Key Points:
•Macroevolution is term for evolutionary changes recognized above the species level.
•One major macroevolutionary pattern testable in the fossil record is whether species-level change is mostly gradual (phyletic gradualism) or in short bursts separated by long period of stasis (punctuated equilibrium).
•Under phyletic gradualism, then species don't have real beginnings or ends, and are just "chronospecies" with anagenetic origins; under the latter, the case can be made that species are "real" rather than purely arbitrary.
•In recent decade it has been recognized that evolution is highly controlled by development: this is represented by the new study of "evo-devo".
"Why then is not every geological formation and every stratum full of such intermediate links? Geology assuredly does not reveal any such finely-graduated organic chain; and this, perhaps, is the most obvious and serious objection which can be urged against the theory." -- Chapter 9 "On the Imperfection of the Geological Record", On the Origin of Species by Means of Natural Selection (1859), Charles Darwin
"Since we proposed punctuated equilibria to explain trends, it is infuriating to be quoted again and again by creationists -- whether through design or stupidity, I do not know -- as admitting that the fossil record includes no transitional forms. The punctuations occur at the level of species; directional trends (on the staircase model) are rife at the higher level of transitions within major groups." --The Panda's Thumb (1980), Stephen Jay Gould
"A plausible argument could be made that evolution is the control of development by ecology. Oddly, neither area has figured importantly in evolutionary theory since Darwin, who contributed much to each." --"Festschrift" (1973), Leigh Van Valen
Macroevolutionary Patterns
With the discovery of evolution by natural selection, biologists from Darwin and Wallace's time onward have documented
many different patterns and processes in evolution. Sometimes they refer to "microevolution" (changes within an species) and
"macroevolution" (patterns on the larger scale; changes from one species to another, or between different lineages of
ancestors and descendants). It is important to remember that "micro-" vs "macro-" is just a matter of scale and perception:
at the level of individuals and populations, there is just variability, heritability, and superfecundity.
Two (or more) distinct variations in an ancestral population convey their own
advantage against the rest of the population
Over time, these two (or more) variations will become more distinct from each other
If they diverge enough, they will no longer be able to mate with each other: will be
different species
Divergence can also occur (perhaps more commonly!) if an ancestral population is divided
into two or more by changes in geography: because natural selection works by chance
survivals, it is unlikely that exactly the same variations of the ancestral population
will survival in the two or more separated populations. Over time, if the populations
meet again, the accumulation of variations may be significant enough that they are
distinct species.
Common Ancestry
Closely related species are close because their common ancestor diverged relatively recently
in Earth history
Other species are more distantly related because of divergences of THEIR common ancestors
even farther back in time
No separate origins for different groups; instead, patterns of common ancestry and diverging
descendants
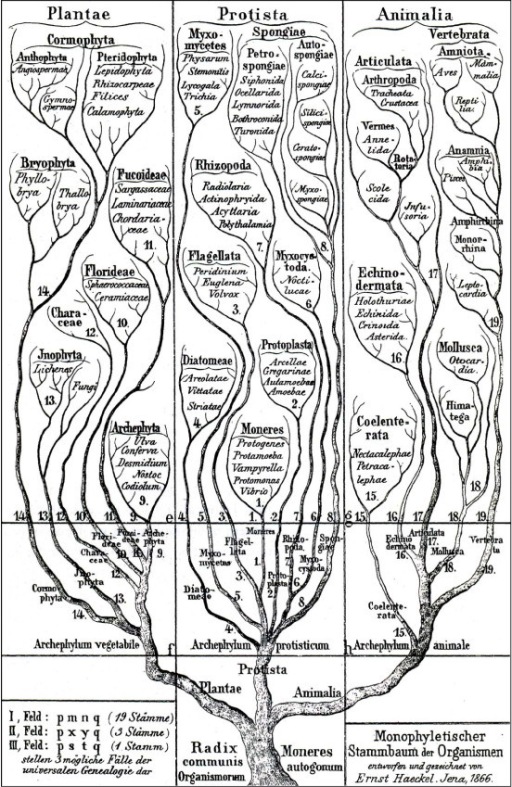
Thus, the basic pattern of the history of living things is a Tree of Life,
where the trunk and stems are lineages of ancestors, the branching points representing divergences between lineages, and
the tips of the branches living species (or extinct species that died without descendants).
Other important patterns and processes:
Sexual Selection, a variation of Natural Selection recognized by Darwin, where the
variation is "being more sexy" (and thus have better than average chance of breeding,
and thus passing on "sexiness", compared to other members of the population [increased reproductive success]).
Explains many extravagant display structures and behaviors (such as peacock tails, bird song, lion manes, etc.)
Correlated Progression: Ancestor and descendants form a lineage (historical line). Sometimes a particular
life habit favors the slight increase in multiple different traits (e.g., longer and longer legs, more compact body, more
efficient heart and respiration for fast running; longer and longer necks, longer legs, better cropping teeth and/or grasping
tongue for browsing in trees; more and more streamlined body profile, more paddle like legs, more dorsal nostrils, etc. in swimmers; etc.)
Traits that go against the general trend will be selected against; traits that go with the general trend will be selected for.
(For many people, this series of trends in adaptations represents the totality of evolution)
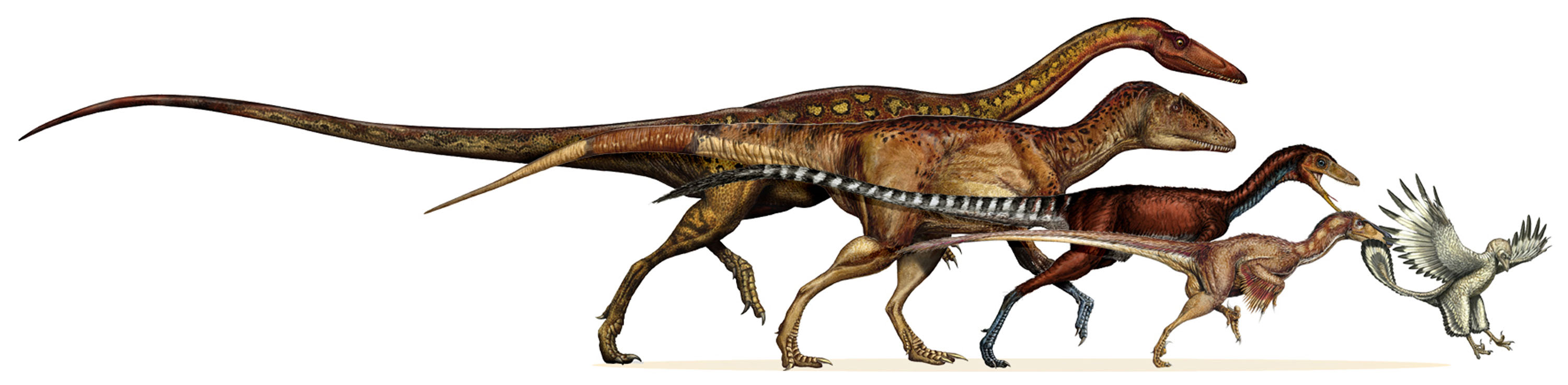
Adaptive Radiation: If a population evolves some significant new adaptation, or colonizes a region without competitors,
or is present when competitors die off, many different variations from that common ancestral population might survive
(fill new or unoccupied "niches" (ways of life) in environment). Over a geologically short period time, a common ancestor can
radiate into many very different descendant lineages.
Niche Partitioning: during an adaptive radiation, the early members of the divergence will (naturally) still be
relatively similar to each other (and to their common ancestor) in terms of size, shape, behavior, etc. Over time, those
variations in each lineage that are least like their relatives will more likely survive, because they will have less
competition. Consequently, the different species will "partition" (divide up) the niches and the resources.
Convergence: Some adaptations are mechanically advantageous and easy to produce developmentally. Different
lineages of organisms can independently develop some of the same features, even though ancestors were quite different
(i.e., streamlining in sharks, tunas, ichthyosaurs & dolphins).
Living Fossils: Species do not have a fixed duration, but will persist until the evolve into something else
and/or go extinct. In some cases, species (or genera) may persist for extremely long periods of time with no major
changes.
Exaptation: Formerly called "preadaptation", the co-option of a structure that previously had some
entirely different function for a new use. Seems to be the more common pattern of evolution than the appearance of
entirely novel structures. For example, the wings of birds and bats were initially arms and hands; the mouthparts of
various arthropods were legs; etc. Developed as part of Punctuated Equilibrium.
Heterochrony: Evolution by changes in rate of development from embryo to adulthood (as discussed in a previous lecture)
History of Evolutionary Models
Non-natural selection evolutionary models in the early 1900s:
Orthogenesis/Aristogenesis
Commonly supported by paleontologists
Predicted anagensis as dominant pattern of speciation
Found great "support" in Kelvin's abbreviated geologic time scale
Saltationism
Supported by many geneticists
Some plants seem to follow this model, but unusual genetically
Following these early attempted modifications of Darwinism, the rest of the 20th Century onward stayed largely within a Darwinian model. However,
there were different major schools of thought. Many of these differences hinged on views of microevolution (evolutionary change within a
species) and macroevolution (evolutionary change above the species level). While most agreed that the ultimate processes in macroevolution
were ultimately microevolutionary, there were disagreement whether the patterns produced were actually reducible to microevolutionary changes.
First major school: New Synthesis (also called Modern Synthesis and Neo-Darwinism): product of collaboration of work by geneticists, statisticians, and traditional
organismal biologists (including paleontologists) (but notably NOT embryologists!!)
Developed in 1940s; dominated evolutionary biology from the 1950s onward. A major 1970s contribution: "Selfish Gene" or "Gene’s Eye View" models
Source of the statement "Natural Selection is the Change in Gene Frequency Over Time"
Panselectionist: tended to emphasize selection for all different aspects of organisms phenotype (rather than considering them to be product
of contigency)
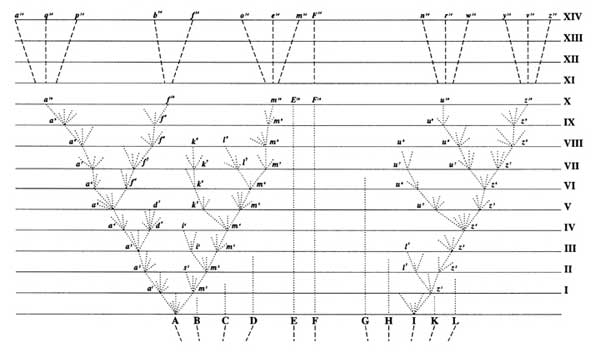
Promoted phyletic gradualism
Anagensis dominates over cladogenesis
Morphological change distributed throughout lineage's history
Parodied as "evolution by creeps"
However, also promoted the allopatric/peripatric speciation model
Primarily paleontologically-generated challenge to New Synthesis: Punctuated Equilibrium:
Speciation primarily by cladogenesis
Morphological change limited to short speciation events (punctuations) representing only
tiny fraction (10% or less) of duration of speciation
Long periods of morphological stasis in between
Rather than the gradualist model (which assumed that there must be tremendous gaps in the fossil record to explain lack of changes
within "chronospecies"), punctuated equilibrium considered stasis to be data.
Often falsely characterized as a "saltationist" model, but in reality rate of change during
puncuations are on par with field and lab observations (weird aspect is stasis, not punctuation)
Parodied as "evolution by jerks"
Species Selection: Not a separate phenomenon/process from Darwinian Natural Selection, but a pattern expressed at a higher level
Species as "individuals" with origins (speciations) and deaths (extinctions)
Two clades (esp. sister taxa) compared in terms of species selection by differential patterns of origins & extinctions, just as two
subpopulations might fare differently over time under Natural Selection proper
Emphasis on emergent properties: To the Modern Synthesists, all macroevolution is reducible to microevolution (i.e., it has no
properties not also exhibited at the lower level. To PE, macroevolution has emergent properties:
Even though the ultimate phenomena is differential selection of variants in a population, patterns emerge at higher levels that are not
directly predictable/observable at lower level
Analogies in other sciences: Behavioral patterns (e.g., migration, mating displays, etc.) of vertebrates are not directly observable
from individual cells; Metamorphic grade and fabric not directly predictable from atomic physics
Emphasis on contingency: "The tendency of complex systems with substantial stochastic components and intricate nonlinear
interactions among components to be unpredictable in principle from full knowledge of antecedent conditions, but fully explainable after
time's actual unfolding" - S.J. Gould (2002, Structure of Evolutionary Theory)
Related concept: phylogenetic constraint/inertia: Limitations on future evolutionary trajectories imposed by previous adaptation
What are the Relative Effects of Biotic vs. Abiotic Factors in Macroevolution?
Red Queen Hypothesis:
Vertebrate paleontologist Leigh Van Valen observed constant probability of extinction at "family" level over time: i.e., no sign that
lineages had improved survival chance no matter how long they had existed
Ascribed this to co-evolution of predators, competitors, prey/food items
These ecological interactors are themselves evolving adaptations in response to the lineage in question
Likened to the "Red Queen" of Wonderland, who had to run to stay in place
Court Jester Hypotheses:
Environmental changes rather than interspecific interactions dominate evolutionary pattern
Developed in opposition to Van Valen’s Red Queen Hypothesis
Includes several related ideas: Stenseth & Maynard Smith's 1984 "Stationary Model"; Vrba's 1985 "Turnover-Pulse"; Brett & Baird's 1995
"Coordinated Stasis"
Periods of evolutionary change (i.e., punctuations and extinctions) occur at times of climate shift
Hence, would be expected to occur at same times across taxa in same enviroment
Are the New Synthesis and Punc Eq Really That Different? Evolutionary Stable Strategies (ESS): Combination of game theory and behavioral ecology. An ESS is a strategy which, if adopted by a
population of players, cannot be invaded by any alternative strategy. A Nash equilibrium which is "evolutionarily" stable meaning that once
it is fixed in a population, natural selection alone is sufficient to prevent alternative (mutant) strategies from successfully invading:
"A population is said to be in an evolutionarily stable state if its genetic composition is restored by selection after a disturbance,
provided the disturbance is not too large. Such a population can be genetically monomorphic or polymorphic." --Maynard Smith (1982).
Although ESS proponents (many of them behavioral ecologically-oriented neontologists) and Punc Eq proponents (mostly paleontologists)
often argued (especially in the 1970s-1990s), ESS provides a likely explanation for stasis: during punctuation events, the environmental
changes are so extreme that one or more mutants actually does do better than the previous ecologically stable state
Evo-Devo and the Source of Evolutionary Novelties
Since the mid-1990s, emphasis on the interrelationships between development (as studied in embryology), genetics, and whole organism biology
(esp. paleontology as record of Life's changes). Name given to this field (at conference at UMCP!): Evo-Devo (evolution & development).
Following Wikipedia's definition (!), it compares the developmental processes of different organisms in an attempt to determine the
ancestral relationship between organisms and how developmental processes evolved. Evo-devo addresses:
The origin and evolution of embryonic development
How modifications of development and developmental processes lead to the production of novel features
The role of developmental plasticity in evolution
How ecology impacts in development and evolutionary change
The developmental basis of homoplasy and homology.
Evo-devo reflects the discovery that there are developmenetal genetic toolkits (such as the HOX genes of animals) that control the timing, sequence,
rate, and duration of embryological changes. Modification of these genes (first seen in homeotic mutants) can produce both minor and major
morphological variations that can be acted on by natural selection.
Evo-devo also reveals deep homologies: while some organs may be the product of convergent evolution in different lineages (classic
example is the eye) and are thus analogous structures, the tissues from which the eyes are made, and the developmental genes that control this,
are often homologous at a much more ancient level.



{kind=link}
{kind=link}