


Key Points:
•Tetrapoda represents the clade of terrestrial four-legged vertebrates (including secondarily aquatic forms)
•Extant Tetrapoda includes Lissamphibia (the living amphibians), Mammalia, and Sauropsida ("reptiles", including birds). Mammals and sauropsids are united in the clade Amniota. Among other traits, tetrapods are united among living vertebrates by segmentation of the vertebral column into cervical (neck), dorsal (back), sacral (hip), and caudal (tail) regions, and by well-developed pectoral and pelvic girdles with well-developed fore- and hindlimbs (respectively) terminating in digits (fingers and toes.)
•"Amphibians" (broadly defined) represent those stem-tetrapods, stem-lissamphbians, lissamphibians, and stem-amniotes that retained the ancestral habit of an aquatic larval stage. Amphibian-grade vertebrates first appear in the Devonian. Many were ecologically and physiologically quite different from lissamphbians: for instance, many retained fish-style scales and used internal gills rather than lungs to respire carbon dioxide.
•The development of the amniotic egg (and associated anatomical transformation) allowed vertebrates to leave the water for their entire life-cycle. This transformation occurred during the Carboniferous Period.
•Synapsida (the mammal total-group) was ecologically dominant in the Late Carboniferous and Permian Period. Transformations including differentiation of the teeth and improvement of the limbs document the transition from the basal "pelycosaur" grade to the Therapsida.
•Therapsida remained present in the Triassic, and within this clade mammals arose. Mammals remained small but ecologically diverse during the Mesozoic; ecological release with the extinction of large-bodied dinosaurs at the end of the Cretaceous allowed for a mammalian radiation during the Cenozoic.
•Sauropsida includes several diverse lineages. Four of the most diverse clades are Pan-Testudines (total-group turtles), Euryapsida (an extinct clade of Mesozoic marine reptiles), Lepidosauromorpha (lizards (including snakes), tuataras, and their extinct kin), and Archosauromorpha (crocodilians, birds, and their extinct kin)
•Among other major ecological trends, tetrapods document: the widespread presence of herbivory; evolution of tremendous size in multiple clades; diverse forms of terrestrial locomotion; widespread presence of gliding, and the convergent evolution of powered flight three times; many cases of the evolution of secondarily aquatic habits; the highest intelligence of any clade; and multiple instances of highly elevated metabolism ("warm-bloodedness").
I come from the water
I crawled upon the shore
I left my brothers there
I got what I came for" -- Todd Lewis, lyrics for "I Come From the Water" (1994) by The Toadies
Last lecture, we saw the diversity of marine vertebrates ("fish"), ending with the transition from fully aquatic forms to taxa which could at least on occasion come up on land. In this lecture, we will examine what happened to the diversity of terrestrial vertebrates.
FOUR ON THE FLOOR: EXTANT TETRAPODA
The living four-limbed vertebrates form the clade Tetrapoda ("four-footed ones"). Among a great number of other traits, this clade is distinguished from the other living vertebrates by:
The living clades of Tetrapoda are:
Below is a general cladogram for the living vertebrates and their fossil kin:
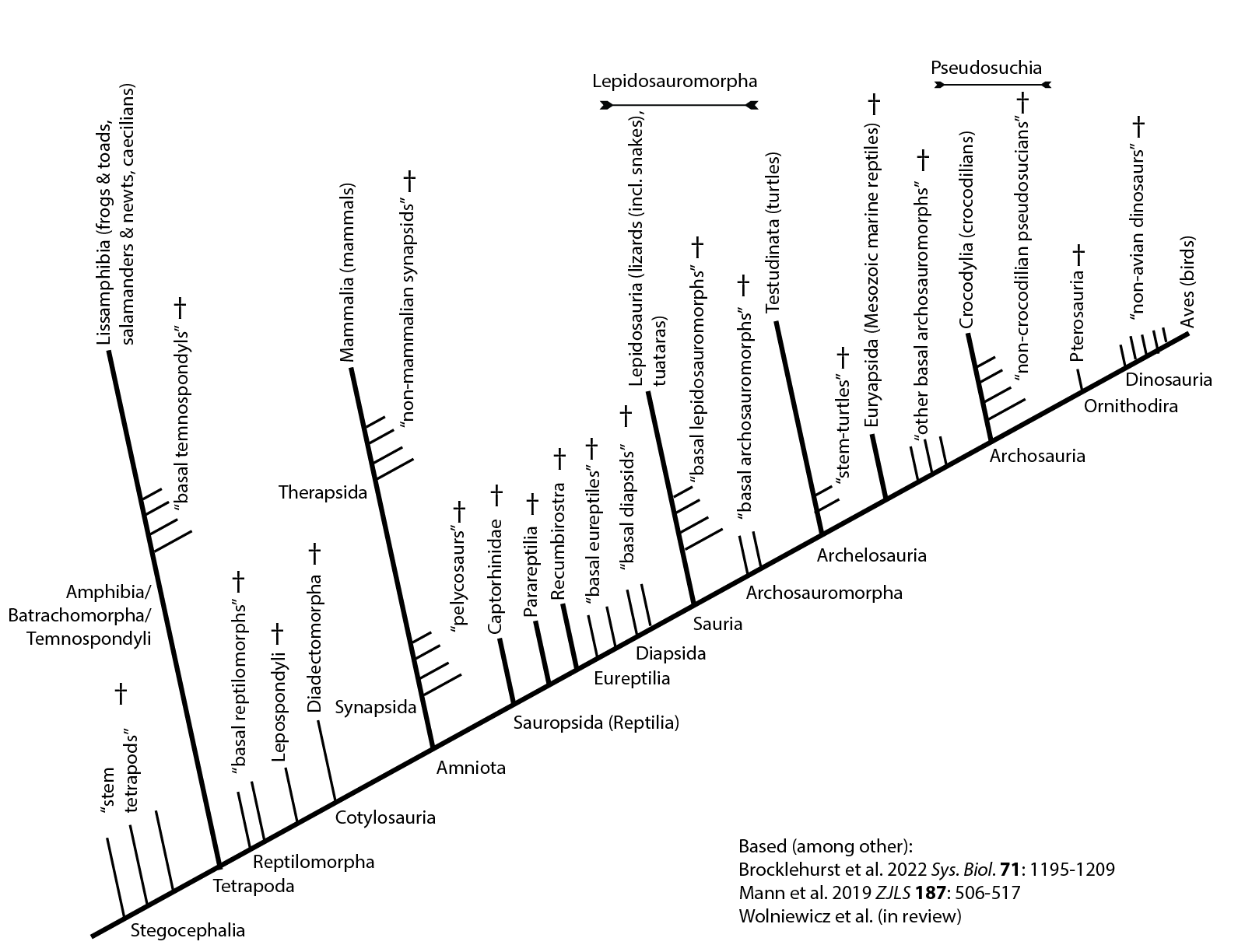
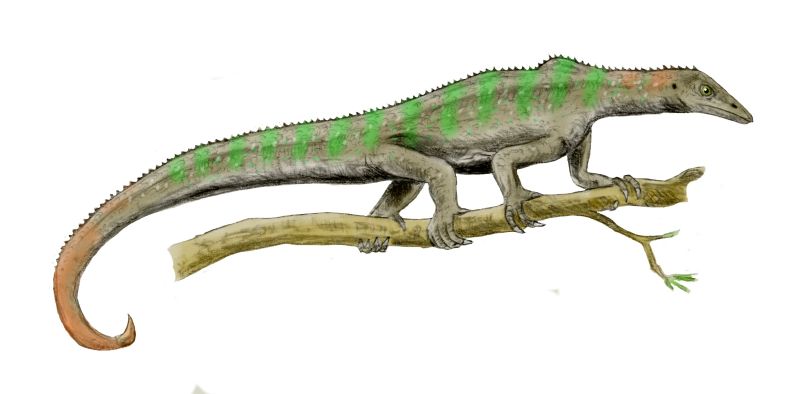
A subclade of the sauropterygian fish, stegocephalians are first known from the Devonian. Relative to their kin, stegocephalians (tetrapods and their extinct relatives) are distinguished by:
In the Mississippian many stegocephalians (such as Crassigyrinus) were still essentially "fingered fish" incapable of movement on land. Others, such as Pederpes, had well-developed arms and legs, and digit counts of five on each limb: these animals probably were fairly terrestrial as adults.
Tetrapoda proper is generally restricted to the crown group of Stegocephalia: roughly, the concestor of frogs and dogs, and all of its descendants. Exact taxonomic make-up of Tetrapoda is hindered by uncertainty about the status of various clades of amphibian-grade tetrapods: are the inside the crown-group, or its sister taxon?
Living tetrapods are characterized by:
Basal tetrapods, and indeed many of the basal stegocephalians, were "amphibians" in a sense: they had a fully aquatic larval phase (a "tadpole" in the broad sense) with external gills which metamorphosed into an adult more capable of life on land. However, "amphibians" in this sense are simply a grade relative to the amniote clade.
And many of the Paleozoic "amphibians" were decidedly unlike modern lissamphibians. Some were marine, whereas contact with salt water would be fatal to most living amphibians. Additionally, many of these "amphibians" retained fish-like dermal scales.
[NOTE: the "scales" of fish and amphibian-grade tetrapods and the "scales" of amniotes are not homologous structures by any means! The former are dermal elements, which in many early fish groups contained dentine and enamel (although many derived actinopterygian groups, and convergently the amphibian-grade tetrapods, lost dentine and enamel in their scales and instead had simply keratinous structures. The scales of reptiles (and presumably the scaled stem-mammals) are epidermal keratinous structures. It is the gastralia (belly ribs) of amniotes which are the direct homologues of the dermal scales of fish and amphibian-grade tetrapods (although it may be that osteoderms (bony armor) of tetrapods represent a deep homology with these structures).]
There are some interesting parallels in the terrestrialization of hexapods and tetrapods. In both, the limbs were already present for use in some form of benthic locomotion. In both a neck was evolved to allow them to feed on land. And in both the basal forms had aquatic larvae that use a newly evolved gill structure (rather than the ancestral gills of their outgroups) in order to breath. However, there are major differences as well: for instance, fully terrestrial insects often have more profound metamorphoses from larvae to adult, whereas newborn amniotes are often far more similar to their adults than tadpoles are to amphibian-grade adults.
Tetrapoda divides into two main branches. Batrachomorpha includes modern frogs and salamanders (collectively Batrachia) and everything closer to them than to amniotes: this clade is also called "Amphibia" (although this term comes with a lot of baggage) and Temnospondyli. Reptilomorpha comprises amniotes and everything closer to them than to batrachians.
There has been considerable debate over the relationships of the living amphibians. Many studies suggest that Batrachia (containing the Anura/Salienta (frogs & toads) and the Caudata/Urodela (salamanders & newts)), the Gymnophiona/Apoda (the caecilians), and the extinct Albanerpetontidae (a clade of scaled superficially salamander-like tailed, four-legged amphibians that survived up into the Pliocene) form the monophyletic group Lissamphibia. Although we think of lissamphibians as having naked skin, that is in fact a trait of Batrachia only, as there are subcutaneous collagenous dermal scales with a slight bit of mineralization in gymnophionans, and better developed scales in albanerpetontids. Lissamphibia is characterized by dramatic reductions of the skeleton (for example, the ribs no longer extend far from the vertebrae).
However, some recent studies contend that "Lissamphibia" is paraphyletic, with Batrachia nested among the batrachomorphs, but with gymnophionans closer to amniotes. In other recent studies, batrachians and gymnophionans arise from entirely different clades within temnospondyls. But there are still numerous phylogenetic analyses which find batrachians, gymnophionans, and albanerpetontids as a clade exclusive within the Paleozoic and early Mesozoic temnospondyls.
The stem-lissamphibian temnospondyls were a highly diverse group of the late Paleozoic and Triassic, and survived (at very low diversity) up into the mid-Cretaceous. Some represented batrachomorph versions of the alligator morph; others were more narrow-snouted fish eaters. The largest were, at 4 m length, the largest of the amphibian-grade tetrapods.
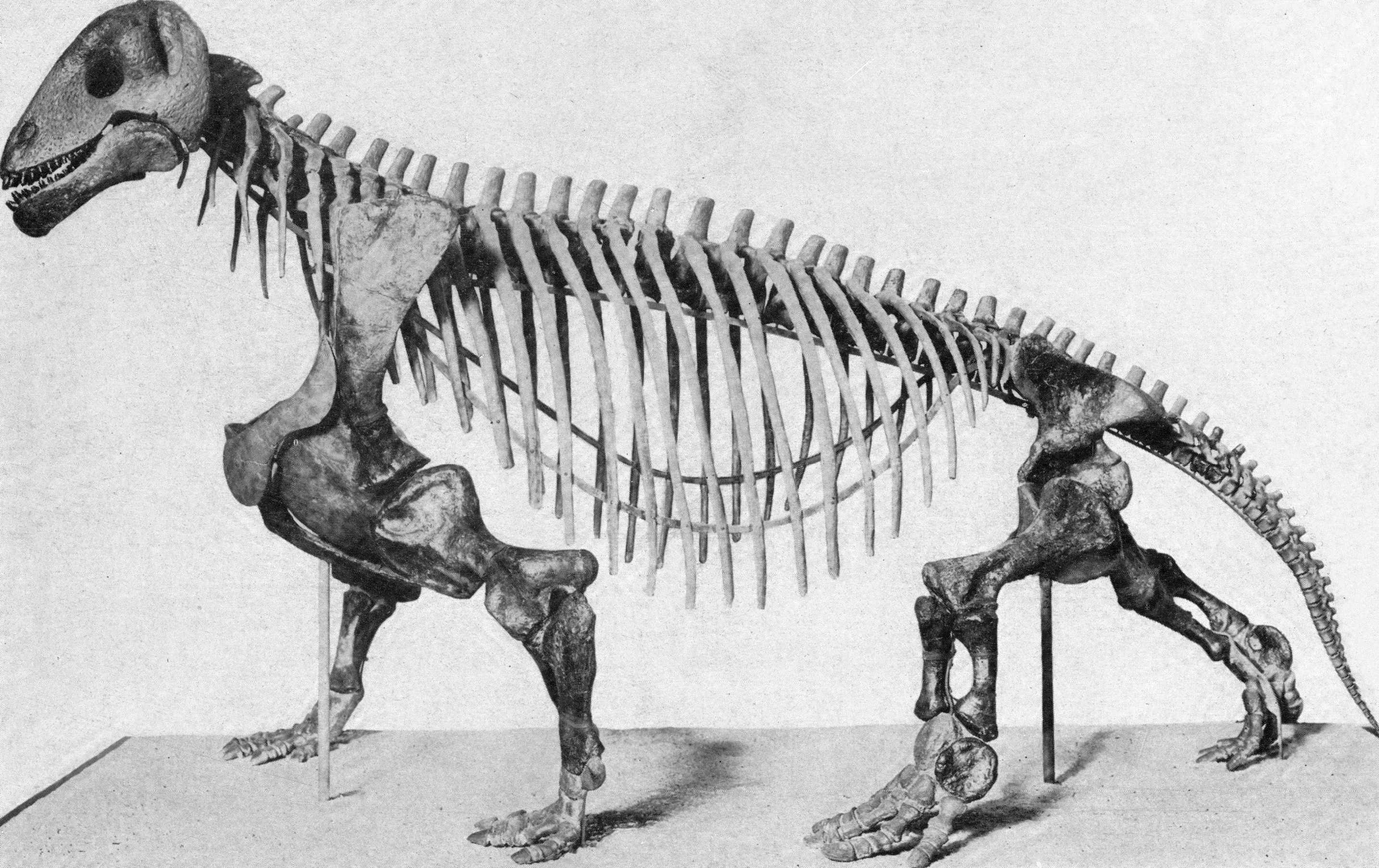
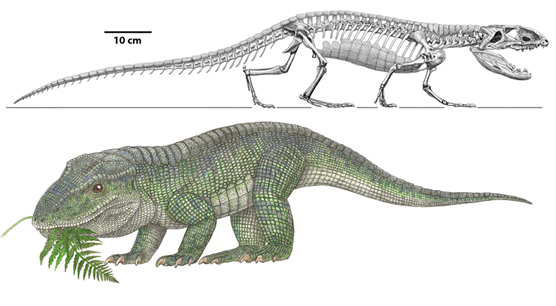
Basal (amphibian-grade) reptilomorphs include a number of late Paleozoic taxa, such as: Anthracosauria (a predominantly marine group of narrow-skulled piscivores: although this may actually be a stem-tetrapod group); Lepospondyli (a diverse group of dubious monophyly including the boomerang-headed Diplocaulus); Semouryiamorpha(another late Paleozoic clade, with very terrestrialized adults but fully aquatic larvae); and Diadectomorpha (a clade or grade representing the sister group or groups of Amniota). Diadectomorphs may have actually had an amniotic egg: at present, their form of reproduction is unknown. Diadectomorphs included both large-bodied terrestrial herbivores and nearly as large terrestrial carnivores.
Mammals, turtles, tuataras, lizards (including snakes), crocodilians, and birds collectively form the living part of Amniota, the crown-group reptilomorphs. Amniotes are characterized first and foremost by the amniotic egg. With an egg with an external shell and an amniotic sac in which the embryo can develop, tetrapods were able to fully leave the water.
The oldest amniotes are from the Pennsylvanian. As the egg itself is not preserved, we must rely on other anatomical features to recognize fossil members of Amniota:
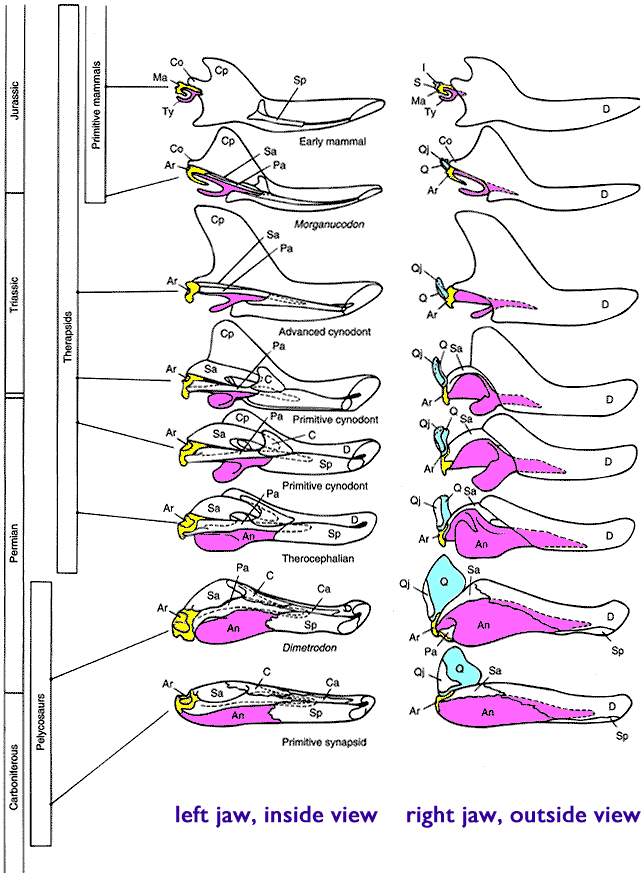
Synapsid phylogenetic relationships are a thing of beauty, with regards to stratigraphic ranges and orders of
appearance:
Basal synapsids were once collectively referred to as "Pelycosauria", but this is a paraphyletic group. During the Carboniferous and Early Permian Epoch primitive synapsids radiated into many different forms:
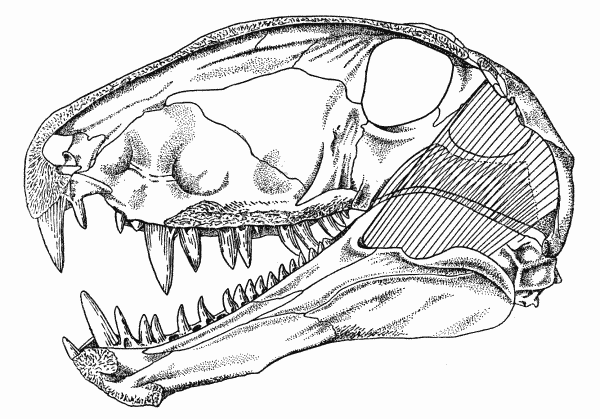
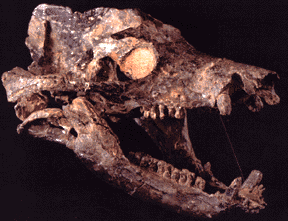
The early synapsids evolved themselves into extinction: that is, they were replaced in the Middle and Late Permian Epochs by the Therapsida: the advanced synapsids. Once called the "mammal-like reptiles", they are not true reptiles. Instead, they are the advanced branch of the synapsid phylogeny. They differed from earlier synapsids by:
The Middle and Late Permian therapsids included:
Ancestrally, all vertebrates are cold-blooded (warm their bodies primarily using sunlight). However, some
evidence suggests that the advanced therapsids of the Late Permian may have had elevated metabolisms (that
is, were at least partially warm-blooded):
Additionally, some therapsids seem to have had parental care of the young, keeping them in burrows.
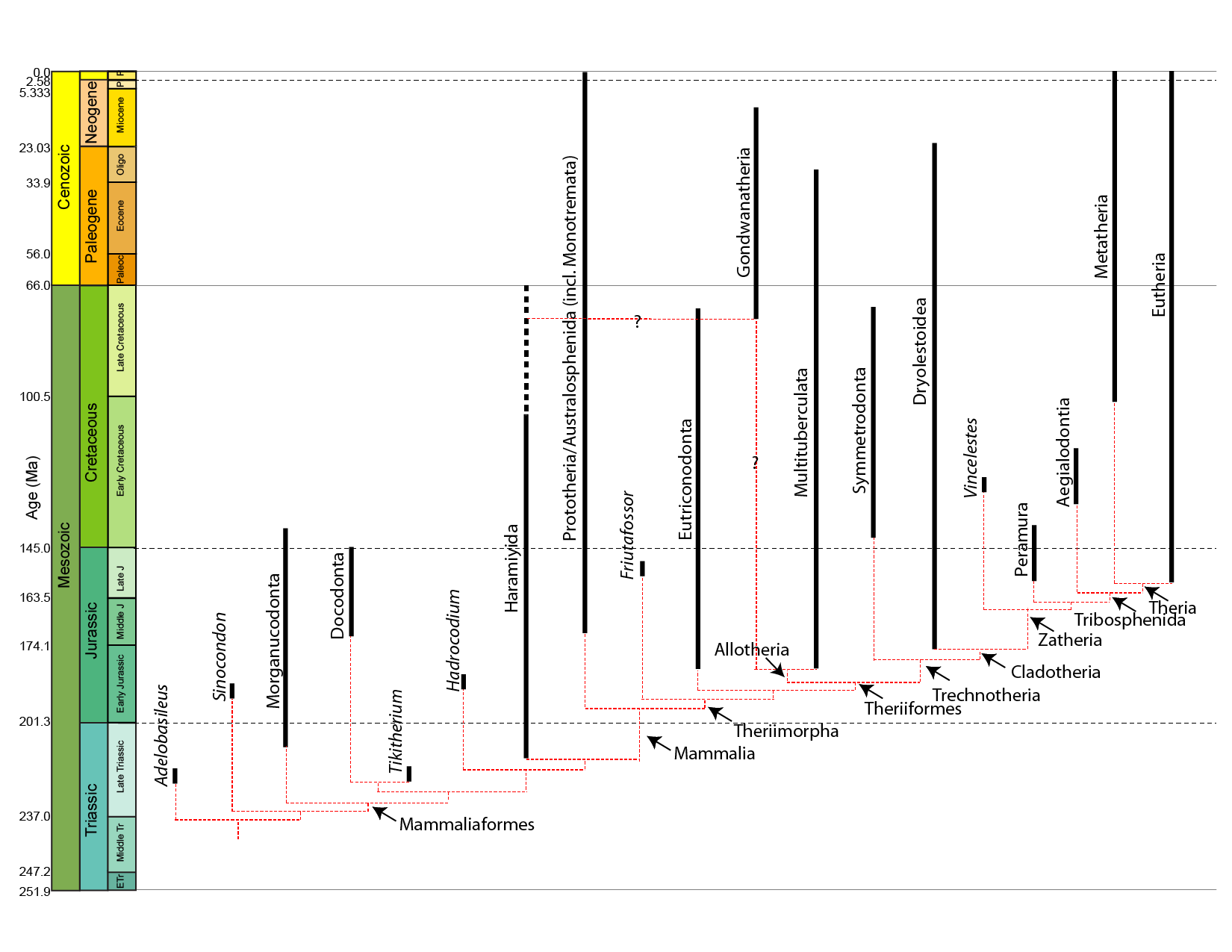
Mammals and their closest relatives (more properly Mammaliaformes) appear in fossil record the same time as dinosaurs, in Late Triassic:
Mammals are very advanced therapsids synapsids.
True mammals (Mammalia) are found from the Early Jurassic onward (or the Late Triassic onward, if the Haramiyida are crown-mammals, as they appear to be in some phylogenies).
Like birds, many of the features that characterize modern mammals don't fossilize:
On the other hand, some mammalian features are preservable:
Many features limited to Mammalia among living amniotes were probably found in their closest non-mammalian therapsids relatives. For example, we can't say for certain when warm-blood, fur, sweat & mammary glands show up. We can determine a few of these, though:
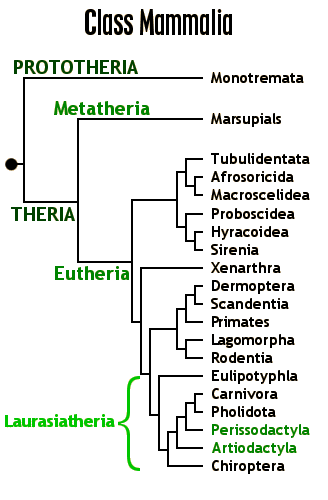
Living mammals are divided into three clades:
However, mammal diversity in the Mesozoic was MUCH different.
Many groups of Mesozoic mammals have long since died out. And some Mesozoic mammal groups survived the end of the Cretaceous, but have since died out.
Most Mesozoic mammals very small (shrew-to-house cat sized, with a few badger-sized forms in the Cretaceous); mammals only become large AFTER extinction of non-avian dinosaurs.
Oldest mammaliaforms of the Late Triassic and Early Jurassic were fairly small. But by Middle and Late Jurassic, there were already some specialized mammals:
Total group-marsupials (Metatheria) and total group-placentals (Eutheria) are united in the clade Theria. Therians are united by various skeletal (parasagittal stance, some dental, etc.) and soft-tissue (nipples, external ears, etc.) features. Therians include the metatheres and eutheres, which diverged by the earliest Late Jurassic.
Metatheria (marsupials and their extinct relatives):
Eutheria (placentals and our extinct relatives):
During the Cenozoic, tremendous radiation of the placental (and to a lesser degree, marsupial) mammals. Among the evolutionary patterns:
"Reptilia" has traditionally been a paraphyletic assemblage (e.g., all non-bird, non-mammal amniotes). Currently the name is used for a clade that contains the living turtles, tuataras, lizards (including snakes), crocodylians, and birds, and all taxa closer to them than to mammals. Given that "reptile" has been used in the paraphyletic sense, many taxonomists prefer Sauropsida for this clade. The crown members of the clade are characterized by:
Several Permian groups (plus one group that survived through the Triassic) form a clade Parareptilia. Among the parareptiles were the aquatic
Mesosauridae, the long-legged (and possibly bipedal at a full run) Bolosauridae, the large armored herbivorous Pareiasauria, and the smaller herbivorous
Procolophonoidea of the Permian through Triassic. Turtles were once thought to lie among the parareptiles, but are now known to be diapsids.
The more diverse clade of sauropsids is Eureptilia. Basal eureptiles are generally anapsid-skulled, "lizard-like" animals (of course, true lizards are actually highly specialized animals!). Among the eureptiles, however, the quick-snapping bite associated with the diapsid skull condition evolved:
In diapsids, the hindlimbs are often longer than the forelimbs, allowing for a bipedal run for at least short distances.
Among the basal diapsids are some highly specialized forms, such as the gliding coelurosauravids and the arboreal
drepanosaurids.
The extraordinarily diverse Sauria includes two branches: Lepidosauromorpha andArchosauromorpha. Lepidosauromorphs include the extinct gliding Kuehneosauridae, the Rhynchocephalia (a clade containing the living tuarara as well as extinct relatives such as the aquatic pleurosaurs), and the Squamata (lizards).
In the Mesozoic, two major highly specialized groups of squamates evolved: the Cretaceous marine Mosasauroidea and the long-bodied Serpentes (aka the snakes), which first arose in the Jurassic. While the former perished in the K/Pg extinction, the snakes survived and (during the Neogene) underwent a major radiation to become a dominant group of small-bodied predators in meadow environments.
Pan-Testudines (total-group turtles) seem to be early diverging diapsids; recent molecular and even some morphological studies place them as early-branching members of Archosauromorpha. Early stem-turtles such as Eunotosaurus and Middle Triassic Pappochelys are unarmored, toothed, and show a diapsid skull. Middle Triassic Odontochelys
Paleontologists often characterize amniotes by their temporal fenestration: that is, the presence and number of openings in the rear of the skull to accommodate jaw muscles. Primitive amniotes, like primitive tetrapods in general, have the anapsid condition: that is, no temporal fenestration. Some amniotes have only the lower or infratemporal fenestra, in what is termed the synapsid condition.

In others, there is both an infratemporal and a supratemporal fenestra: the diapsid condition:

Tetrapods fall into two clades: Synapsida (aka Theropsida), mammals and our extinct kin; and Sauropsida (aka "Reptilia" in the cladistic sense), the other ones (technically Lacerta, Crocodylus, and everything closer to them than to Homo).
SYNAPSIDA: MAMMALS AND OUR KIN
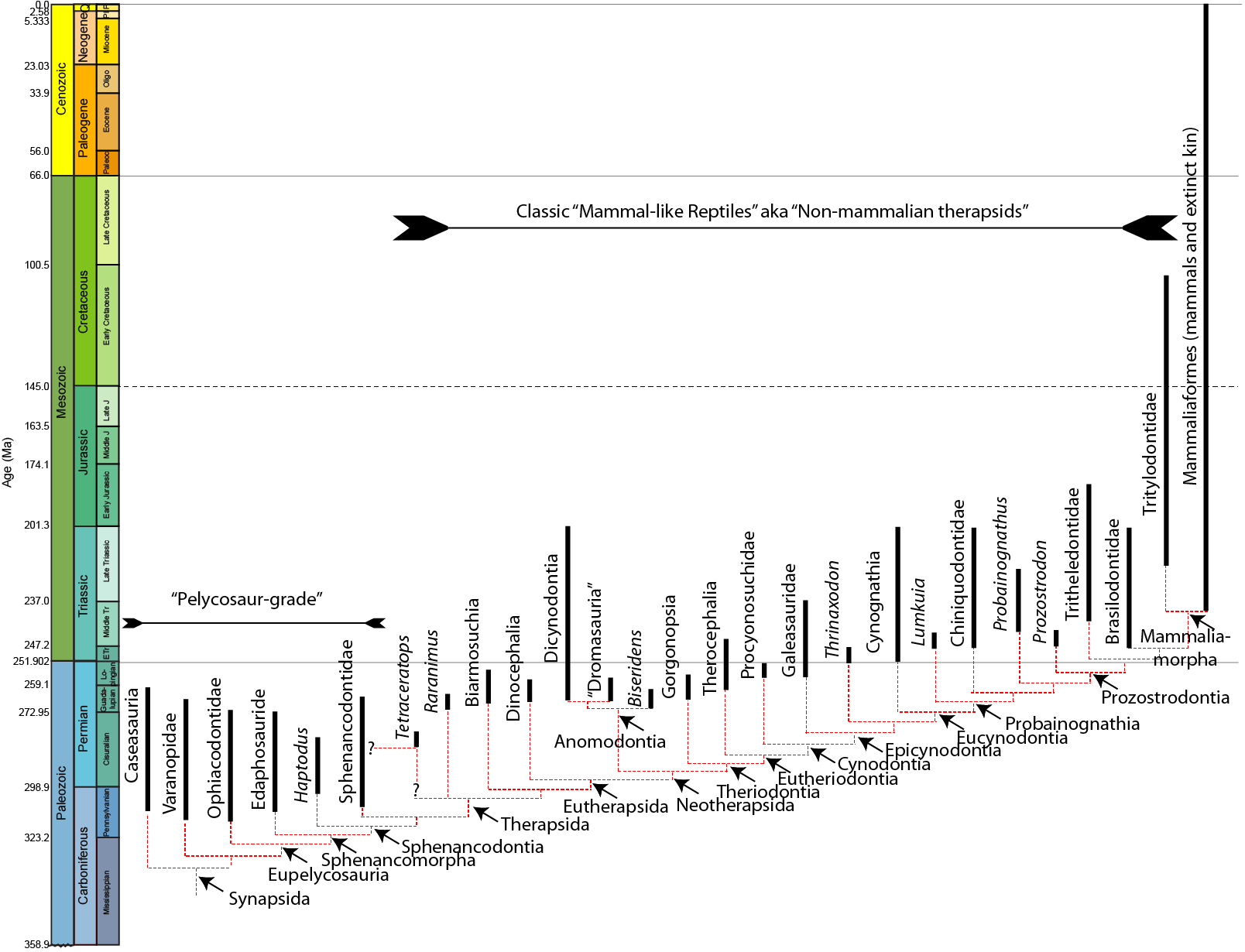


As large-bodied terrestrial animals during an age of relative continental isolation, many morphotypes evolve convergently in the different regions.
SAUROPSIDA: THE GREAT REPTILIAN RADIATION
Reptile interrelationships have been extensively studied. Historically a primary point of contention is the phylogenetic position of turtles (see discussion below). However, recent studies have challenged the relationships of the basal forms, and even found that the (normally thought to be early diverging) Parareptilia lies within Diapsida, and that some groups thought to be synapsids are also really diapsids! Consequently, identifying the basal condition for many traits in Sauropsida is currently

Another diapsid (and likely early archosauromorph) clade is the highly successful group Euryapsida, the premier group of marine reptiles of the Mesozoic. Members of this group possessed ovoviparity or even true vivipary (the ability to develop the young inside the body until they were capable of swimming on their own). Thus freed from the land, some groups of euryapsid became highly specialized swimmers. Basal forms of the Triassic tended to be nearshore animals, but two groups of pelagic euryapsids arose:
The archosauromorphs include numerous other branches beyond turtles and euryapsids. Other primitive archosauromorphs were the broad-skulled herbivorous Rhynchosauria, the freshwater Choristodera (one branch of which, the long-snouted Champsosauridae, survived well into the Paleogene), the sometimes extraordinarly-long-necked Tanystropheidae, and the large herbivorous Trilophosauridae.
By far the most successful group of archosauromorphs (and indeed one of the most successful groups of tetrapods) are the Archosauriformes: crocodilians, birds, and their extinct kin:
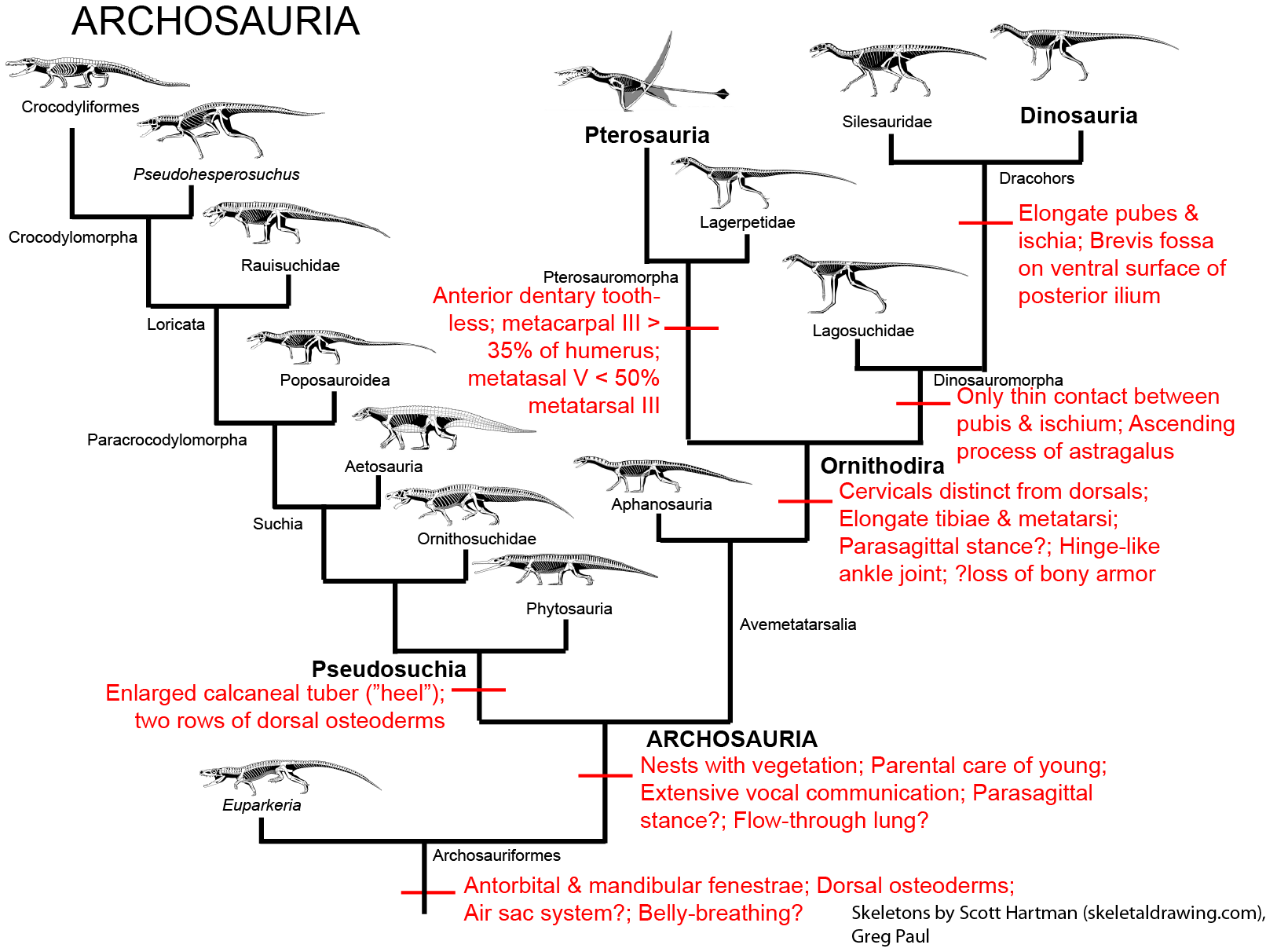
First appearing at the tail-end of the Permian, early archosauriforms contained both flesh- and fish-eating forms. Archosauriforms are distinguished by the presence of additional openings in the skull:

as well as:
It is not certain if these early archosauriforms had the behavioral traits found in both the living groups of Archosauria (that is, crocodylians and birds). Because both crocodylians and birds share the following derived traits, however, it is fairly certain that at least the concestor of all Archosauria had them, and passed them on to its descendants:
Crown-group archosauriforms (Archosauria) contains two clades: Pseudosucha, crocodilians and their extinct kin; and Avemetatarsalia (more typically referred to by one of its subclades, Ornithodira), birds and their extinct kin.
Pseudosuchians were a major group of Triassic animals, including:
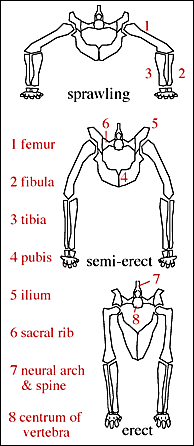
The pseudosuchians include some of the first terrestrial animals to exceed the size of oxen and hippos. Most of them could stand with a semi-erect posture of the limbs, and a few had the fully-erect (that is, parasagittal gait). All pseudosuchians except for Crocodylomorpha perished in the Triassic-Jurassic extinction. During the Mesozoic, crocodyliforms radiated to produce numerous forms, including the marine Thalattosuchia, snub-nosed plant-eaters, giant semi-aquatic ambush predators, and terrestrial carnivores. Cenozoic Crocodylia remains a major component of the freshwater ecosystems.
For all their diversity, however, pseudosuchians are dwarfed by their ornithodiran sister taxa. Basal ornithodirans differed from typical diapsids by having elongate lower limb elements, suggesting that they were even faster than typical reptiles. All (except perhaps for the most primitive) had cervical vertebrae greatly modified so that they look very different from dorsal vertebrae, but instead could allow the neck to form an S-shaped curve (hence the name of the group).
One branch of the ornithodirans was the
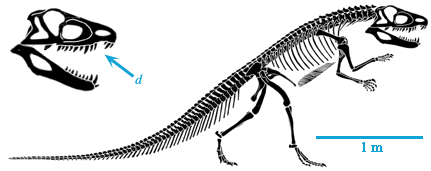
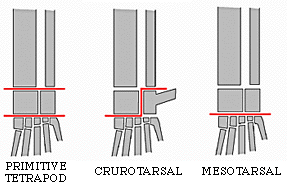
The last group of tetrapods to consider is Dinosauromorpha (in Dr. Holtz's opinion, the BEST clade of vertebrates), which first appeared in the Middle Triassic of Argentina. The taxa modified the hindlimbs further to evolve:
The combination of these traits allowed the little dinosauromorphs to run and accelerate in a bipedal mode all the time, not just at top speeds like typical diapsids. The parasagittal gait and hinge-like ankle also allowed dinosauromorphs the ability to move more actively and constantly rather than only in short bursts of speeds; thus, they were striders. Additionally, although early dinosauromorphs were small (~30 cm long for Marasuchus, 1-3 m long for silesaurids and basal dinosaurs, etc.), the presence of limbs directly underneath the body meant that this lineage to grow to much larger size than any previous clade while still remaining terrestrial and mobile (sprawlers relegated to a semi-aquatic life if they became too big to support their weight).
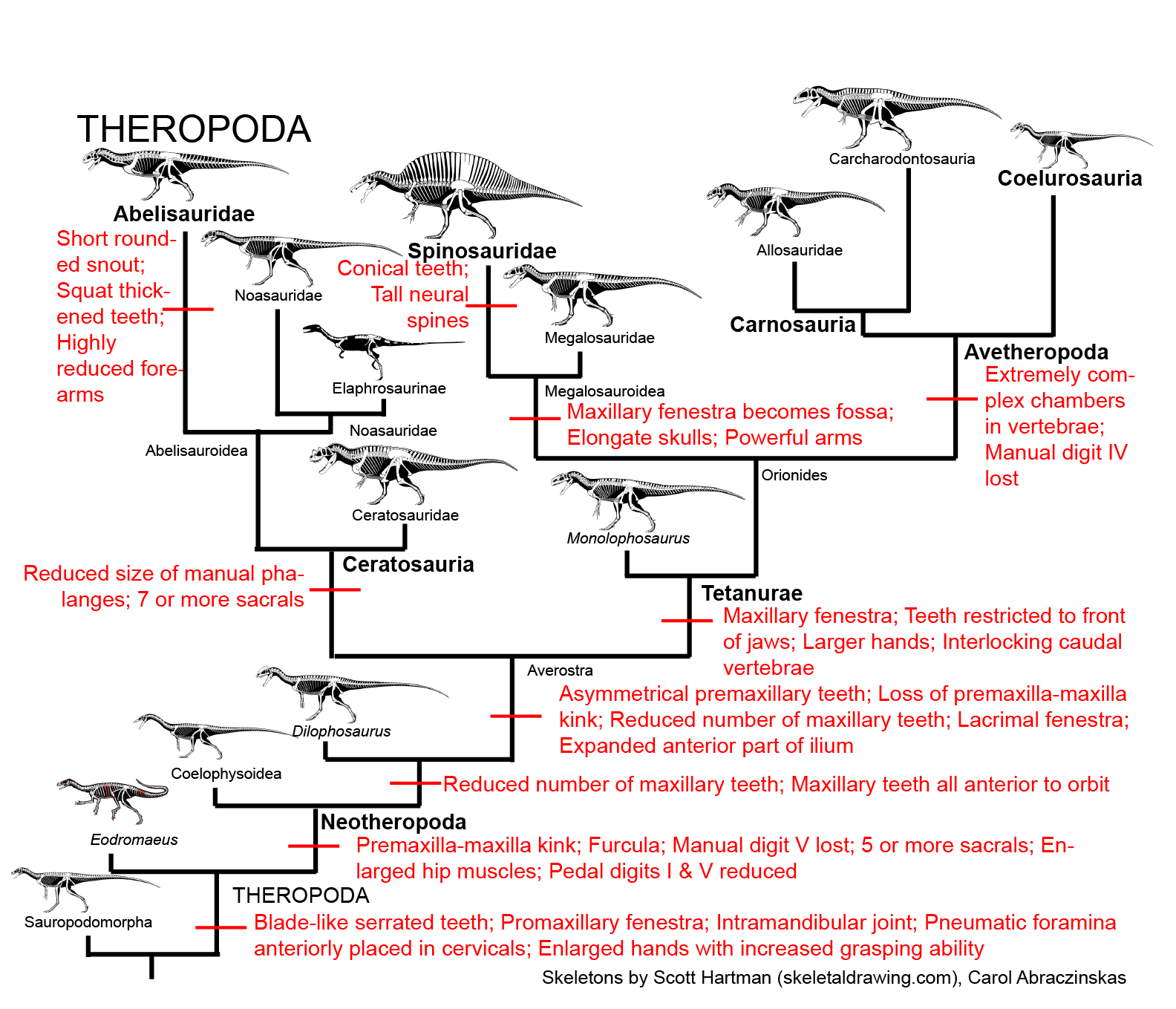
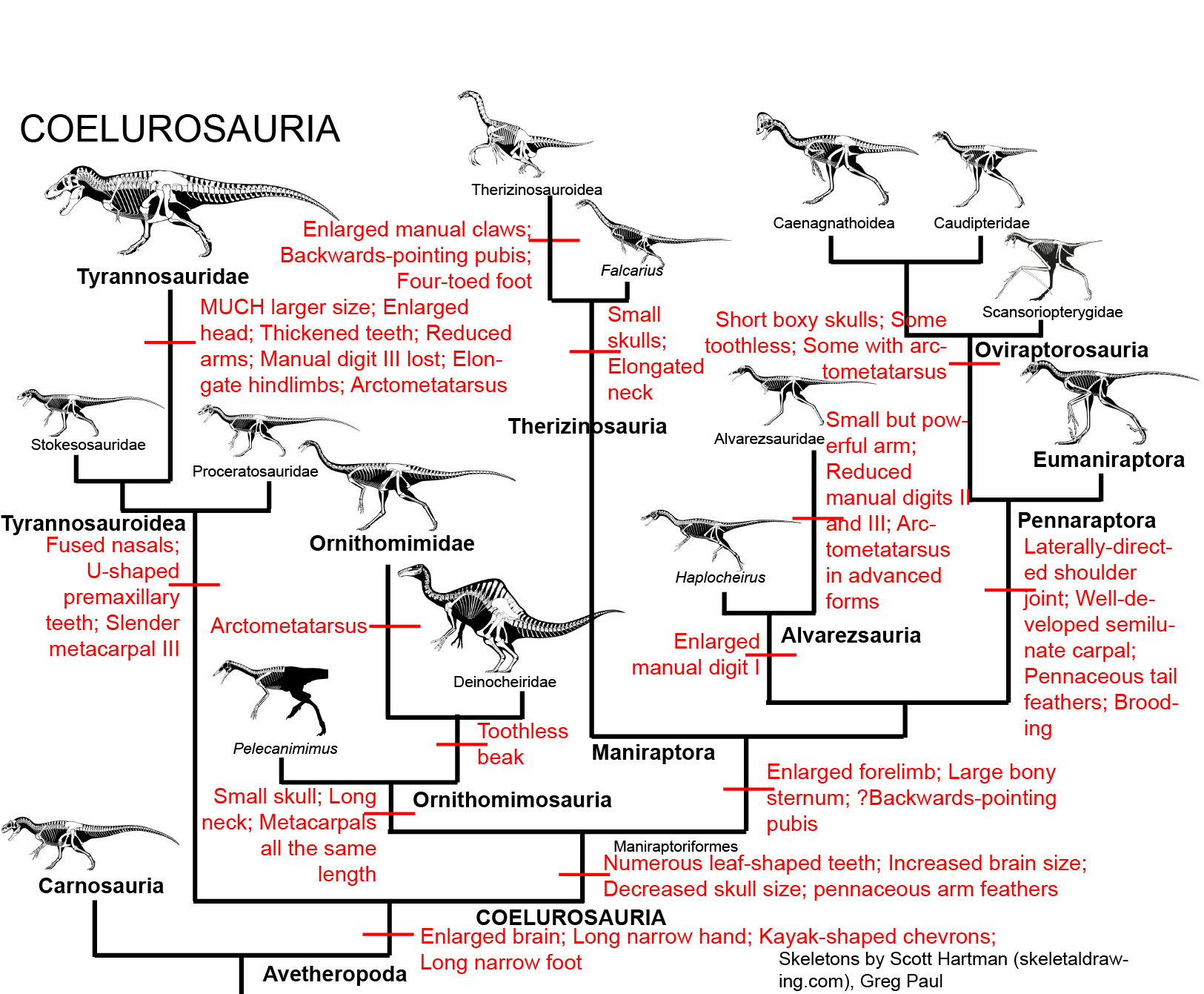
By the Late Triassic, the true Dinosauria had evolved. Within this diverse lineage were:
In their history, tetrapods (and especially amniotes) did some fairly amazing things. Here are some of the larger trends and successes within the history of Tetrapoda:
Herbivory: To our mind, herbivory (eating macroscopic plant parts) seems a normal part of "Nature". But think back on the invertebrate groups we have studied, and how few of them actually munched on plants. In the marine realm this makes sense, as most primary productivity is in the form of unicellular phytoplankton. Consequently there are often many trophic levels of microscopic and very small macroscopic feeders.
Even in the terrestrial realm, actual herbivores are limited to just a few clades. Some gastropods are plant munchers. And while larval lepidopterans (aka caterpillars) and the most diverse clade of beetles eat leaves and stems and so forth, most other groups of insects which feed on plants do so by sucking juices out rather than cropping off plant parts.
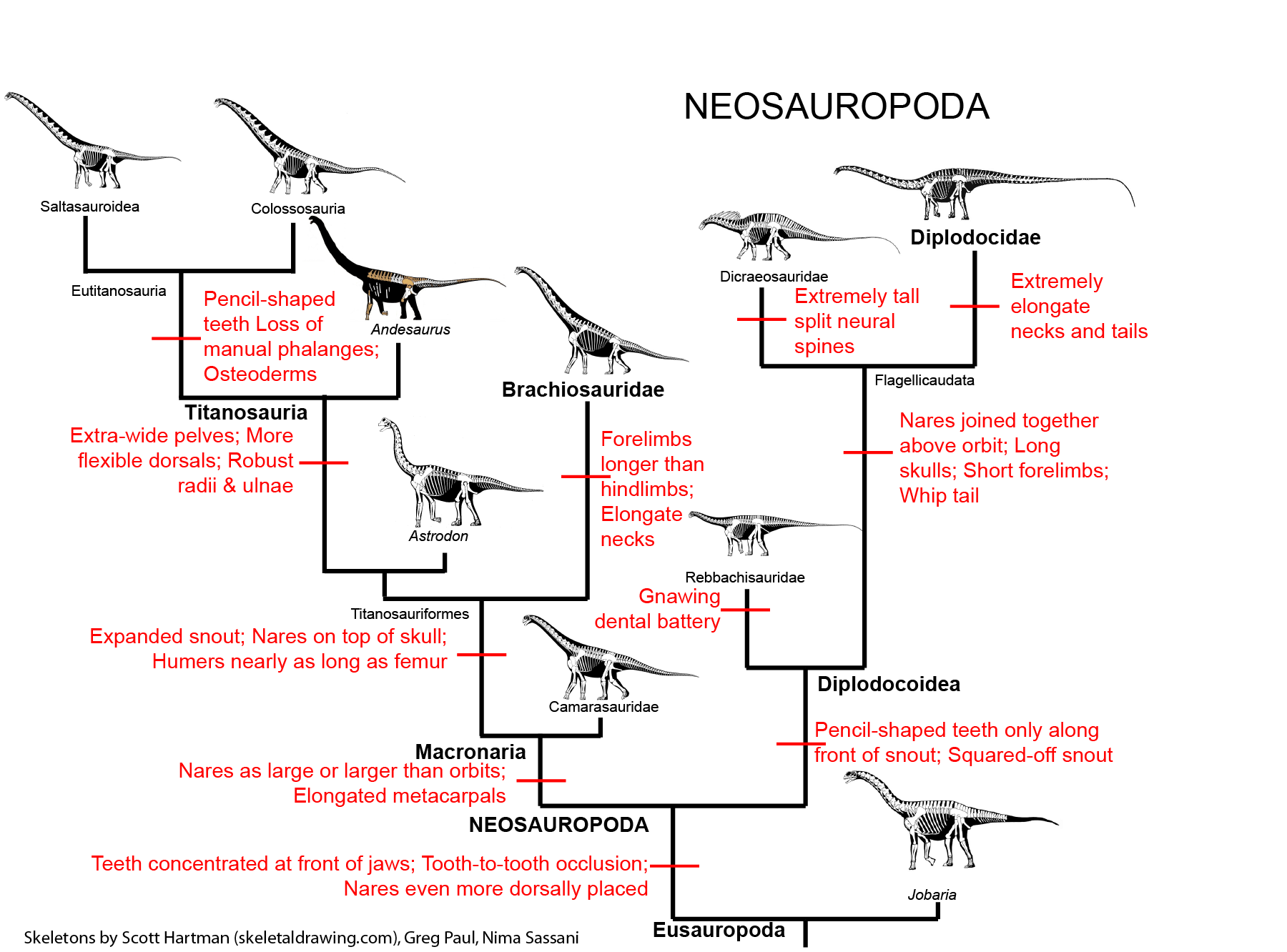
In contrast, many amniotes (and near-amniotes, like some diadectomorphs) ate plants: from tiny seed-eating rodents and songbirds, through medium-sized animals like tortoises, horses, and elephants, to actually large animals like sauropod dinosaurs. (Professional bias there.) As a result of this, there are often highly complex feeding adaptations (most especially chewing teeth) among many herbivorous amniotes. (Additionally, when we look at paleoecology we'll see how having large-bodied plant eaters modifies ecosystems.)
Size: Even the smallest tetrapods are macroscopic, and thus dwarf the smallest arthropods and such. And the modal mammal and modal sauropsid is much larger. Even a moderate-sized amniote like a dog or iguana is a giant compared to all but the largest gastropods and bivalves, and these are giants compared to (for instance) brachiopods. Most significantly, though, is the upper boundary: not even the biggest cephalopod (by far the biggest of invertebrates) comes within an order of magnitude or two of the mass of large whales and dinosaurs. The largest marine amniotes are larger even than the biggest chondrichthyians and actinopterygians
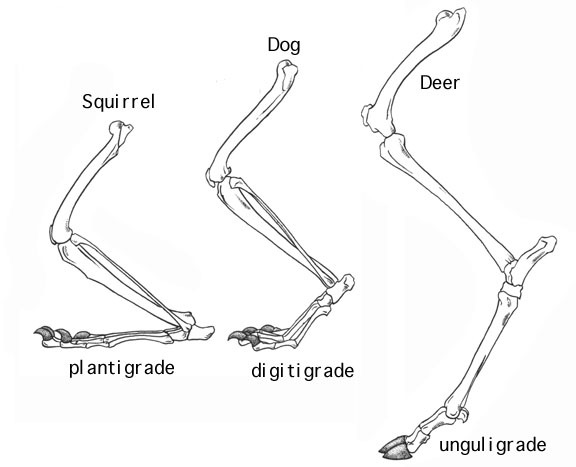
Terrestrial Locomotion: Ancestrally tetrapods move by lateral undulations of the body: essentially "land-swimming". But this is modified in many different ways, such are the jumping of frogs, the legless slithers of snakes, the bipedal bursts of lizards, and notably the independent origins of parasagittal striding and running in therian mammals and archosaurian sauropsids.
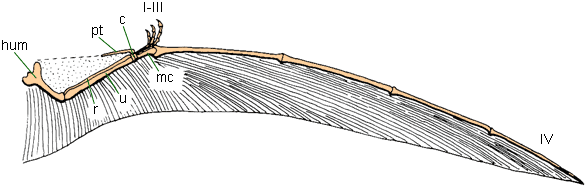
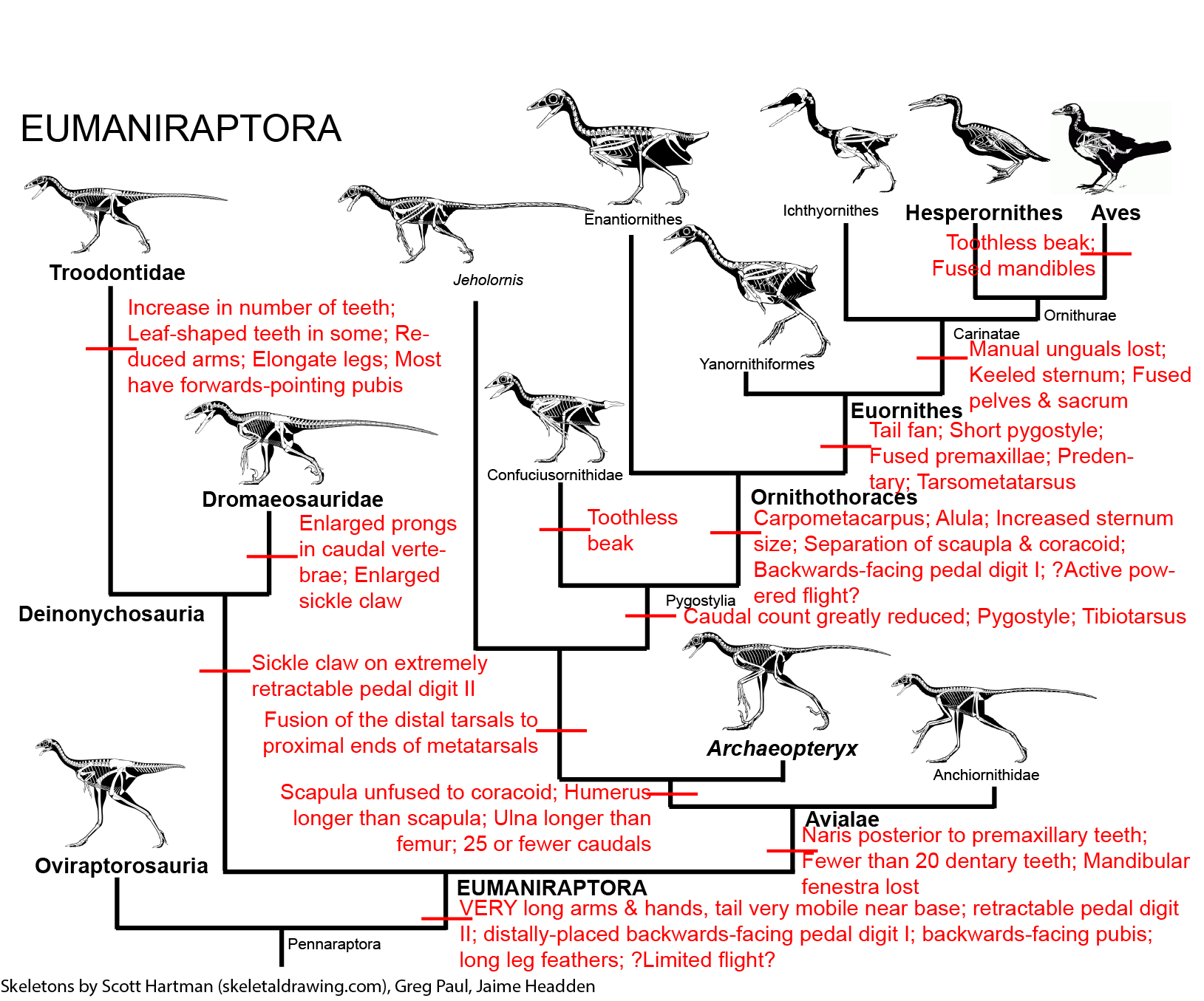
Aerial Locomotion: Gliding and parachuting is widespread: flying fish; flying frogs; flying lizards; flying snakes; sugar gliders (gliding marsupials); flying squirrels; colugos, or "flying lemurs". In addition, there were various extinct gliding reptiles and mammals in the Permian and Mesozoic. But powered flight arises three times in Amniota: once (bats) in mammals and twice (pterosaurs and within the bird-stem) in ornithodiran archosaurs.
The Reconquest of the Seas: After all the effort to adapt to life on land, amniotes seem to be in a rush to return to the seas. A great many clades are moderately evolved for aquatic feeding and swimming, and many evolved adaptations that required them to live nearly all the time in the water: ichthyosaurs, plesiosaurs, two separate radiations of marine crocodilians, mosasaurs, cetaceans (whales), pinnipeds (seals and sea lions), sirenians (sea cows), and more. Interestingly, with the exception of the herbivores like sirenians, the marine amniotes often wind up as apex predators (or nearly so) in their communities.
Elevated Metabolisms, including Endothermy ("Warm-bloodedness"): The basal physiological condition for vertebrates is ectothermy (so-called "cold-bloodedness"): having body heat mostly derived from the ambient environment. Thus, if the external conditions get too cold the animal might not function properly, and slows down into torpor. This restricts the range of habitats that the animals can live in. An alternative is to evolve an increased amount of mitochondria, whose function is primarily to generate heat internally and maintain a constant body temperature (and consequently higher levels of activity). This endothermy ("warm-bloodedness") requires a substantial increase in the amount of food needed (sometimes up to 10x that of ectotherms of the same mass.)
Why evolve such an expensive trait as endothermy? Some suggestions have included:
Endothermy has evolved multiple times in vertebrates: in various chondrichthyians; in a few actinopterygians (in particular, among tuna and among billfish); and diversely among amniotes. Therian mammals are fully warm-blooded, with monotremes with not quite as high a metabolic rate; there is substantial evidence for high metabolic rates well down into Therapsida. All birds are full warm-blooded, and this trait seems to be broadly present in extinct ornithodiran archosaurs. New evidence suggest that ichthyosaurs and plesiosaurs independently evolved this condition within Euryapsida, as did mosasaurs. Additionally, some animals such as the lizard Salvator and various snakes are temporarily warm-blooded during brooding.
High Intelligence and Complex Social Structures: This is not true of all tetrapods, of course. But consider how many mammals and birds have social structures in which individuals might rise and fall in rank, develop coalitions, friendships, and so on. This is not at all like the pre-programmed castes of "eusocial" insects. More broadly, birds, mammals, crocodilians, and even some turtles and lizards show problem-solving abilities, engage in play, and so forth. Only cephalopods come close to vertebrate levels of intelligence.
Vertebrate (and especially tetrapod) paleontology has some particular difficulties:
To Syllabus.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}