

Key Points:
•Sauropodomorpha is a clade of herbivorous dinosaurs characterized by small skulls, long necks, and (at least ancestry) leaf-shaped teeth.
•The Triassic and Early Jurassic taxa--basal sauropodomorphs, core prosauropods, and near-sauropods--represent a transitional series starting as meter-long obligate bipeds to 10 meter long obligate quadrupeds. These include the only successful Triassic dinosaurs.
•The remaining taxa--all obligate quadrupeds--form the clade Sauropoda. Sauropods were among the most successful of all dinosaur groups, and were a major part of the herbivorous niches in every region from the Middle Jurassic until the end of the Cretaceous. Sauropods include the largest animals ever to live on land.
•Various primitive sauropod branches lie outside the derived Neosauropoda. The latter include the long-skulled, pencil-toothed Diplodocoidea (divided into the enormous Diplodocidae, the tall-spined Dicraeosauridae, the gnawing Rebbachisauridae) and the large-nosed Macronaria (including the upwardly-built Brachiosauridae and the extraordinarily diverse Titanosauria).
MAJOR GROUPS OF SAUROPODOMORPHS
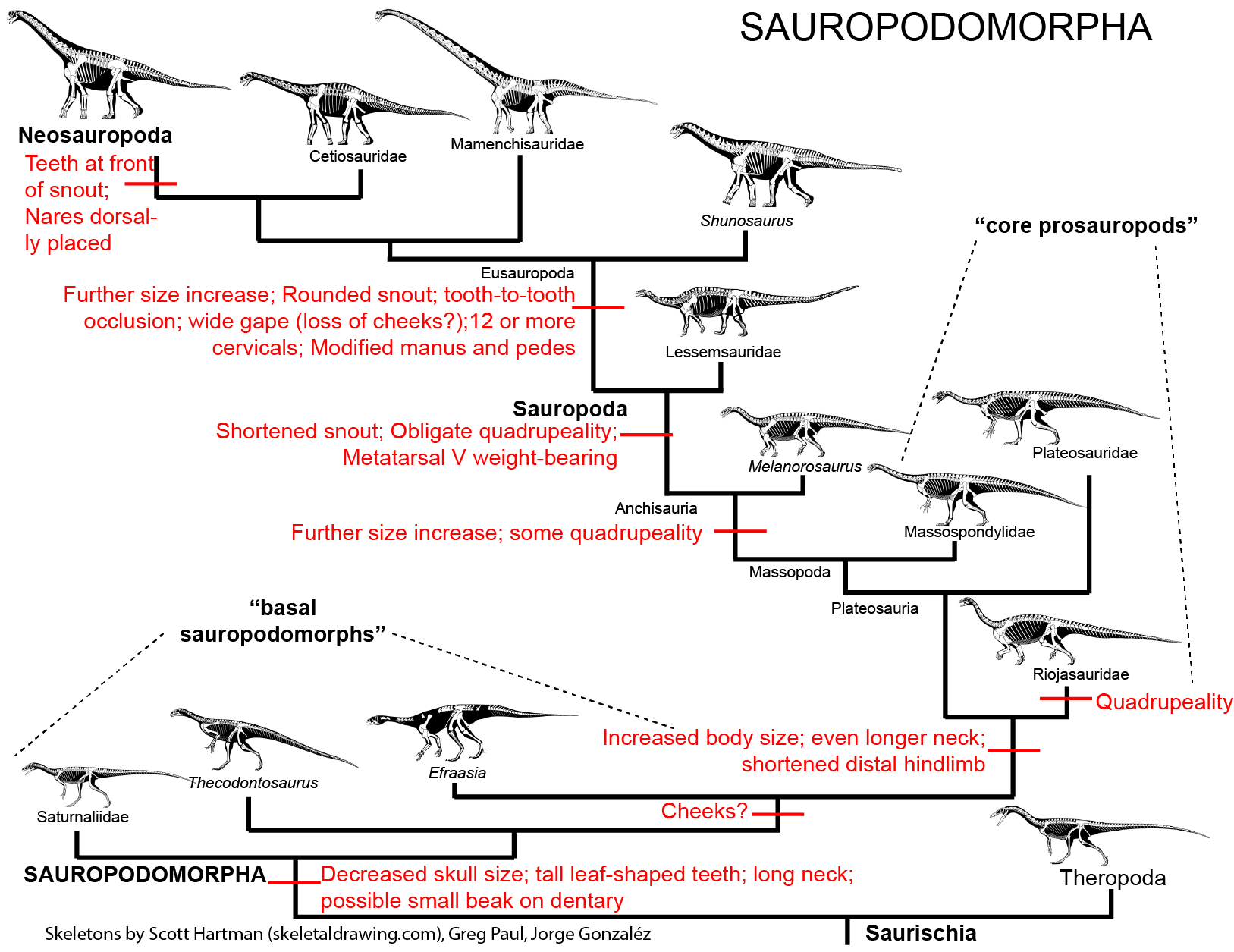
The derived sauropodomorphs are the Sauropoda (lizard feet), giant obligate quadrupeds that include the largest land animals of all time. Primitive sauropodomorphs (i.e., sauropodomorphs other than sauropods) were traditionally grouped together as "Prosauropoda". Modern studies differ as to whether all prosauropods form a paraphyletic grade (with no major prosauropod groups more closely related to each other than to sauropods), while others recognize a monophyletic "core Prosauropoda" composed of plateosaurids, riojasaurids, massospondylids, etc. This course follows the first (paraphyletic grade) hypothesis (see cladogram and phylogeny above).
BASAL SAUROPODOMORPHS
Specializations of the most primitive sauropodomorphs relative to other saurischians include:
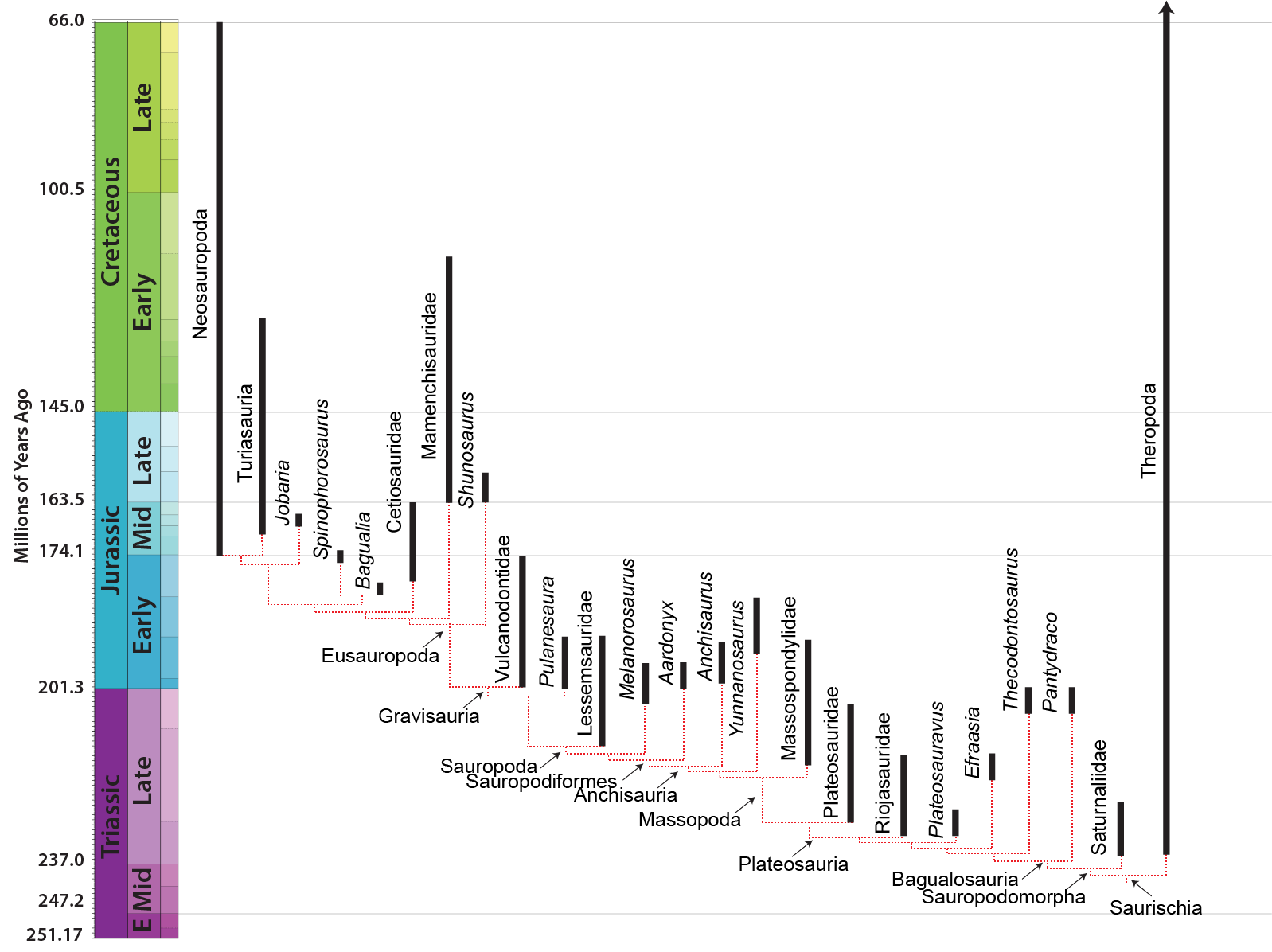
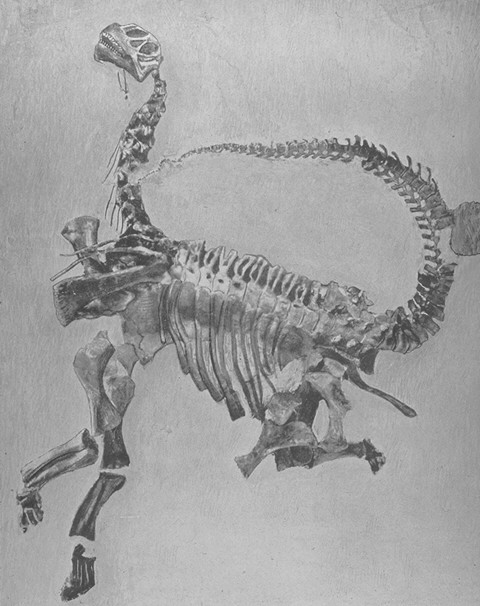
These forms were small (~1-2.5 m long), bipedal plant eaters. Their long necks would allow them to feed on higher leaves than most of their contemporaries. Recent work suggest a paraphyletic series of basalmost sauropodomorphs. The most primitive sometimes are found to form one or more clades (depending on the exact tree shape, these groups are called Saturnaliidae and Guaibasauridae), although they sometimes are found to be a paraphyletic series relative to all other sauropodomorphs. These taxa are restricted to the Carnian (the early part of the Late Triassic). The most completely known of these are Panphagia and Eoraptor of the Late Triassic of Argentina, and newly discovered Mbiresaurus of Zimbabwe. (Eoraptor was long thought to be a basal theropod dinosaur, hence its name.) Chromogisaurus lived alongside them. The clade gets its name from Saturnalia of the Late Triassic of Brazil: its contemporary Guaibasaurus maybe a a saturnaliid, or a basal theropod, or a basal saurischian.
The teeth of the basalmost sauropodomorphs are less phyllodont than the more derived taxa, and some teeth approach the ziphodont (blade-like) teeth of theropods in shapes: consequently, the earliest sauropodomorphs were almost certainly omnivores rather than strict herbivores. (In fact, that is the etymology for the name of Panphagia, "all eater" or "omnivore".) Intriguingly, the recently-discovered Buriolestes has honest-to-goodness ziphodont teeth.
Slightly more derived basal sauropodomorphs Thecodontosaurus, Asylosaurus and Pantydraco of Late Triassic England; Efraasia of Late Triassic Germany; and Bagualosaurus and Plateosauravus of Late Triassic South Africa. Mussaurus of the Early Jurassic of Argentina has often been considered one of these basal forms, but new evidence places it as a more advanced form (a near-sauropod).
Like most saurischians, basal sauropodomorphs and core prosauropods had a "wrap around overbite": the toothrow of the mandible fit entirely within the tooth row of the upper jaw. Efraasia and later prosauropods (and basal sauropods) seem to have had a small fleshy cheek similar to (but smaller than) those interpreted for ornithischians. Less convincingly, some evidence suggests a small keratinous beak in front of the dentary (and possibly premaxilla) of basal sauropodomorphs and/or core prosauropods. The diets of basal sauropodomorphs would have included at least some plants, but these may have been more omnivorous than later, more derived sauropodomorphs.
CORE PROSAUROPODS
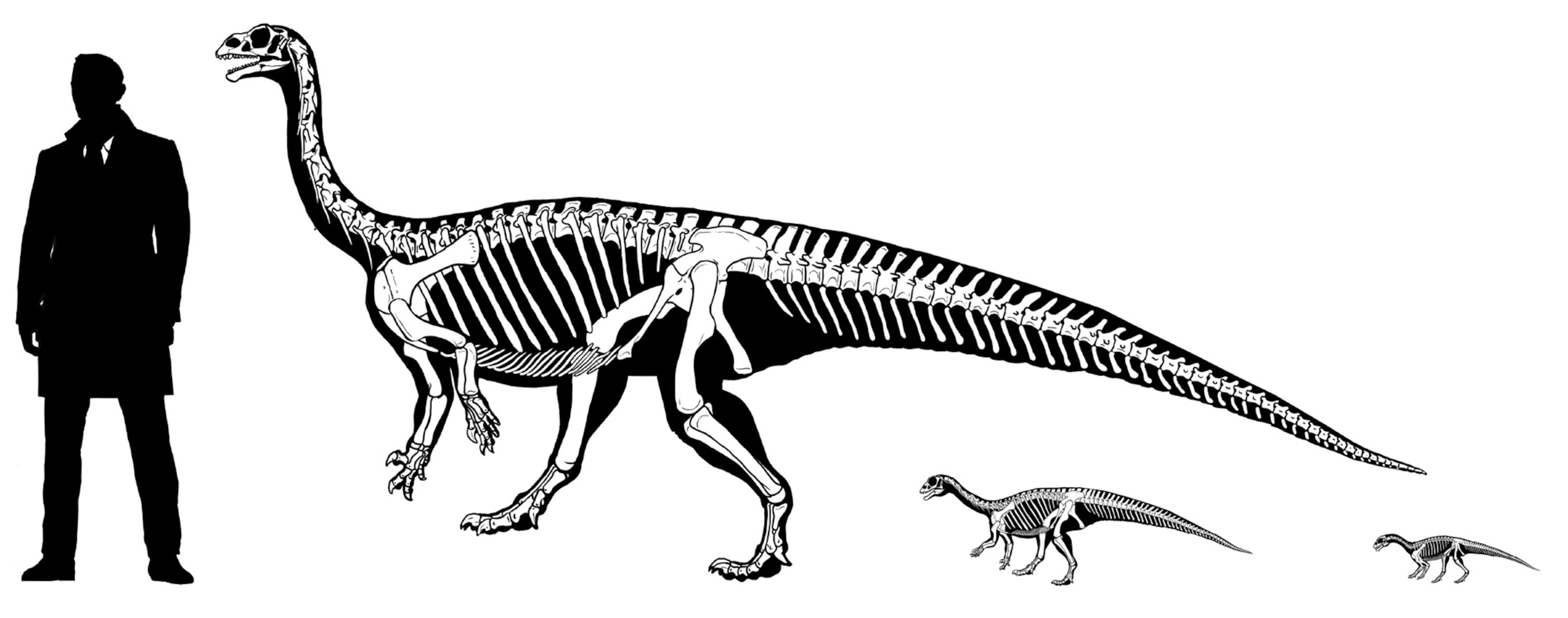
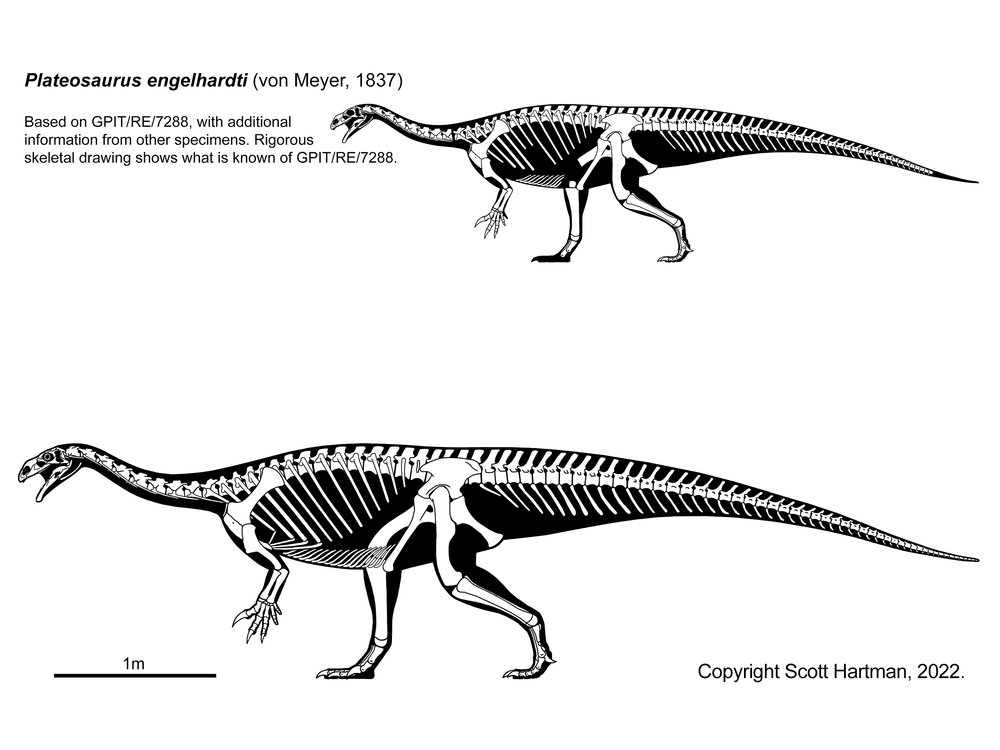
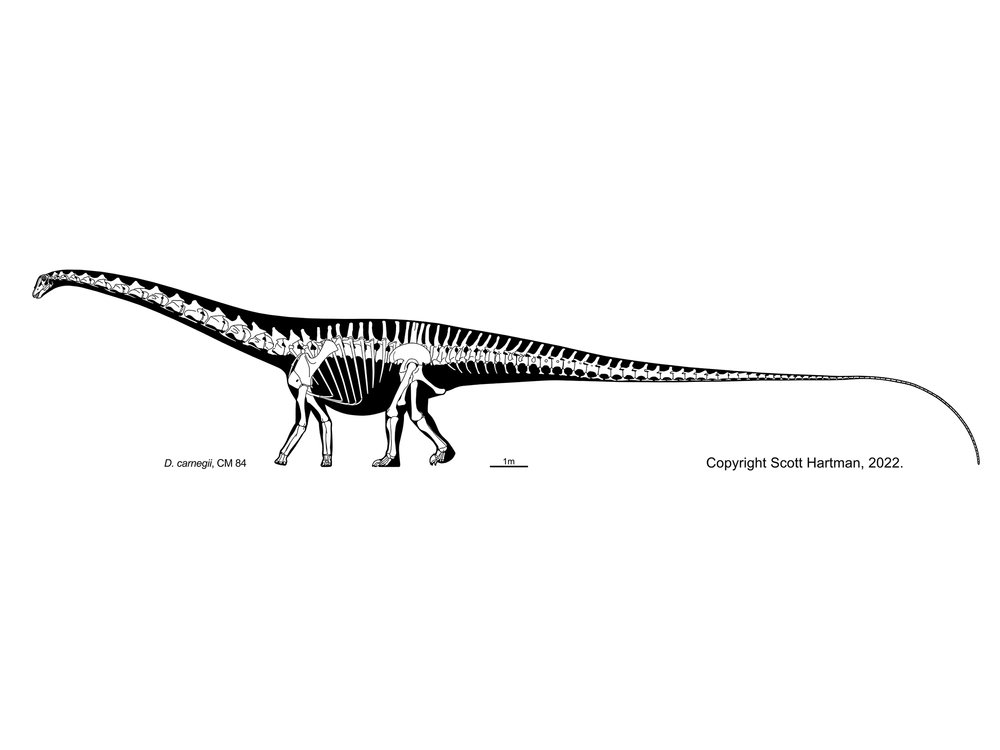
Plateosauridae (including Late Triassic European Plateosaurus), Riojasauridae (including Late Triassic Argentine Riojasaurus), and Massospondylidae (including Early Jurassic South African Massospondylus, Argentine Adeopapposaurus, Antarctic Glacialisaurus, North American Seitaad, and Chinese Lufengosaurus): considered by some to form a monophyletic "Prosauropoda" and by others a paraphyletic series relative to melanorosaurids and sauropods. Core prosauropods were the dominant group of herbivores worldwide in the later Late Triassic and Early Jurassic: in some communities they were up to 95% of the known biomass. They represent the first time dinosaurs dominated a major way of life. (They are the only common dinosaurs in Late Triassic Europe, for example.) Traditionally interpreted as facultative bipeds, new studies of their forelimbs (especially forearms and wrists) show that they were not used for locomotion in most of these and so these dinosaurs were obligate bipeds, although slowly built ones. (In Riojasaurus, however, limb proportions show that they were almost certainly primarily quadrupedal.) Like basal sauropodomorphs, the core prosauropods retained the ancestral dinosaurian grasping hand. Their distal hindlimb (tibia, fibula, and metatarsus) was shorter than in other saurischians (a trait they passed on to Sauropoda).
The expansion (in terms of both physical size and diversity) of core prosauropods may be a result of the Carnian Pluvial Event. Many new forms of tree-forming plant evolved or diversified in the wake of this wet climate episode, and likely provided the new resource that these larger dinosaurs could exploit.
Larger than the basalmost sauropodomorphs, most core prosauropods range from 3 to 8 m longer (although some grew larger, up to or more than 10 m). Their necks were proportionately even longer than in their ancestors. They have been found in many environments: from swamps to deserts.
Although very common in the Late Triassic and Early Jurassic, no core prosauropods are known after the end of the Early Jurassic. They may have "evolved themselves into extinction", by being outcompeted by their specialized descendants: the sauropods.
"NEAR-SAUROPODS" AND THE PROSAUROPOD-SAUROPOD TRANSITION
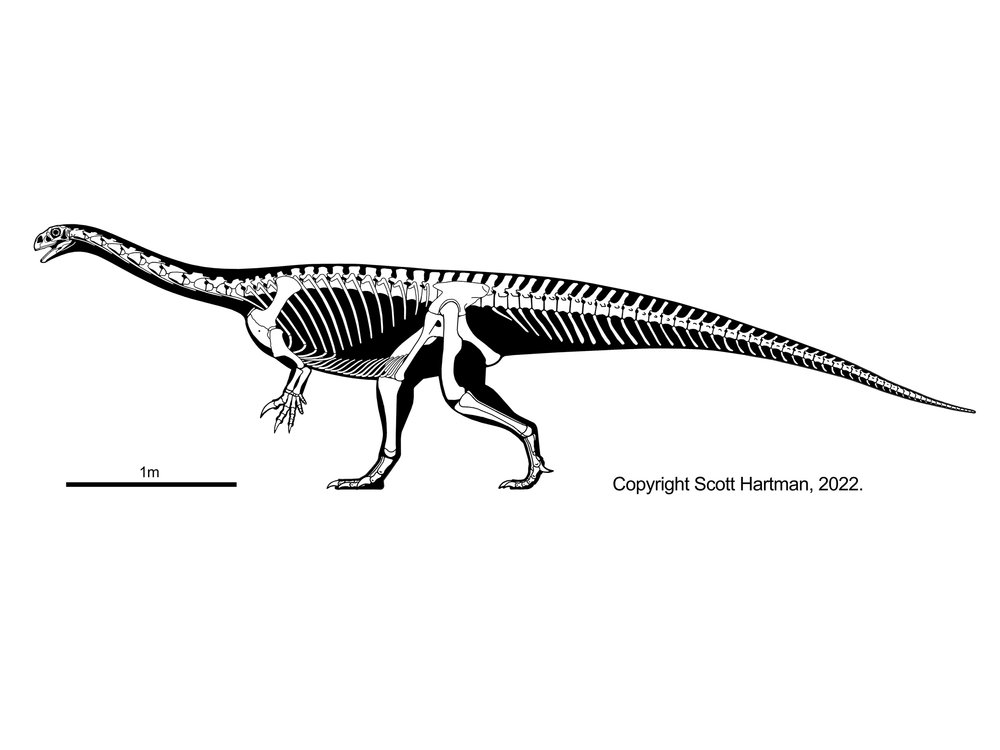
The most advanced prosauropods (Anchisaurus, Yunnanosaurus, Lamplughsaura of India, Leonerasaurus of Argentina, and Melanorosauridae (such as Late Triassic-Early Jurassic South African
Melanorosaurus) are considered basal sauropods by some workers. For the purpose of this course, we'll restrict "Sauropoda" to Saltasaurus and everything sharing a more recent common ancestor with it than with Melanorosaurus, and use the unofficial name "near-sauropods" for those sauropodomorphs phylogenetically intermediate between Massospondylus and Sauropoda.
Like sauropods, the near-sauropods were typically larger than the more primitive sauropodomorphs: in this case, some 10-13 m long or more as adults and perhaps 5 tons. (Anchisaurus [sometimes called Ammosaurus] is a tiny exception: only as big as the basal sauropodomorphs.) Recent work shows that it might have actually been a quadruped. Aardonyx of the Early Jurassic of South Africa is among the last branches of Sauropodomorpha which was an obligate biped. Most near-sauropods were at best facultative bipeds, and many may have been obligate quadrupeds. Most add another sacral vertebrae (total of four).
The evolution of sauropodomorphs shows a correlated progression towards larger body size, smaller heads, longer necks, and quadrupedality. This may have been related to two (not mutually exclusive) selective forces:
These transitional near-sauropod forms are Late Triassic-to-Early Jurassic in age.
SAUROPODA
Postcranially and in size (10-13 m length), basal sauropods do not differ much from melanorosaurids and other near-sauropods. But early sauropod skulls have been transformed:
Additionally, sauropods are recognizable from other sauropodomorphs by:
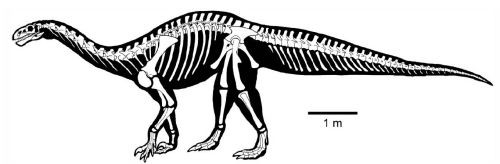
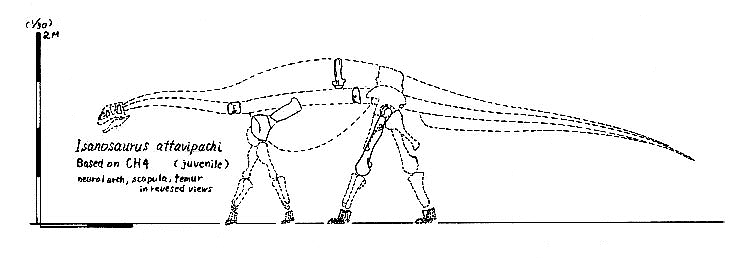
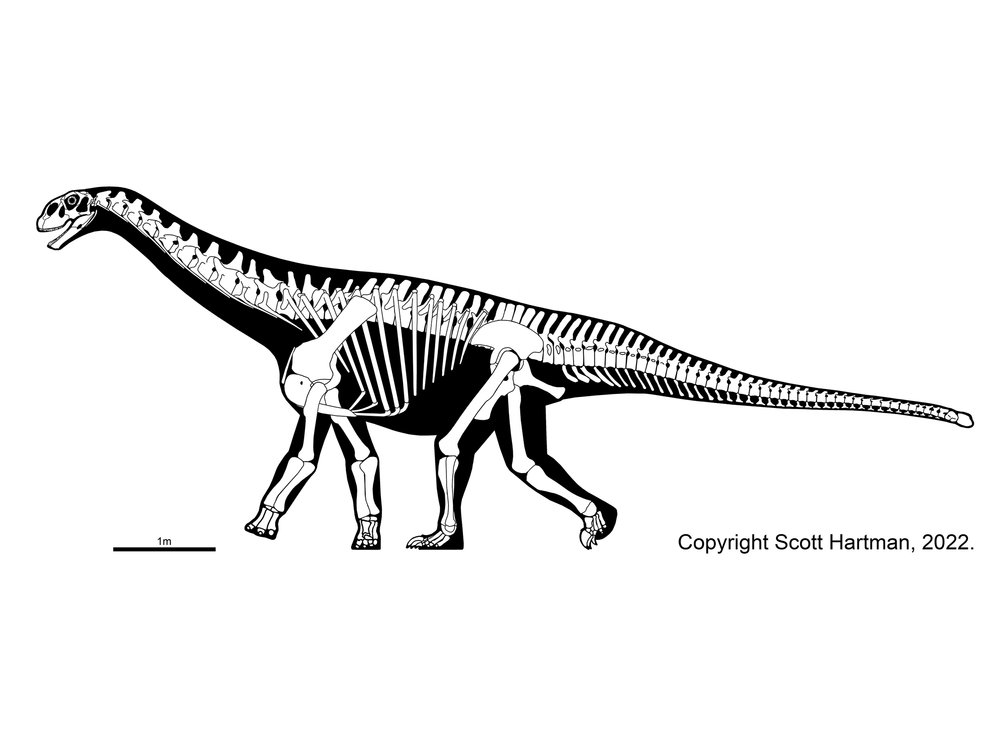
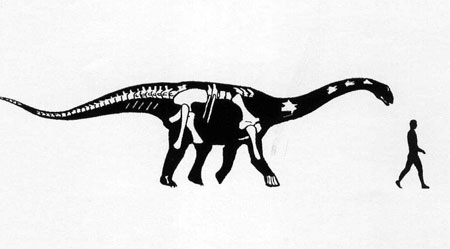
The oldest and most primitive of the sauropods are the Late Triassic-to-Early Jurassic Lessemsauridae. Named after Argentine Late Triassic Argentine Lessemsaurus, other lessemsaurids include its contemporary Ingentia and Early Jurassic South African Antetonitrus and the 12-ton giant Ledumahadi. Other basal sauropods include Early Jurassic South African Pulanesaura, Thai Isanosaurus, Chinese Gongxianosaurus, and the Vulcanodontidae.
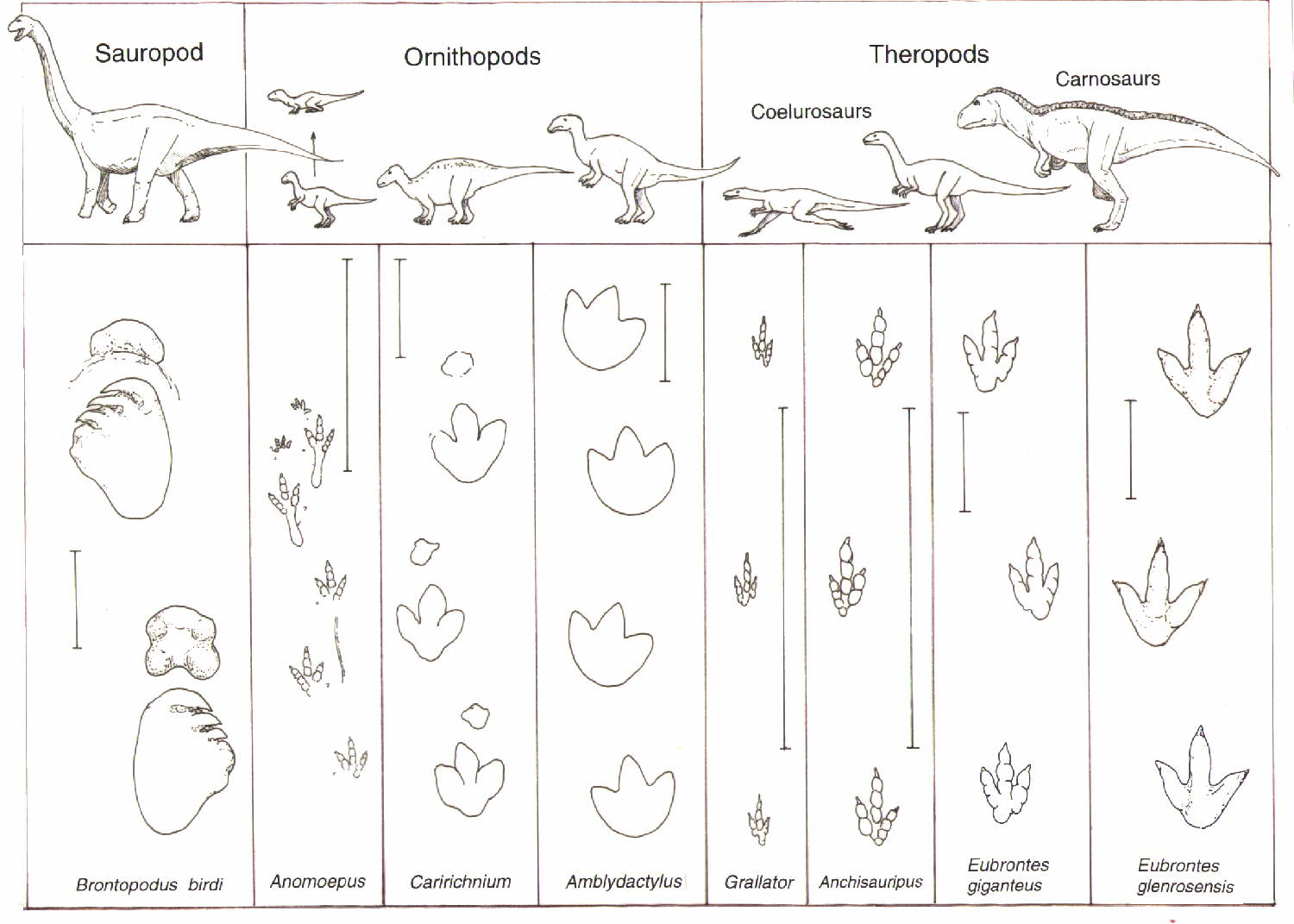
Sauropods of the Triassic: Rare, but Not Absent: Up until recently, many South African and Thai basal sauropods were interpreted as coming from the Late Triassic, but new stratigraphic analyses show they are from the earliest Early Jurassic. However, Lessemsaurus and Ingentia are pretty securely Late Triassic, and some track ways do strongly support the existence of stump-footed quadrupedal sauropod-style sauropodomorphs in this epoch.
That said, there is some disagreement on where the term "Sauropoda" should be placed on the phylogeny. In this course we are using "Saltasaurus and all taxa sharing a more recent common ancestor with it than with Melanorosaurus": in this case, the animals mentioned in the last few paragraphs are sauropods. An alternative definition--also commonly used--is "Saltasaurus, Vulcanodon, their concestor, and all of its descendants." Under this definition, Lessemsaurus, Ingentia, Antetonitrus, Gongxianosaurus, Pulanesaura, Isanosaurus, and the like would NOT be sauropods, and there would be no definite evidence of pre-Jurassic Sauropoda. (For what it is worth, we are using this "Vulcanodon+Saltasaurus" definition for Gravisauria, the major subgroup of Sauropoda.)
In Gravisauria there are four or more sacrals, and there is a transformation of the jaws and teeth:
Early sauropods are known from the southern continents (Gondwana) in the Late Triassic, and only show up in northern continents in the Early Jurassic. They seem to have been only rare components in dinosaurian communities until the Middle Jurassic.
EUSAUROPODA
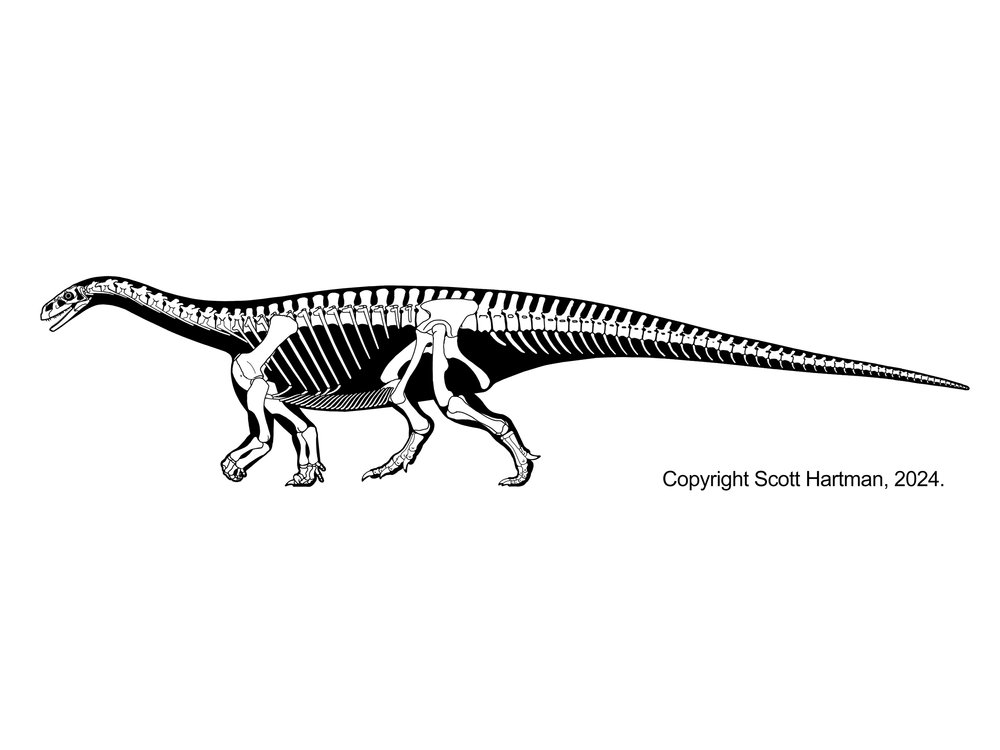
The sauropods of the Middle Jurassic onward comprise the Eusauropoda (true sauropod). Eusauropods differ from their ancestors in a number of ways:
There has been an ongoing debate over the position of the necks of eusauropods (well, long-necked sauropodomorphs in general, but especially for these guys). Computer studies of possible ranges of motion in sauropod necks suggested that they were basically horizontal. However, recent work on a diversity of living animals shows an upright curved neck in nearly all amniotes, so it is quite likely that a raised neck was the natural position for sauropods.
Recently discovered Spinophorosaurus of the Middle Jurassic of Niger is the most completely known primitive sauropod. It might be a eusauropod, or it might be the sister taxon to Eusauropoda. Its tail (like that of early Late Jurassic Chinese Shunosaurus) was interpreted as ending in a tail club: in fact, that of Spinophorosaurus was thought to be convergent in form on stegosaurian thagomizers. (New information indicates that the bones of the "tail club" may be misidentified.)
Eusauropods are the dominant herbivores worldwide in the Middle and Late Jurassic, and in at least some parts of the world all the way until the end of the Cretaceous. The oldest eusauropods, such as Bagualia and Patagosaurus actually date from the late Early Jurassic of South America. The best and most completely known early eusauropod is early Late Jurassic (long thought to be Middle Jurassic) Shunosaurus of China. Jobaria of the Middle Jurassic (once thought to be Early Cretaceous) of Africa has sometimes been considered a primitive member of Macronaria, but other analyses suggest it was a typical member of this early eusauropod radiation.
Several groups of primitive eusauropods are known. Recent studies suggest a paraphyletic series of "barapasaurs" like Barapasaurus of the Early Jurassic of India, Spinophorsaurus, Shunosaurus, and the Cetiosauridae, named after Cetiosaurus of the Middle Jurassic of England and possibly Patagosaurus of South America.
Different clades of eusauropods show different specializations towards reaching higher in trees. Several Late Jurassic-Early Cretaceous Asian sauropods evolved extra cervical vertebrae which were proportionately very long. Some of these sauropods (including Late Jurassic Omeisaurus and Mamenchisaurus) seem to form a clade, Mamenchisauridae. Some mamenchisaurids seemed to have reached truly tremendous size. Tonganosaurus may be an Early Jurassic representative of this clade; Wamweracaudia of Late Jurassic Africa may be a non-Asian example (most other examples are from China or Thailand).
One of the most successful groups of eusauropods is the Turiasauria. Some of these turiasaurs (such as Turiasaurus itself) evolved into tremendously large forms (rivaling various giant neosauropods). Initially known only from Late Jurassic European taxa, Turiasauria is now known to include taxa from the Middle Jurassic through the Early Cretaceous, and regions including North America (such as Moabosaurus), China, Madagascar, and Africa.
NEOSAUROPODA
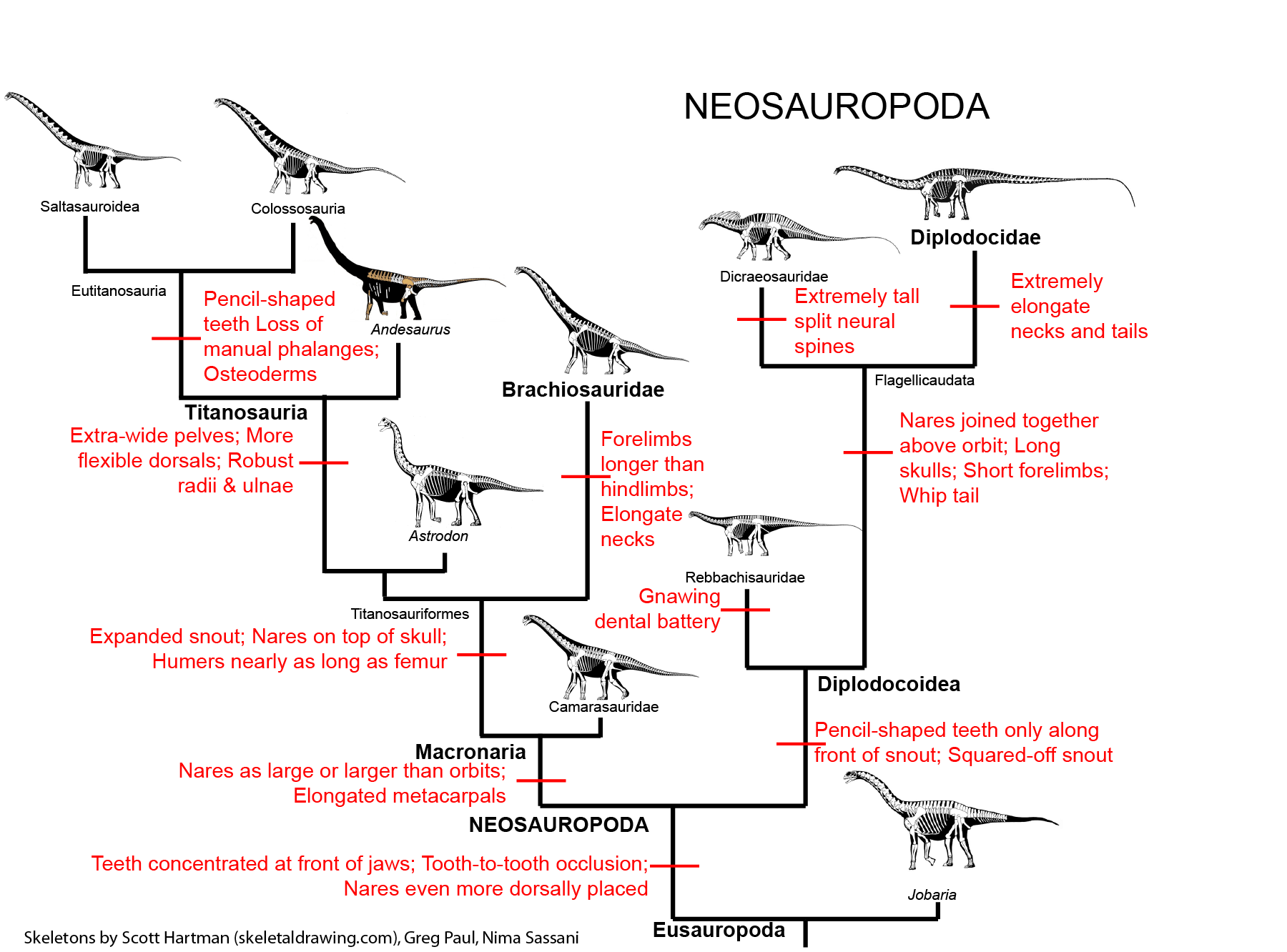
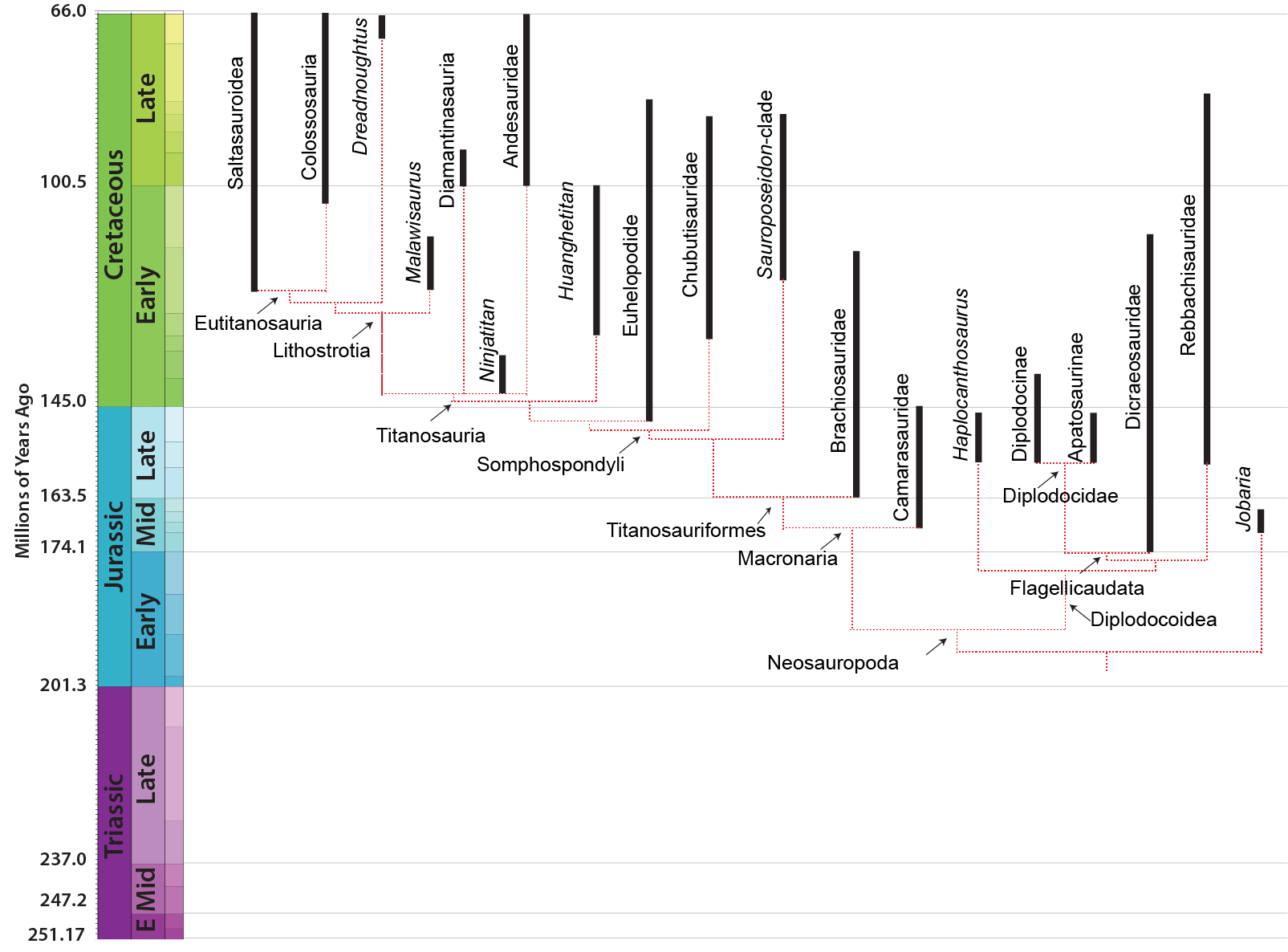
The dominant group of post-Middle Jurassic sauropods is the Neosauropoda (new sauropods). Their transformations from primitive eusauropods are:
Neosauropods have two major subdivisions: Diplodocoidea and Macronaria.
DIPLODOCOIDEA
This group differs from their ancestors by a number of traits, including:
The pencil-shaped teeth were used for raking or scraping leaves off of trees, or grazing on low vegetation. Recent (2018) discoveries of a baby Diplodocus shows teeth more like basal eusauropods or macronarians, for cropping up whole plants. So there was likely an ecological switch as diplodocoids got older.
The basalmost branch of diplodocoids currently known is the Late Jurassic North American genus Haplocanthosaurus. Unfortunately the skull is not yet known: its membership in this clade are based on details of the vertebrae and limb bones.
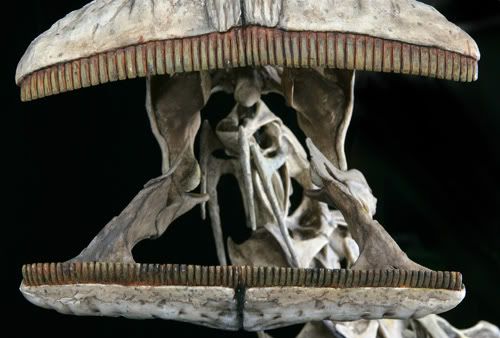
The first major branch of diplodocoids is the truly bizarre Rebbachisauridae, currently known from the Cretaceous, mostly from Gondwana. (However, a lost specimen which has been named "Maraapunisaurus fragillimus" is from the Late Jurassic of Colorado: it appears to be a primitive rebbachisaurid of tremendous size (~30-32 m long)). Rebbachisaurids are best known from Nigersaurus from the late Early Cretaceous of northern Africa. At least Nigersaurus was "little" (as sauropods go, that is: only the size of an Indian elephant!). Their skulls are highly transformed, most importantly with:
Unlike the slicing dental battery of ceratopsids or the grinding dental battery of hadrosaurids, the rebbachisaurid dental battery is at the front end of the jaw. The rate of replacement of the teeth was very high (one tooth/month; about twice as fast as hadrosaurids).
Other rebbachisaurids include Rebbachisaurus and Tatouinea of northern Africa and Limaysaurus, Rayososaurus, Cathartesaura, and Nopscaspondylus of South America, and Histriasaurus and Demandasaurus of Europe. In recent years rebbachisaurids have been found from all over the world.
The remaining diplodocoids are the Flagellicaudata (whip tails). These form a major part of the sauropod communities of the Late Jurassic and early Early Cretaceous. Their specializations include:
There are two major divisions of the flagellicaudates. The Dicraeosauridae were smaller (only 10 m or less for most), and had:
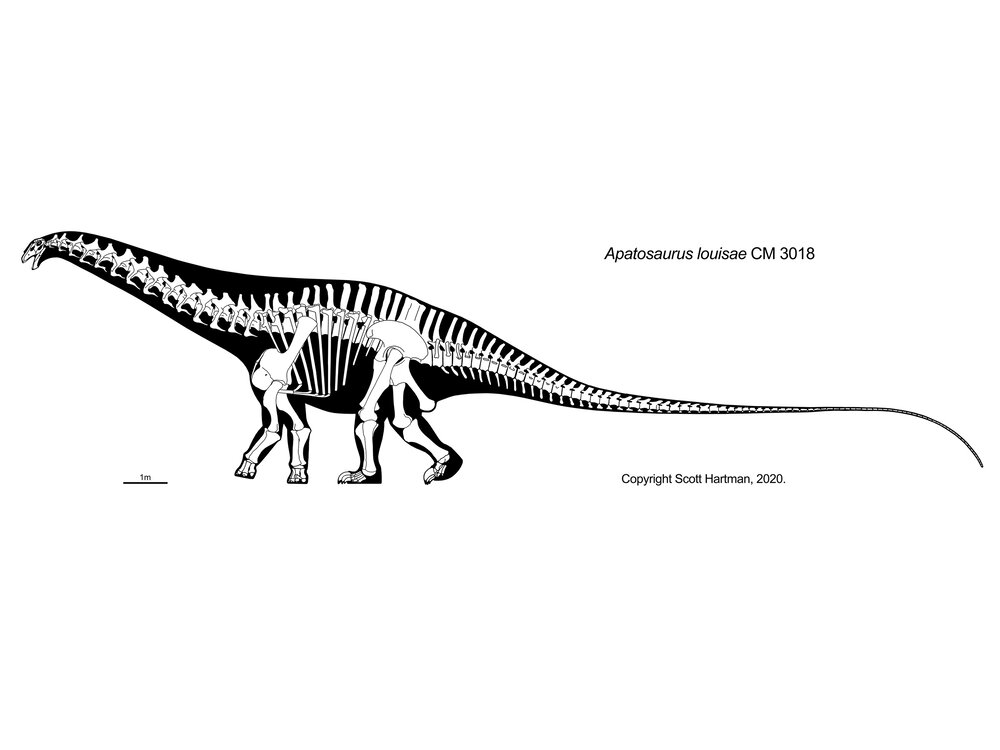
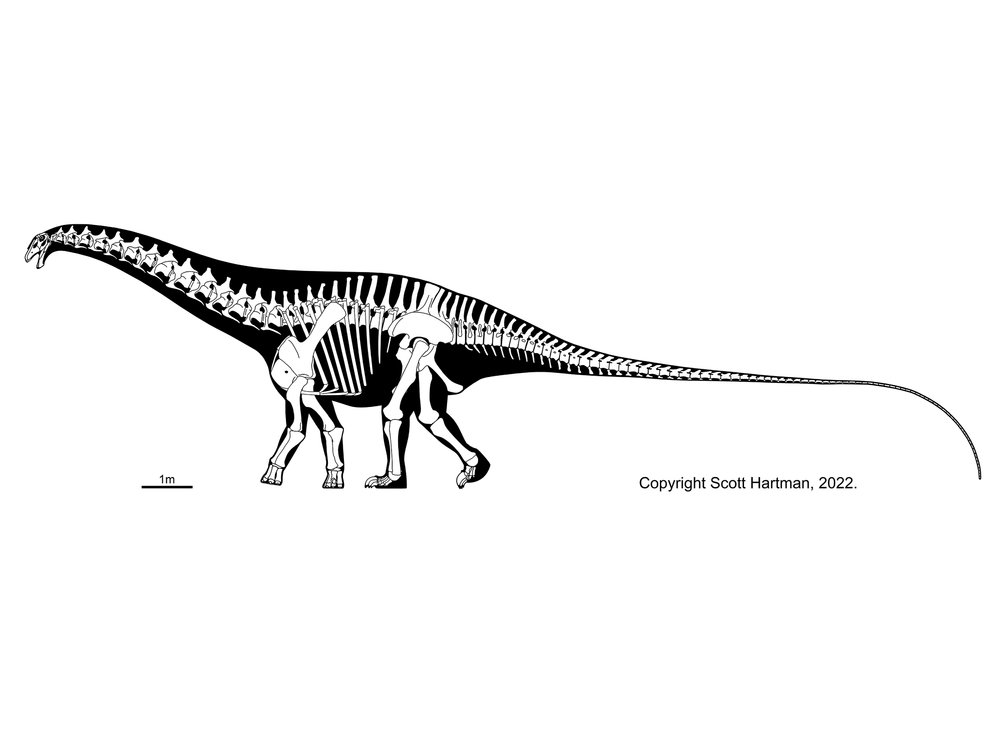
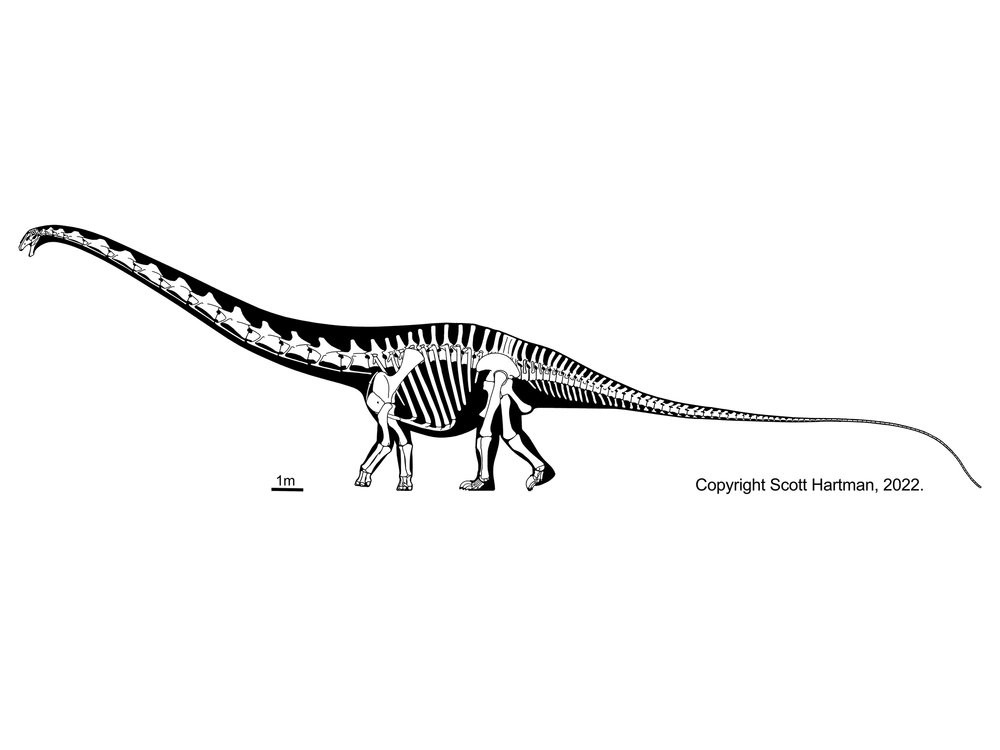
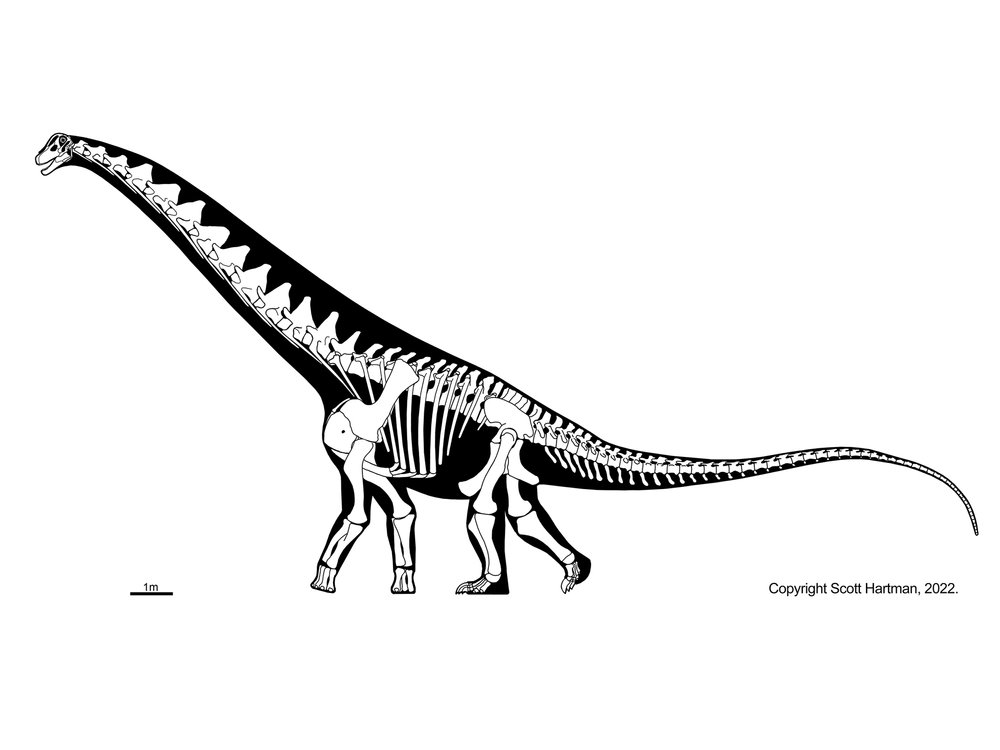
More famous are the Diplodocidae. These are very large sauropods (over 30 m in the massively-built apatosaurines Apatosaurus and Brontosaurus, and more slender Diplodocus [the individual formerly considered Seismosaurus], Galeamopus, and Supersaurus. They are characterized by the following specializations:
Diplodocids (and other diplodocoids?) may have been able to rear up on their hindlimbs to feed even higher in trees than they could using their long necks. Alternatively, they may have been low-browsing/grazing specialists: we'll come back to this question in a later lecture. At present, almost all diplodocids are known only from the Late Jurassic (Leinkupal of Early Cretaceous Argentina being the exception). In western North America they were tremendously diverse: at least six genera were present at one time.
Thunderstruck: Diplodocoid Whiptails: The tails of diplodocids (and possibly other diplodocoids, which are currently not well known) are muscular close to the hips, but are very, very, VERY long and slender for their posterior half. It has long been suggested that it was used defensively, to strike against an attacking theropod just like a modern monitor lizard or iguana does. (In fact, the Smithsonian's specimen of Allosaurus is damaged along its left dentary, scapula, and ribs, perhaps the result of a clobbering from a diplodocid!). Some have gone even further, and suggested that the whip was specifically used like a bullwhip: that is, that its tip went supersonic, forming a powerful crack sound. It remains debated to what degree either such use would damage the bones and (very limited) flesh of the posterior tail; furthermore, the latest studies show that they couldn't actually make sonic-boom speeds. Nevertheless, the whapping subsonic tail of large lizards today are effective defensive weapons, so the multi-ton appendages of diplodocids would have been devastating even at non-cracking speeds.
MACRONARIA
The other major branch of neosauropods is the Macronaria (big noses). They range from the Middle Jurassic to the very end of the Cretaceous. Their changes from ancestral sauropods include:
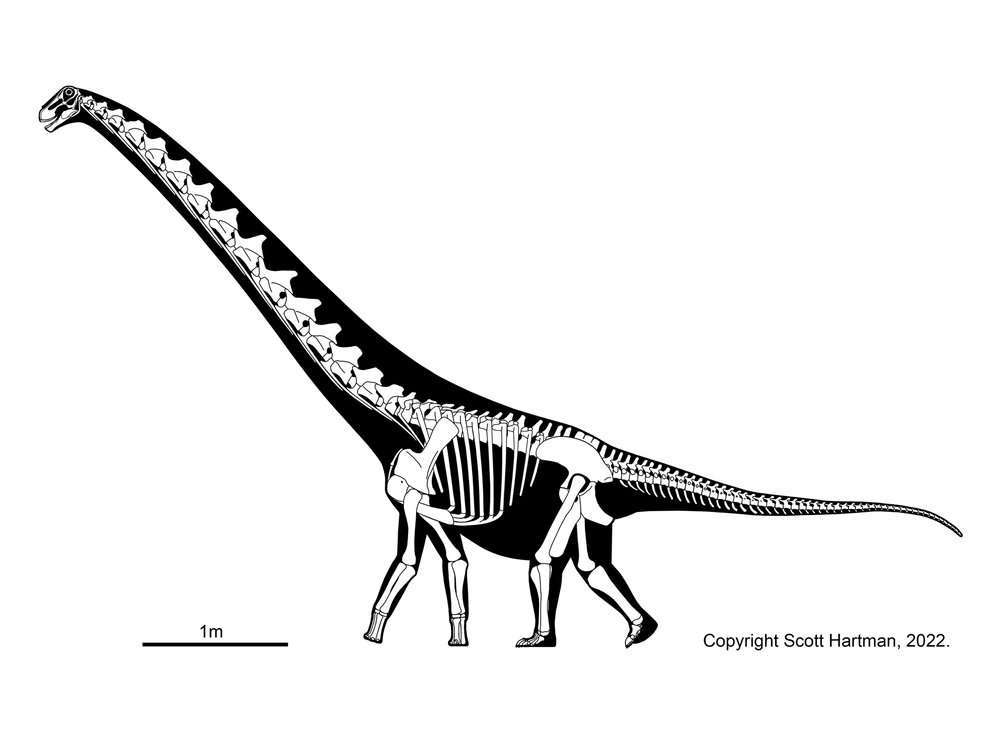
Primitive macronarians include Camarasaurus of the Late Jurassic of western North America (and possibly elsewhere); late Middle Jurassic Tehuelchesaurus of Argentina; Lourinhasaurus of Portugal; and others. The first three may form a clade Camarasauridae.
(However, a note for the future: some big on-going phylogenetic analyses threaten to shake up the family tree. It may be that Camarasaurus will fall outside of a clade comprised of diplodocoids, brachiosaurids and their kin, and titanosaurs. Stay tuned.)
The more advanced macronarians (the Titanosauriformes) are specialized by:
Astrodon (State Dinosaur of Maryland; Early Cretaceous of Maryland, and possibly Texas) is a titanosauriform, as is as is gigantic Sauroposeidon (which includes specimens previously called "Paluxysaurus" of Early Cretaceous western North America. Three major clades of titanosauriform include the Brachiosauridae, the Euhelopodidae, and Titanosauria.
Brachiosauridae (Middle Jurassic to Early Cretaceous) are very distinctive. They are characterized by:
The result is that brachiosaurids were built "up hill", making them some of the tallest animals that ever lived. Best known are Late Jurassic Brachiosaurus of North America, Giraffatitan of Africa, Lusotitan and Europasaurus of the Late Jurassic of Europe, and Early Cretaceous Abydosaurus, Venenosaurus, and Cedarosaurus of western North America. Early Late Jurassic Vouivria of Europe and Middle Jurassic Atlasaurus of North Africa appear to be the oldest and most primitive brachiosaurids.
Astrodon and Sauroposeidon are sometimes considered to be brachiosaurids. Alternatively, some studies find the "brachiosaurids" as a paraphyletic series with regards to euhelopodids + titanosaurs. Some recent studies find that these, plus several other Early Cretaceous North American forms and possibly even the Late Cretaceous Argentine Malarguesaurus form a clade.
Chubutisaurus of late Early Cretaceous Argentina (once considered part of Titanosauria) may form a clade with numerous South American, Asian, European, African, and North American taxa of mid-Cretaceous to Late Cretaceous age.
The Euhelopodidae is a clade of ultra-long-necked Cretaceous titanosauriforms. Most known taxa are from Asia, but Late Jurassic African Australodocus may be a primitive euhelopodid (alternatively, it may be a brachiosaurid, more consistent with its age and location). Euhelopodids include (among others) Early Cretaceous Euhelopus, Qiaowanlong, Phuwiangosaurus, and early Late Cretaceous Erketu. Daxiatitan and Ruyangosaurus of the early Late Cretaceous of China may both be euhelopodids: the latter is one of the contenders of "largest dinosaurs of all time", with a 2.35 m (7.7 foot) tall femur. (However, some traits of the vertebrae of Ruyangosaurus suggest it is a titanosaur, perhaps an Andesaurus-relative, perhaps a colossosaur.) Huabeisaurus may be a later Late Cretaceous euhelopodid; alternatively, it might be an andesauroid. Gannansaurus and Jiangxiatitan (from near the end of the Cretaceous) are possibly the youngest known euhelopodids, but their exact phylogenetic position isn't well known.
In a recent (2024) study, the species of the giant Early Cretaceous Chinese Huanghetitan might be the sister group to Titanosauria.
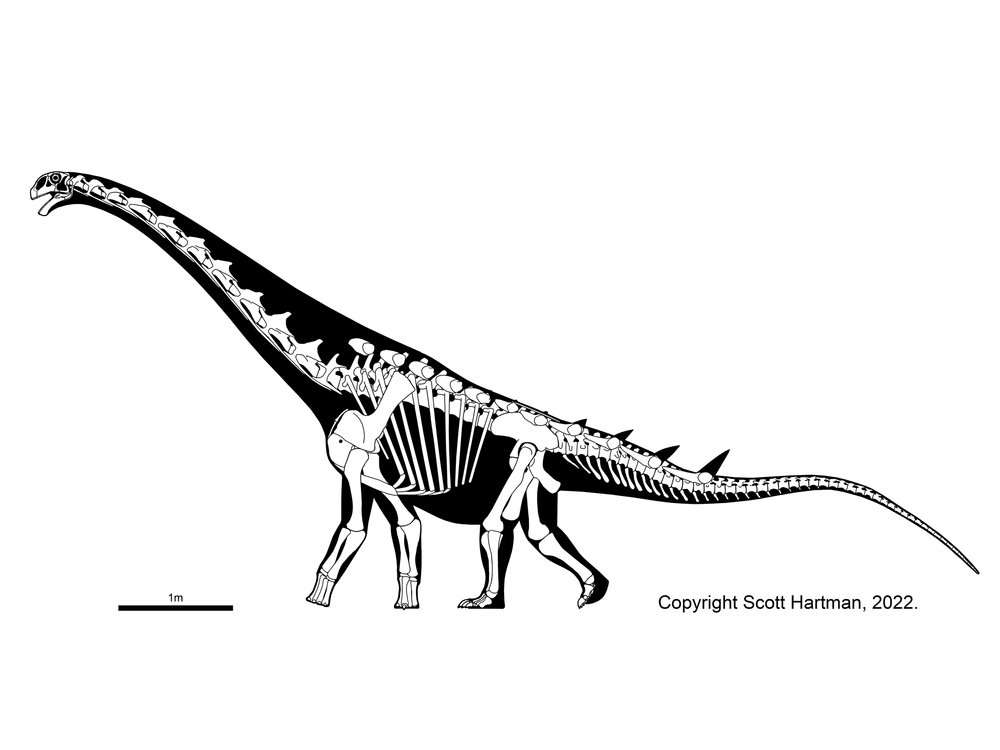
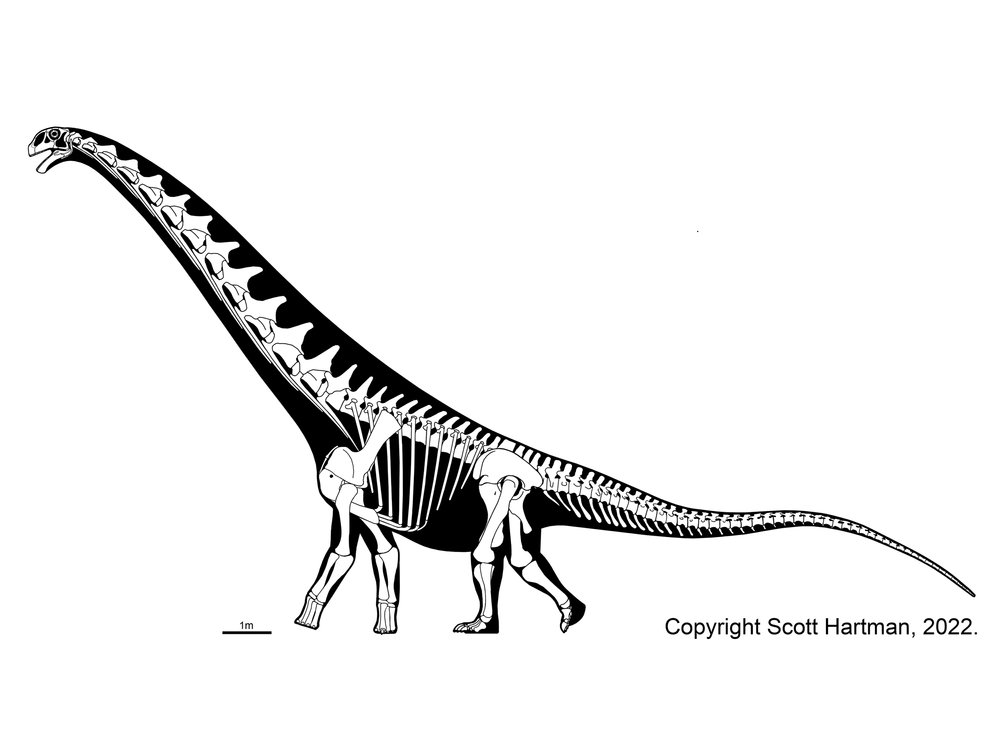
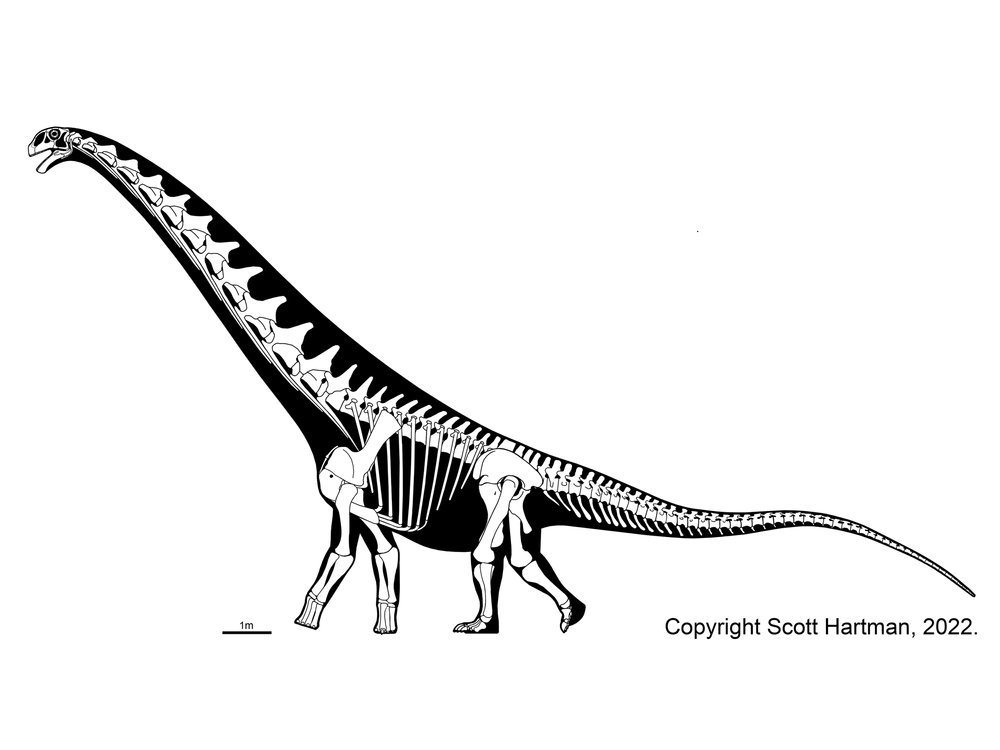
Titanosauria (Cretaceous) were the dominant group of Cretaceous sauropods in most parts of the world. They were very diverse. The different groups share:
Our understanding of titanosaur phylogeny is, quite frankly, a mess. There are a lot of phylogenetic analysis using quite a number of taxa (titanosaurs represent nearly 1/2 of all sauropod genera!), but very little in the way of consensus between them. We are using a grossly simplified version here, based on a combination of several (conflicting!) recent studies. The earliest branching member of Titanosauria is Andesaurus and its kin, while newly (2021) named Ninjatitan is the oldest of all definite titanosaurs: its position within Titanosauria is very poorly constrained at present.
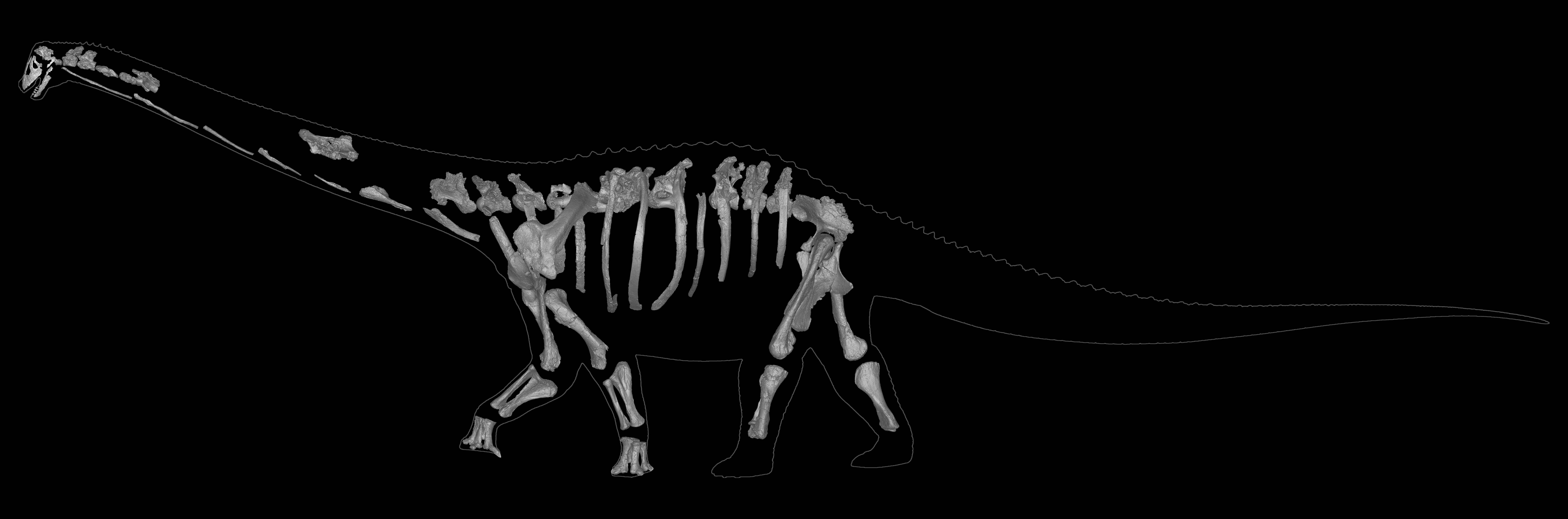
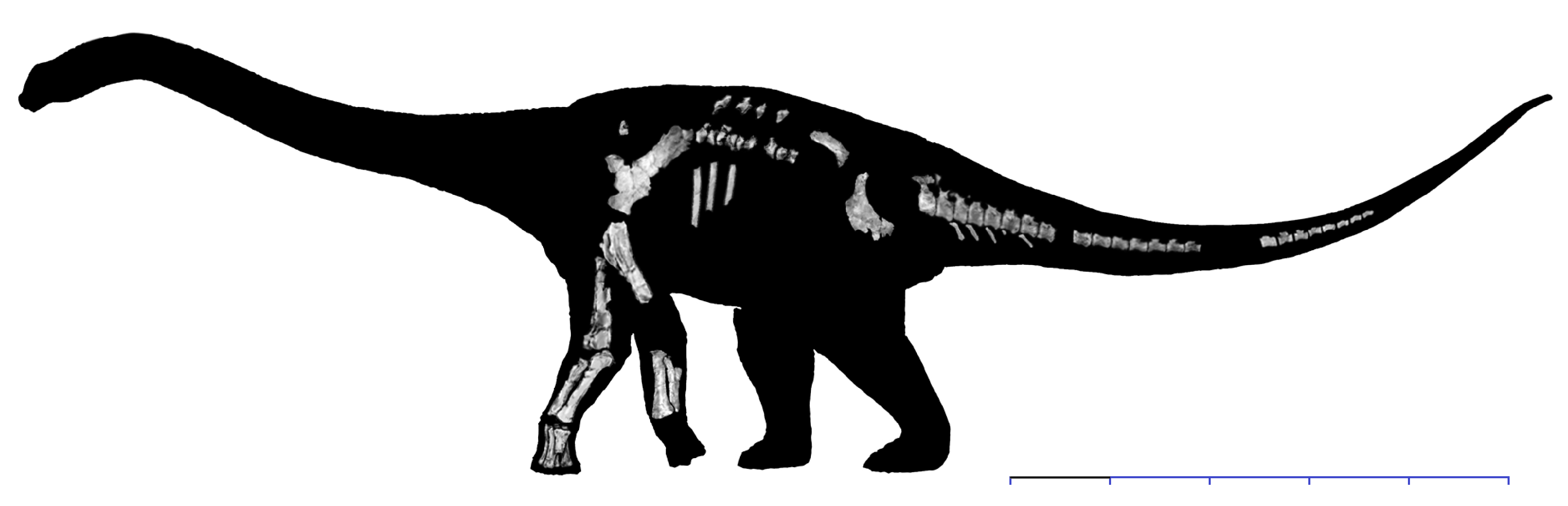
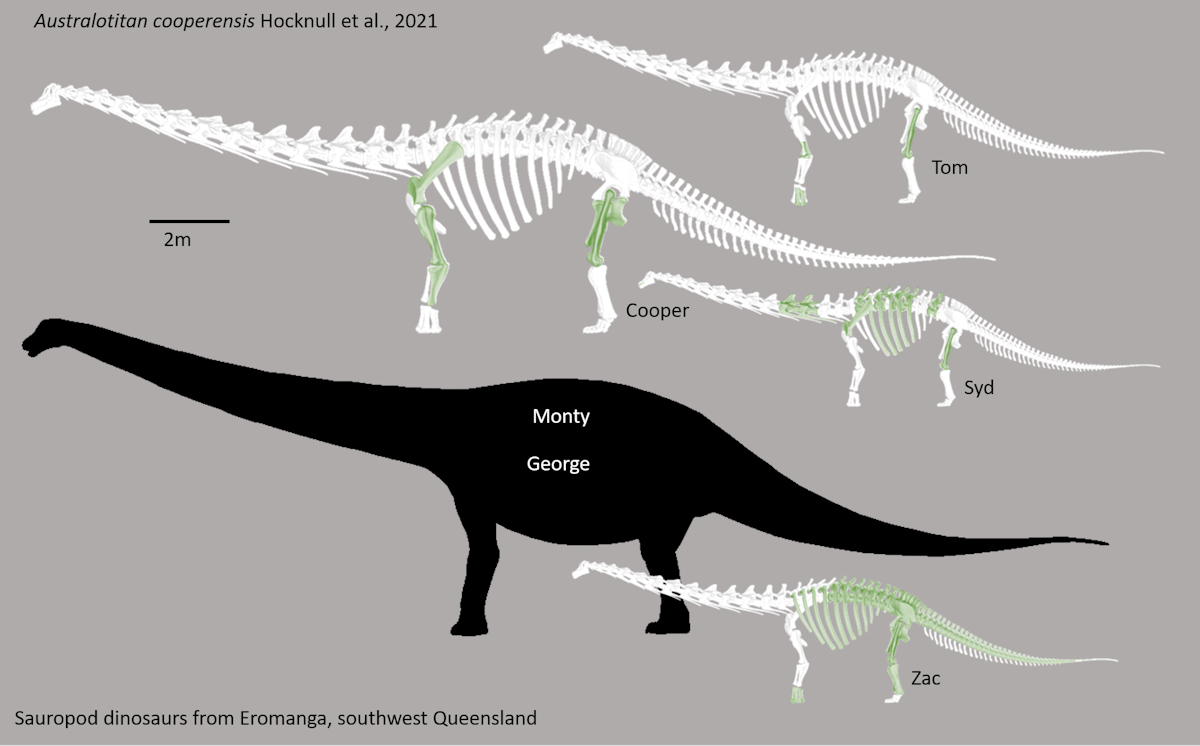
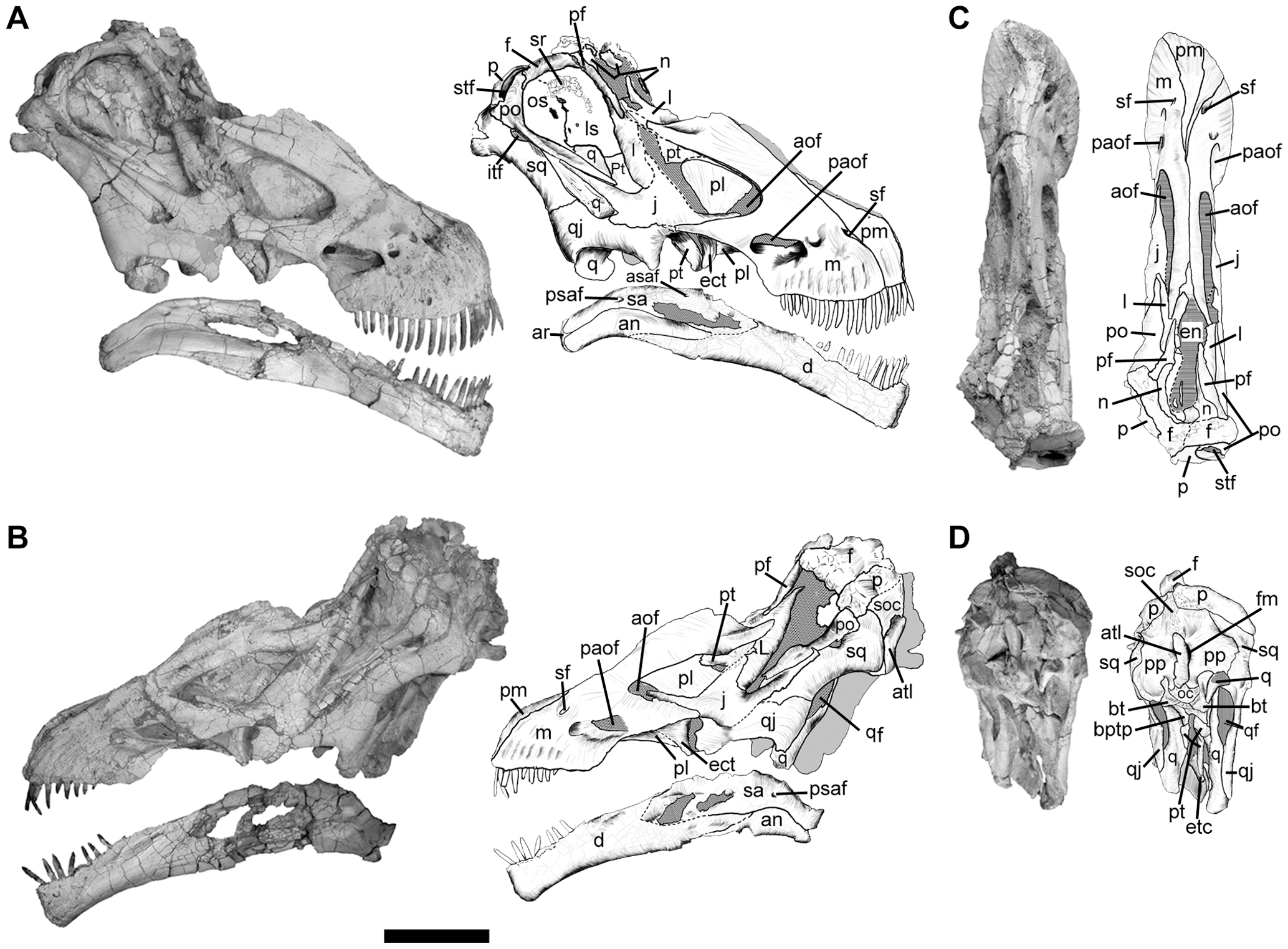
The Diamantinasauria of the Early Late Cretaceous are a newly-recognized group of early-branching titanosaurs (or just outside Titanosauria) (or a subgroup of the derived Saltasauroidea: I SAID the phylogeny is a mess...). Two of these taxa, South American Sarmientosaurus and Australian Diamantinasaurus are known from nearly complete skulls. Wintonotitan (also from the late Early Cretaceous of Australia) also seems to be a diamantinasaur. A specimen of Diamantinasaurus once considered its own taxon ("Australotitan") is nearly as big as the Argentine giant titanosaurs.
Malawisaurus of the Early Cretaceous of eastern Africa is the oldest known member of Lithostrotia ("stone skins"), the clade of titanosaurs known to have osteoderms. (However, it might be that osteoderms are found even more basally within Titanosauria). This "armor" may not have been for protection, however; these osteoderms seem to have been sparse on the backs of the dinosaurs, separated by lots of skin. Furthermore the bone texture is quite "spongy". One possibility is that these osteoderms really functioned as a mineral store that the body could draw upon; for instance, during harsh seasons or when females were producing eggs.
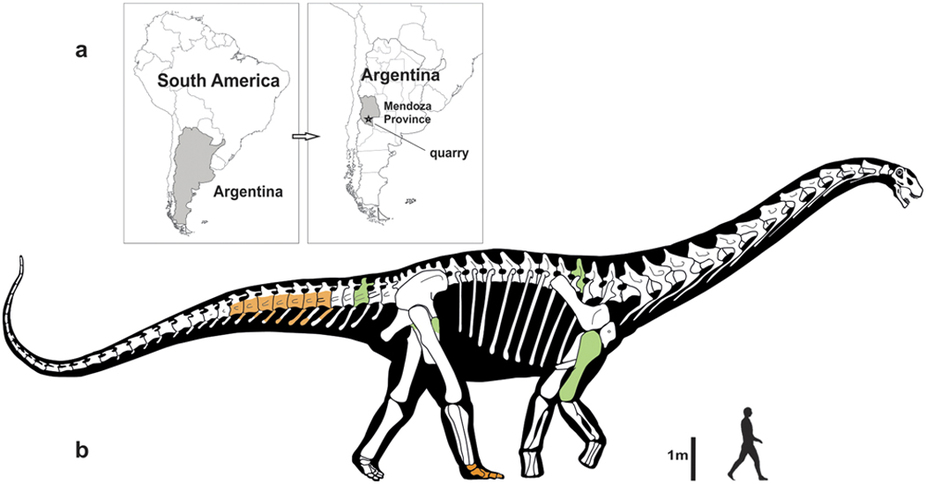
Gigantic Dreadnoughtus of the Late Cretaceous of Argentina might be the sister group to the Eutitanosauria, but some early studies found it to be outside the lithostrotians or (alternatively) as a primitive saltasauroid.
In addition to these primitive taxa, the titanosaurs include a more derived clade Eutitanosauria, which is further modified by the evolution of:
Relationships among the eutitanosaurs are still being worked out with no strong resolution at present. A new (2022) attempt to simplify the taxonomy (if not the various internal relationships) splits Eutitanosauria into Colossosauria and Saltosauroidea (the version shown here), but the memberships of these clades vary and the arrangement of other eutitanosaurs come out differently in various analyses.
Colossosauria is a robust clade that seems to show up in many recent studies. It includes two strong subclades: Rinconsauria (Rinconsaurus and Muyelensaurus) and the Lognkosauria (including huge Notocolossus, stupendous Futalognkosaurus, tremendous Patagotitan, and its sister taxon, the even more gigantic 90-100 ton Argentinosaurus (perhaps the largest dinosaur of all)). Nearly all the very largest titanosaurs (indeed, dinosaurs) are lognkosaur colossosaurs, but not all. For instance, Alamosaurus (the last North American sauropod) seems to be a saltasauroid. The position of tremendous Argyrosaurus (and many other titanosaurs) remains unresolved.
Saltasauroids include most of the last of the titanosaurs. Some (such as Alamosaurus) are true giants; others are smaller, such as Saltasaurus itself (the first titanosaur known with osteoderms) and Rapetosaurus; broad-snouted forms such as Nemegtosaurus (known only from a head); broad-bodied Opisthocoelicaudia (known only from a body, and possibly the same animal as Nemegtosaurus); and many others. The phylogenetic position of Early Cretaceous Brazilian Tapuiasaurus is uncertain: it was originally considered in the saltasauroid lineage, but may be an earlier branch of titanosaur. Tapuiasaurus's skull is very similar to that known from the saltasaurid Nemegetosaurus and what is known of the skulls of aeolosauroids, and to that of diamantinasaurs.
The extreme members of the broad snouted saltasauroids are the Antarctosauridae, a clade of late Late Cretaceous South American forms whose jaws are convergent on those of rebbachisaurids. One of the most completely known examples of this group is Inawentu.
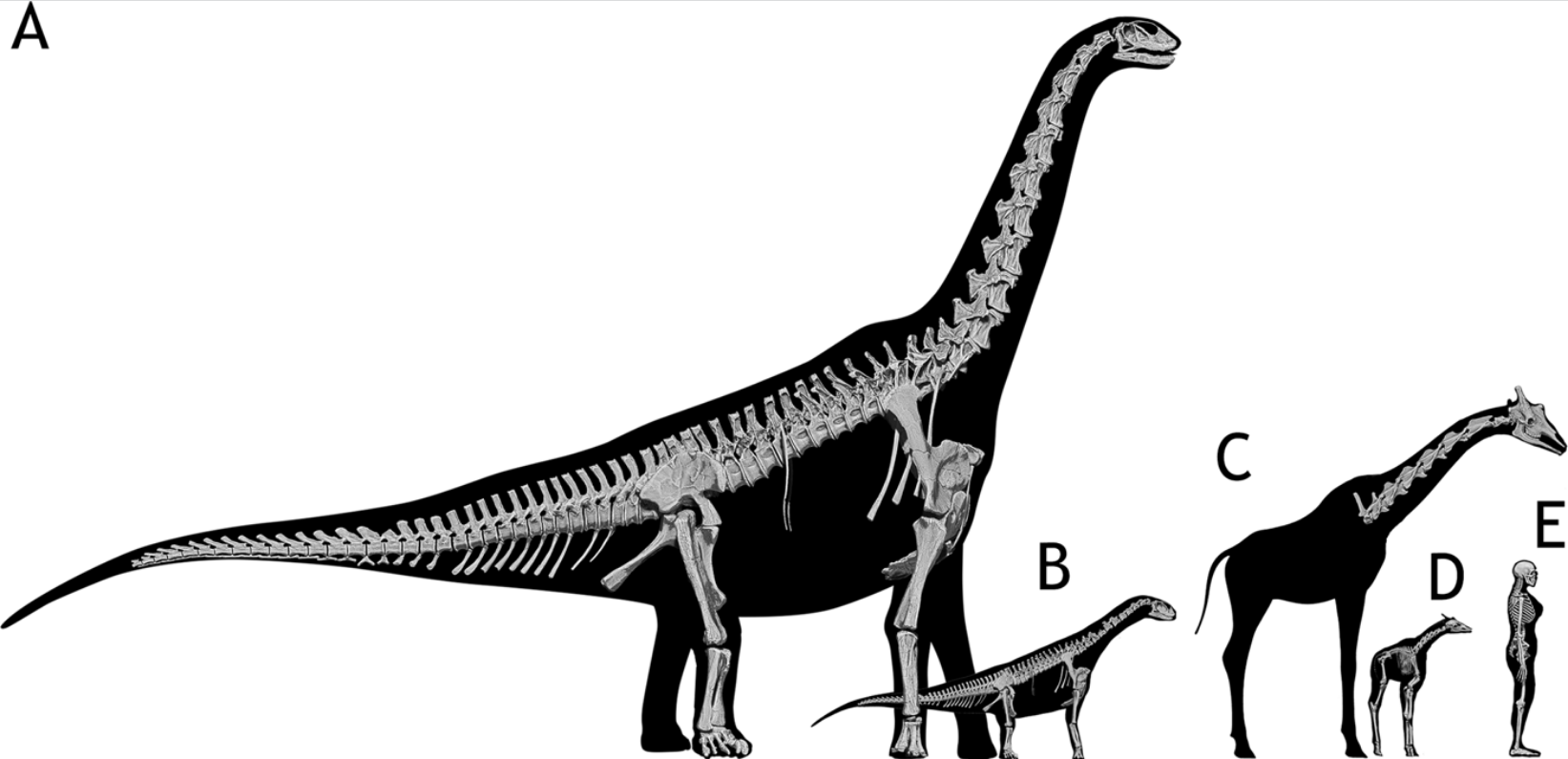
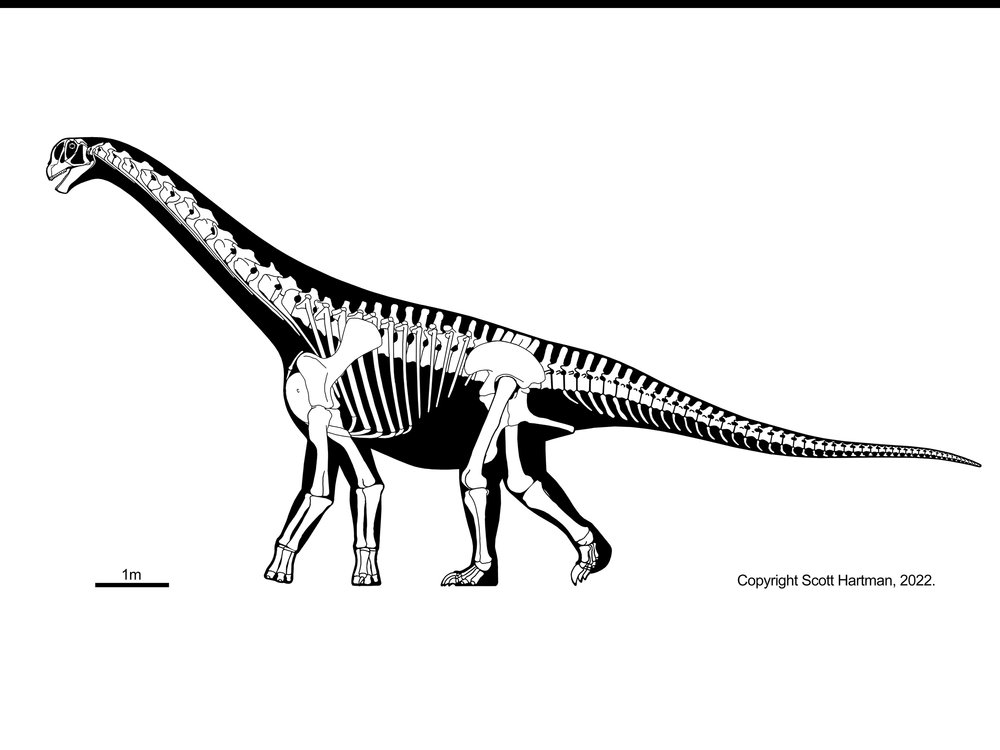
Titanosaurs are the main group of herbivorous dinosaur in the southern continents (and parts of Europe) until the end of the Cretaceous, and were found on all continents. Titanosaurs include some truly tremendous forms: Futalognkosaurus, Argentinosaurus, Puertasaurus, Alamosaurus, and Patagotitan all seem to reach just about the same size: masses of perhaps 100 tons or so (depending on the methodology used; possibly around half that under other estimates), and lengths much more than 30 m. Other clades of eusauropod have multiple examples nearly as large. These dinosaurs are still smaller (less massive) than the modern blue whale, but otherwise exceed any living species of animal in mass. We will explore later on HOW it was that dinosaurs could be able to reach this supremely gigantic sizes.
Diets for Giants: A question many have pondered is how sauropods managed to get enough food for their sustenance in the course of a day. A very recent study examined the nutritional value of extant plants of Mesozoic style when grown under range of possible Mesozoic atmospheric conditions and compared them to likely metabolic rates of sauropods. They found that even though the nutritional value of plants doesn't vary under these different conditions as much as was once thought, but that sauropods could easily acquire enough food every day to grow and survive.
Sauropods: the Aquatic Dinosaurs?: In the late 19th through the mid-20th Century, sauropods were often portrayed in both scientific and popular literature as "the aquatic dinosaurs." Some imagined that because they reached sizes far surpassing the largest land animal today, they simply could not have been land animals at all! Instead, sauropods were envisioned to have been swamp- and lake-dwellers, using the water to buoy up their enormous masses. One school of thought suggested that their long necks and dorsally-placed nares was used as snorkels, with just the tops of their heads occasionally breaking the surface of the water to breath, like a dinosaurian blowhole.
However, others pointed out the flaws with this model. For one thing, the narrow feet of sauropods were the opposite of what you would want as an adaptation for wet mud. The snorkel hypothesis wouldn't work: the water pressure on the lower neck and lungs would be too great to allow them to expand to let air in: in fact, it would be a great way for a dinosaur to try to kill itself! As we now recognize, the nostrils themselves are NOT dorsally placed, but are instead at the anterior end of the snout. And sauropod fossils are known from a wide variety of habitats: dry uplands, forests, river systems, and yes, even lake shores, sea shores, and swamps. Surely some of them lived at least part of their time in wet environments, but the group as a whole was not adapted to it.
EVOLUTIONARY PATTERNS IN SAUROPODOMORPHA
Feeding adaptation transformations:
Gigantism:
Locomotory changes:
Heterochrony and the origin of Sauropoda:
Niche partitioning:
Many dinosaur communities had multiple sauropod species at the same time. How could they support this diversity? Niche partitioning: each clade (and even each species) specialized in feeding in a slightly different manner. For example, in Late Jurassic communities there were ground-feeding dicraeosaurids, mid-level feeding primitive eusauropods and primitive macronarians, high feeding brachiosaurids, mamenchisaurids, and diplodocoids (which could feed exceptionally high if they could actually rear on their hindlegs)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}