest that this is a two-phase extinction, with an earlier one between the Middle and Late Permian (the Guadalupian extinction), then EACH of these is among the greatest extinction events. As with the K/Pg there are many names for this event, including the Changhsingian/Induan extinction, or (using older terminology) the Tartarian/Scythian extinction.
This was a "game changing" event. Most estimates show that diversity of marine organisms was essentially stable from the Late Ordovician until the end of the Permian (minus the crash-and-recovery intervals at each mass extinction). In contrast, life ever since the Permian has been relatively steadily increasing in diversity, surpassing Paleozoic levels sometime in the Cretaceous.
Victims that die out entirely include tabulate and rugosan corals, trilobites, eurypterids, graptolites, many types of echinoderms, primitive plant groups, and primitive insect groups. Bryozoans, rhynchonelliforms, crinoids, ammonoids, crinoids, and vertebrates all suffer major collapses but recover to one degree or another.
The ultimate cause appears to be the Siberian Traps, a huge lava field in Siberia with an area of 5 million km2, and a volume of about 3 million km3! (This would cover North America to a depth of 121 m (nearly 400'!). It erupted over the space of less than 1 Myr: a tremendous volume in a geologically-short interval. The Siberian Traps are the greatest example of a flood basalt: a form of eruption where vast volumes of lava (along with dissolved gases) spread out over huge regions in burst after burst. They are also called "large igneous provinces". The don't represent eruptions from a single conical volcano: rather, a huge region would have fissure eruptions.
However, although the Siberian Traps are the causal agent, the killing agent isn't the lava (except for the unlikely animals and plants of latest Permian Siberia!). Instead, the killing agents are produced mostly from the gases and aerosols released by the eruptions, and the side effects of these.
First and foremost is the carbon dioxide release, producing some of the most extreme global warming in Earth's history. Today (2018) there is about 873 Gt (gigatons) of C (carbon) in our atmosphere, reflecting about 410 parts per million CO2 (1 ppm = 2.13 Gt C). Before the Industrial Revolution, that value was around 600 Gt C. The Siberian Traps dwarfed these values by several orders of magnitude. Initial estimates were that over the course of the eruptions about 12,000-18,000 Gt C were released; newer modeling points to values closer to a mind-staggering 170,000 Gt C!! This didn't happen as a single burst, so there was no moment when Earth's atmosphere had 80,000 ppm CO2 during this interval, but it was still tremendously higher than today (or, more important for this issue, compared to the time before the extinction. Values aren't certain, but carbon dioxide levels had an increase of at least 8 times, and possibly more! This lead to extreme global warming (surface temperatures rising by more than 7°C, possibly much more: the latest models suggest air temperatures averaging 45°C (113°F)!!), which lead to warming of the sea floor, which lead to melting of the methane clathrates (methane frozen in ice on the sea floor), which bubbled into the atmosphere, which led to even more global warming. The oceans would also become more acidic because of the carbon dioxide mixing with water, causing damage to shell-forming organisms.
Additionally, the new studies point to as much as 18,000 Gt HCl (that's hydrochloric acid!) being released by the melting of underground rock salt deposits. Between the HCl, sulfuric acid (a byproduct of sulfur dioxide, a common component of eruptions), and carbon dioxide there would likely have been catastrophic levels of acid rain and oceanic acidification, worldwide. On top of that, atmospheric sulfate aerosols would generate 10-15°C cold pulses!
Additionally, there were tremendous drops in atmospheric and oceanic O2 due to the mass death of so many land plants and phytoplankton and to oxidation of the methane. To make matters worse, increases in global temperature would decrease the temperature differential around the world, decreasing oceanic and atmospheric circulation, and thus reducing the churning up and oxygenation of the ocean water.
A consequence of the ocean anoxia is the promotion of sulfur bacteria. These prokaryotes prefer anoxic and dysoxic conditions. Their photosynthesis does not release oxygen as a waste product; instead, they release hydrogen sulfide (H2S), a gas highly poisonous to both aquatic and land animals. With shallow sunlit water devoid of their oxygen-generating competitors, the purple bacteria spewed out hydrogen sulfide, further making the shallow seas and land a hellish condition for other life.
But wait, there's more! The sulfates ejected by the eruptions and the H2S from the bacteria can destroy ozone. As these gases made their way into the higher atmosphere, the Earth lost its protection from dangerous UV radiation reaching the surface. Malformed pollen had already been known from latest Permian rocks; a recent experimental study showed that these malformations can be produced by subjecting plants of the same general groups present in the Permian with high levels of UV. Such malformations would greatly decrease plant fertility. (On top of that, the UV can kill the plants themselves, as well as animals.)
Stratigraphic evidence demonstrates that the main pulse of extinction took a mere 60 kyr, plus or mins 48 kyr, to occur!
And keep in mind, these killing agents are produced independent of living things. The eruptions would continue for 100s of kyrs, so that there would be long slow recovery for the atmosphere, ocean, and pedosphere (soil). This meant that the ecosystems continued to suffer for millions of years.
Triassic/Jurassic (201.3 Ma) Mass Extinction: In some ways a repeat of the P/Tr on a smaller scale. The final extinction of the conodonts, a major diversity crash in ammonoids (AGAIN!), loss of once-major groups of brachiopods and bryozoans, collapse of some corals and sponges, and on land loss of primitive seed plant groups, various diapsid reptiles (especially the majority of the pseudosuchians outside of Crocodylomorpha), and of various therapsids (basically everything but mammals and their immediate closest kin).
The causal agent here is the Central Atlantic Magmatic Province (CAMP for short), a major field of volcanics and intrusive igneous rocks associated with the break-up of Pangaea and the formation of the Central Atlantic Ocean basin. It is comparable in scale to the Siberian Traps of the P/Tr in terms of the area covered and volume erupted, but apparently lower levels of carbon released: "only" 2200-2500 Gt C as CO2 and 4300 Gt C as CH4. Once again, kyr-long cold snaps seem to have punctuated the warming pulses.
Reddening of the Seas (mid-Jurassic-Cretaceous): The shift in the marine realm from "green" chlorophyll algae groups to "red" chlorophyll ones (coccolithophorids, dinoflagellates, diatoms) changes the amount of nutrients in the seas. The C:P ratio shifts, making the plankton more nutritious. This increasing food quality and quantity drives richer and more complex marine food webs.
The Mid-Mesozoic Marine Revolution (mid-Jurassic-Cretaceous: Simultaneous changes happen in the benthos. In the mid-Mesozoic shift from communities dominated by sessile attached animals to ones of modern style, dominated by moving mollusks, crustaceans, fish, asterozoans, echinozoans, etc. Deeper infauna become common. Brachiopods and sessile crinoids become displaced into deeper water or crevices. The remaining epifauna often become more spinose.
This seems to be ecologically-driven by the rise or expansion of various predatory groups: crushing crustaceans, drilling gastropods, asteroids, teleost fish, etc.
The Cretaceous Terrestrial Revolution (mid-Cretaceous): Rise and diversification of more modern mammals, advanced insects, the newly-evolved angiosperm plants, and other small-bodied organisms during middle of Cretaceous. The KTR results in the modern world condition, where most of the biomass and 85-95% of macroscopid species are terrestrial (although a lot of these species are limited to the tropics and live at low population density.)
Cretaceous/Paleogene (66.0 Ma) Mass Extinction: Last and most famous, but not biggest, of the "Big 5". Formerly the Cretaceous/Tertiary (K/T) extinction. 50% or higher genus extinction rate in the marine communities; on land, essentially all animals >5 kg died out. The end of the ammonoids, inoceramids, and rudists; large marine predators (sharks, actinopterygians, marine reptiles); and major crashes in coccolithophorids (which never recover their old diversity) and forams (which do).
Another major flood basalt (the Deccan Traps of western India) does begin within 350-250 kyr before the extinction, and likely contributed to it with the same environmental effects (on a smaller scale) as the P/Tr and Tr/J. But the big deal is the asteroid impact in the Yucatán Peninsula in Mexico (named Chicxulub after a town where a drilling core first documented the disrupted boundary layer.)
The impact was initially discovered by an increase in the level of iridium at the K/Pg boundary in Gubbio, Italy. The 180 km diameter crater, now buried, in Mexico indicates a 1.8 x 108 megaton blast: the equivalent to 1000 to 10,000 times the full nuclear arsenal at the height of the Cold War (or a century's worth of geothermal output). This blasted out a vast amount of material, some of which rained down planetwide on the same day, and others of which stayed high in the stratosphere for years.
The infalling debris burned up in reentry, generating a thermal pulse 8-10 times the heat at the equator at noon everywhere around the world. This pulse lasted for minutes to hours, cooking all living things exposed on the surface. (Aquatic animals would be protected, as would burrowers.) Some of the infalling debris may have ignited forests around the world: there is the suggestion of a global forest collapse.
More significantly the stratospheric dust resulted in an impact winter: recent models show temperature drops of 26C° for 3-16 years, and more than 30 years before the dust clears. And because the impact site was covered by carbonate rock, the impact resulted in a huge increase in atmospheric carbon dioxide. When the air cleared up, a greenhouse summer raised surface temperatures by 5°C for 100kyr.
To Syllabus.

{kind=link}
{kind=link}
{kind=link}