


Key Points:
•Theropods include the majority of carnivorous dinosaurs (although various subgroups evolved towards herbivory, omnivory, insectivory, etc.). They remained entirely obligate bipeds.
•Among the diagnostic characteristics are an intramandibular joint, a promaxillary fenestra, a furcula (wish bone), and a functionally tridactyl pes.
•Early theropods (coelophysoids) were only minor predators in their ecosystem; however, the Triassic-Jurassic extinction eliminated their competition, and from beginning of the Jurassic until the end of the Cretaceous theropods were the dominant group of terrestrial predators.
•Some major groups include: primitive Coelophysidae and Dilophosauridae; Ceratosauria, with reduced hand function; Megalosauroidea, including the fish-eating Spinosauridae; Carnosauria, which dominated the apex predator niche for most communities from the Middle Jurassic until the early Late Cretaceous; and the hugely diverse Coelurosauria.
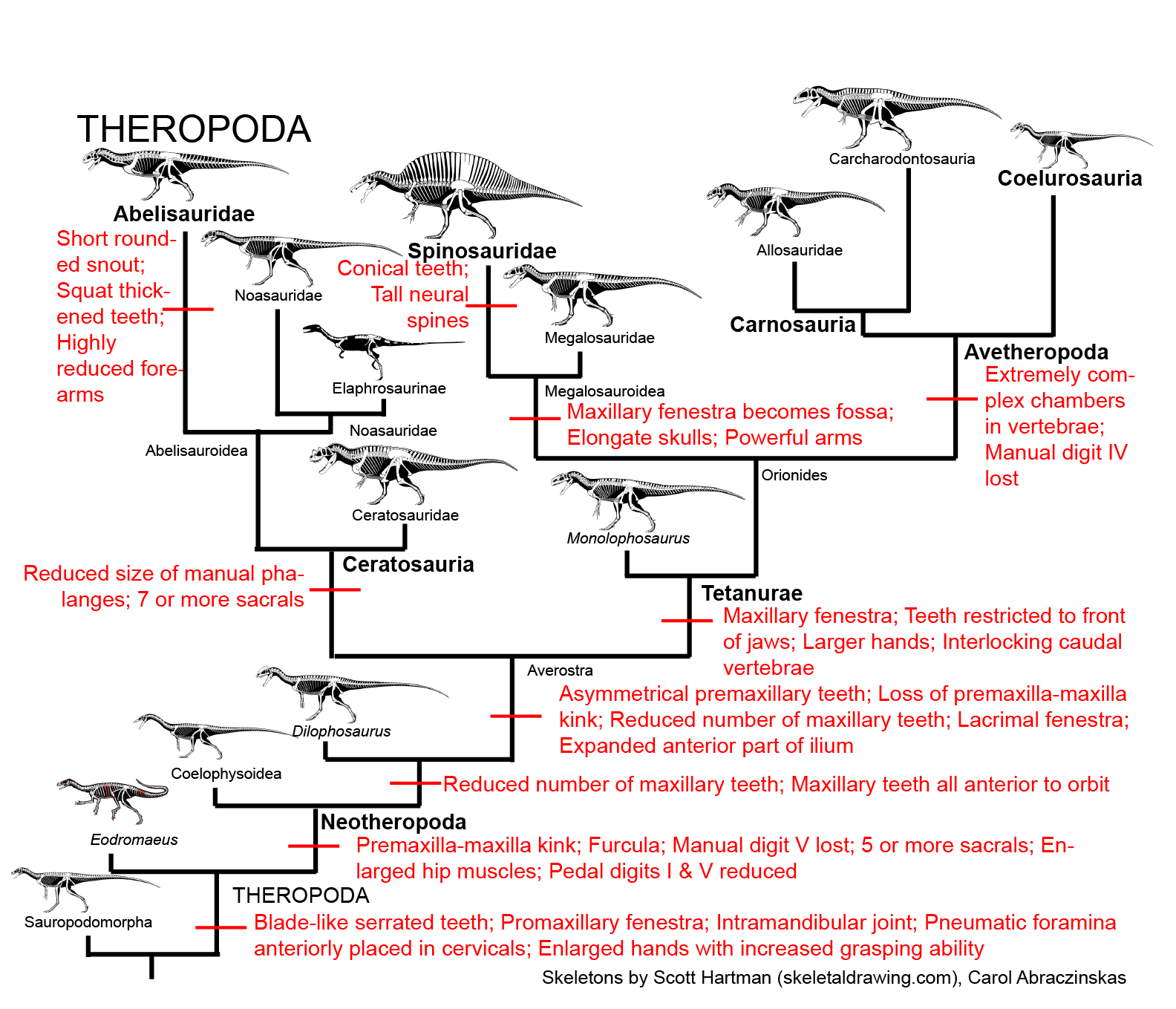
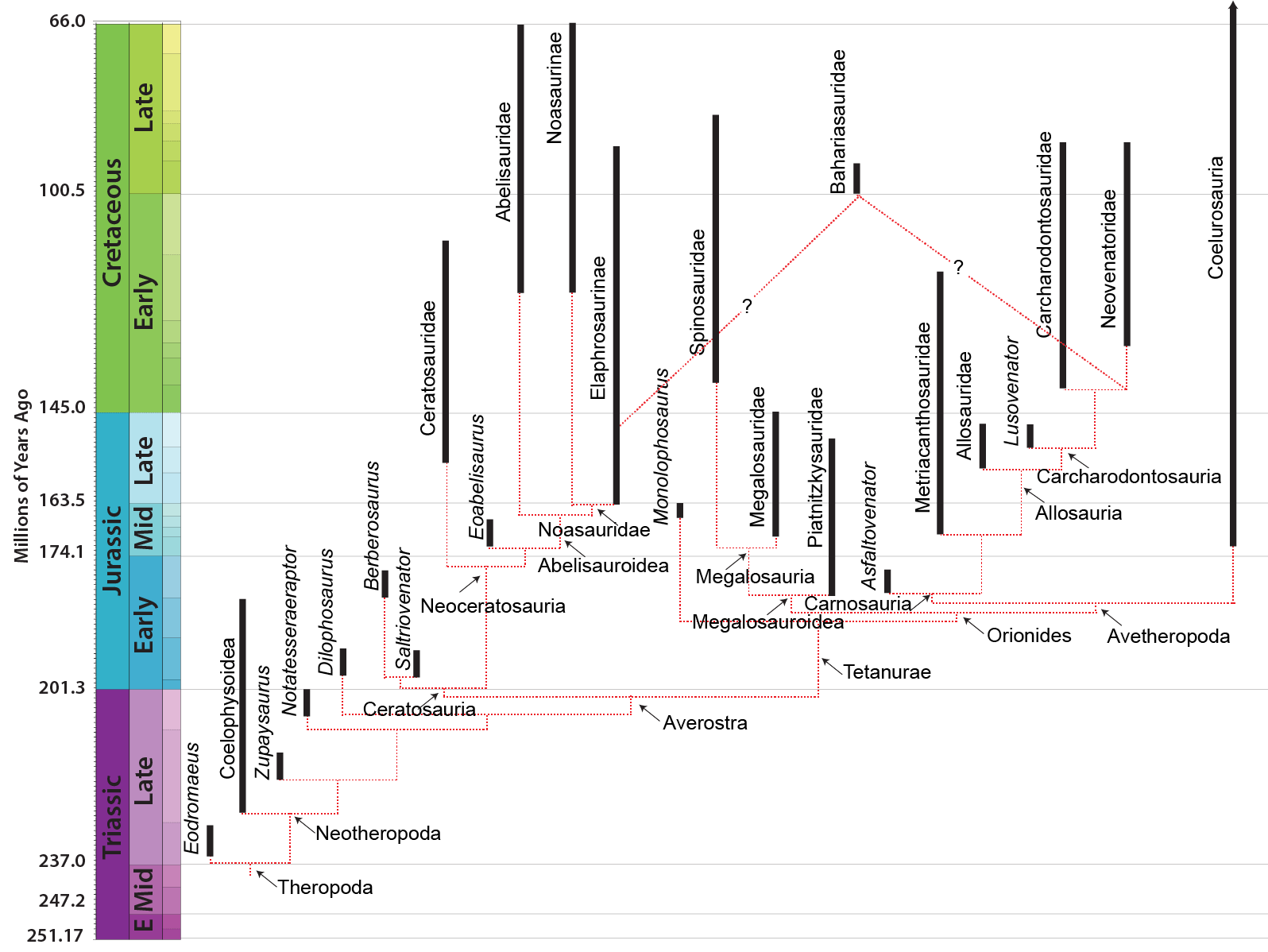
MAJOR GROUPS OF THEROPODS
BASAL THEROPODS
The traits uniting Theropoda seem to include:
As discussed before, there are number of Triassic dinosaurs which might or might not belong to Theropoda: herrerasaurs, Eodromaeus, Tawa, Chindesaurus, and others. The remaining (definite) theropod taxa (coelophysids, dilophosaurids, ceratosaurs, and tetanurines) form a clade called Neotheropoda, although some have called this group "Eutheropoda" ("true theropods") and restricted "Neotheropoda" to the group called "Averostra" below. Theropods are present in the Late Triassic; are the dominant group of terrestrial carnivores throughout the entire Jurassic and Cretaceous; learned how to fly; had some members survive the great extinction; and are still with us today. In this lecture, though, we concentrate on the basal members of the theropod clade.
For this class, we will go with one of the latest studies, which regards herrerasaurids, Tawa, Chindesaurus, and Daemonosaurus are not theropods, but form the clade Herrerasauria outside the sauropodomorph-theropod clade Eusaurischia. However, little 1.2 m long Eodromaeus and newly-discovered (named in October 2025) Anteavis of the early Late Triassic Epoch of Argentina do seem to be the oldest and basal-most members of Theropoda proper.
Exclusive of the Triassic basal saurischians that might be theropods (which have some of the traits mentioned above), definite theropods ("neotheropods") show a number of specializations relative to other saurischians:
COELOPHYSOIDEA & "DILOPHOSAURS": BASAL NEOTHEROPODS OF THE TRIASSIC AND EARLY JURASSIC
There were two major clades of advanced Middle Jurassic and younger theropods: Ceratosauria and Tetanurae. Additionally, there are various primitive branches of the Late Triassic and Early Cretaceous. Some studies put these into two main clusters: Coelophysidae and Dilophosauridae. During the late 20th Century (and some early 21st Century studies), dilophosaurids, coelophysids, and the intermediate forms were collectively considered a clade "Coelophysoidea", and this whole grouping was found to be closer to Ceratosauria than either were to tetanurines; however, this course follows newer analyses that place Ceratosauria and Tetanurae in a clade (Averostra) exclusive of Coelophysoidea, with Dilophosauridae (and some other taxa) as intermediate between coelophysoids and avetrostrans. Collectively, we'll call celophysoids, dilophosaurids, and other non-averostran neotheropods "basal neotheropods" for now.
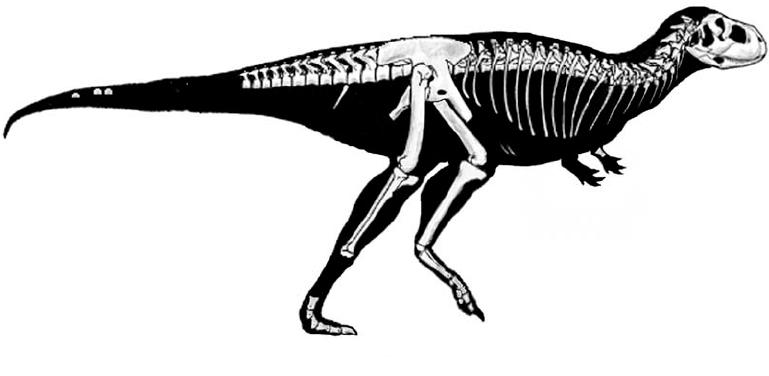
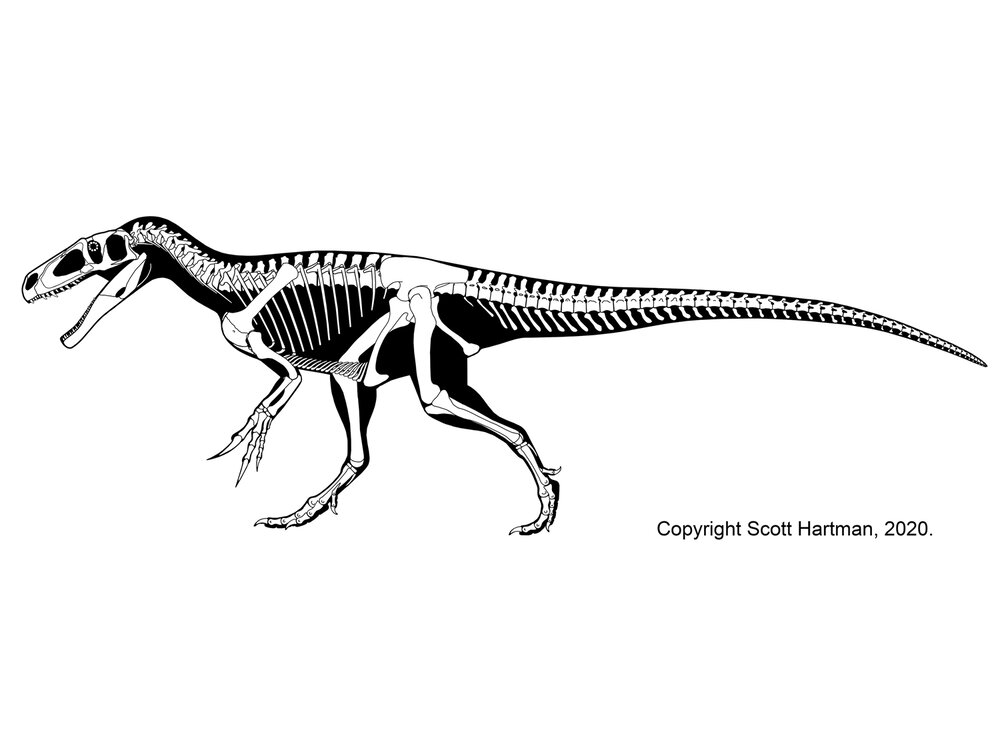
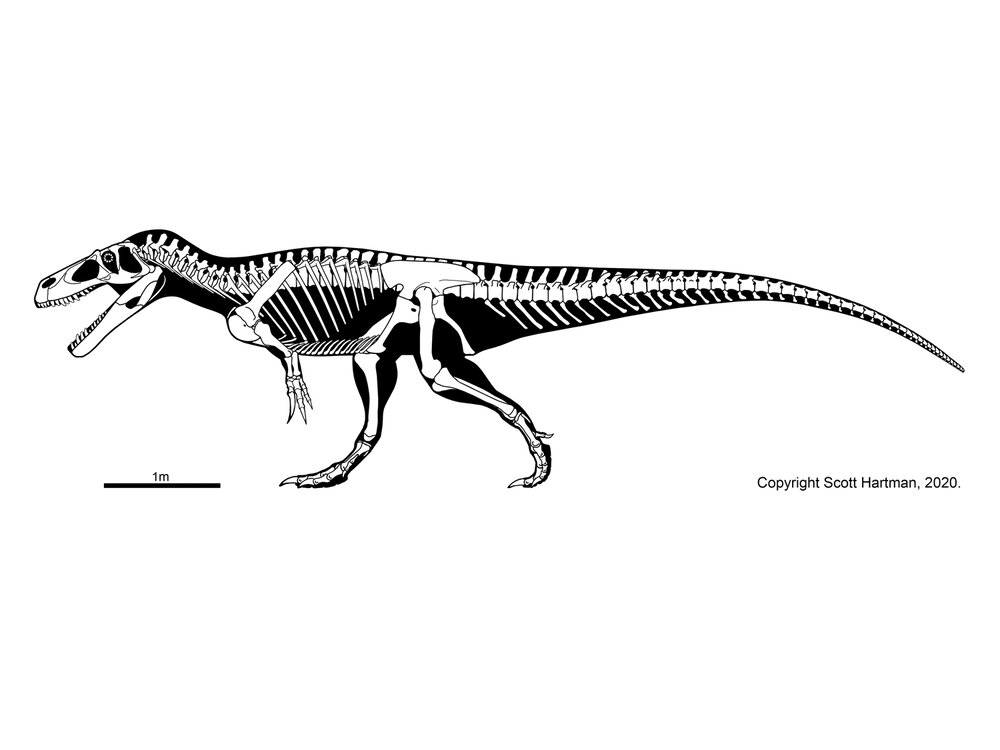
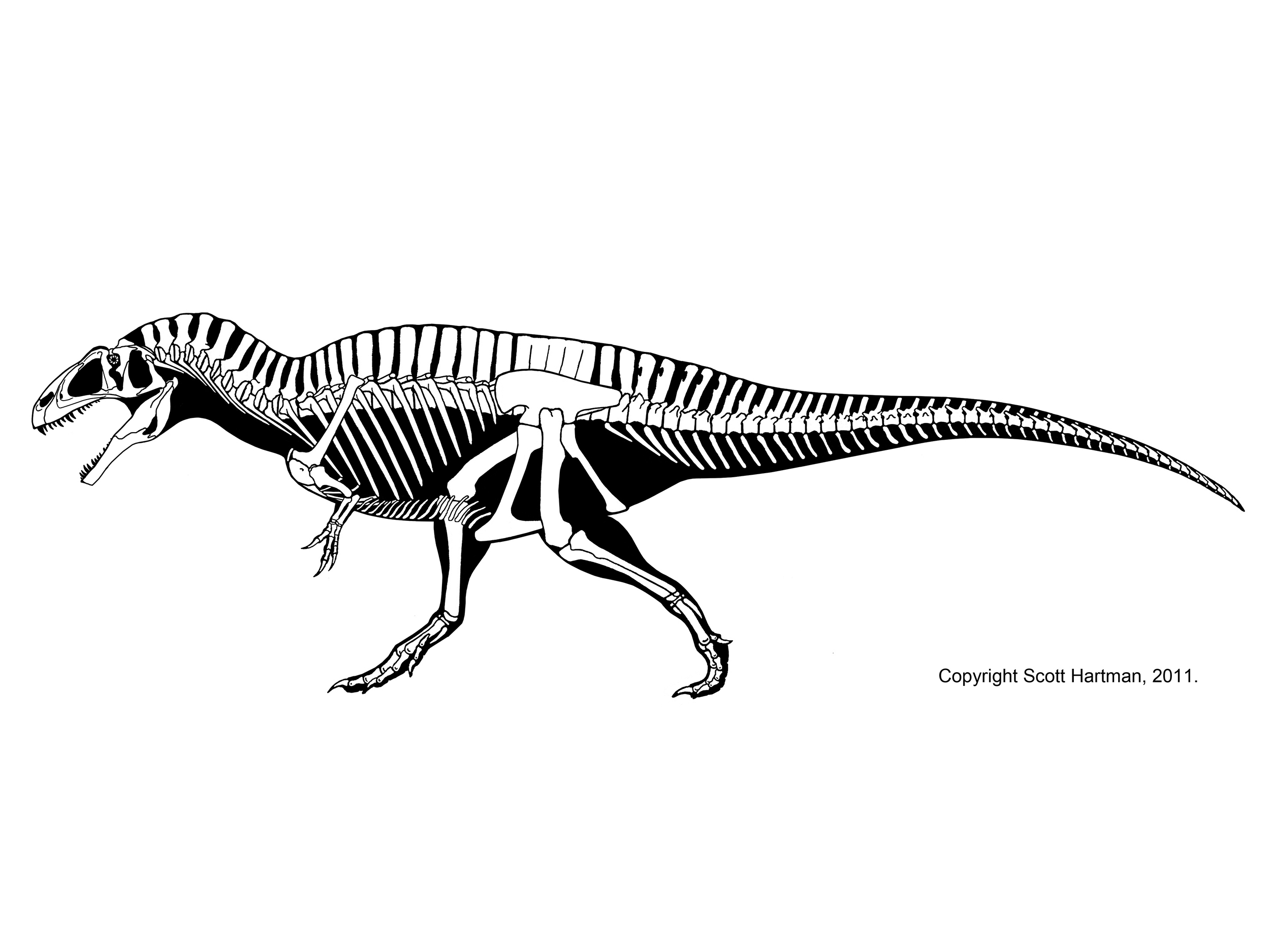
The oldest theropod known (in fact, currently the oldest known North American dinosaur) is Camposaurus of the middle Late Triassic. Much better known, however, is Coelophysis of the late Late Triassic. These were mid-sized carnivores (2-4 m long), and representatives of the true coelophysIDs (Coelophysidae): a clade characterized by long and slender bodies, with slender skulls. The best studied coelophysids are Coelophysis and Early Jurassic southern African Megapnosaurus (formerly called "Syntarsus", but that name is preoccupied by an insect!). (Note, some regard these as the same genus, with "Megapnosaurus" simply late-surviving species of Coelophysis). Camposaurus may be a close relative of Megapnosaurus. Other possible coelophysids were small (~1-2 m long) Late Triassic Procompsognathus of Europe and similar-sized Early Jurassic Segisaurus of the American Southwest and 3 m long Panguraptor of Early Jurassic Asia. Coelophysid footprints are some of the most common trace fossils of the terrestrial Triassic.
Coelophysids seem to be united with a set of larger Late Triassic and Early Jurassic theropods, collectively the Coelophysoidea. These larger coelophysoids include (4-6 m long) primitive theropods of the Late Triassic (Gojirasaurus of the the American Southwest and Liliensternus of Europe). Zupaysaurus of Late Triassic Argentina might be a coelophysoid, or may be intermediate between them and the Dilophosauridae. Another, slightly later Late Triassic Swiss transitional theropod is Notatesseraeraptor.
Long considered the largest coelophysoid is Early Jurassic double-crested Dilophosaurus the American Southwest. However, other analyses place it, Dracovenator of South Africa, Sinosaurus of Asia, and Cryolophosaurus of Antarctica as the clade Dilophosauridae. Shuangbaisaurus of Early Jurassic China is another possible dilophosaurid. Other studies suggest some or all of these are coelophysoids or a paraphyletic grade running from Coelophysoidea up into the base of Tetanurae. In this class we'll refer to this as a grade of "dilophosaurs" rather than a clade Dilophosauridae.
Dilophosaurs (either as a clade or grade) represent the first large dinosaur predators and the first time dinosaurs were the top (apex) predators in their environment, since the big predatory pseudosuchians that "ruled" the Triassic were extinct. As with averostrans, the dilophosaurs have a reduced total number of maxillary teeth. This seems to represent an ecological change from being minor predators feeding on small animals to being predators on other big dinosaurs.
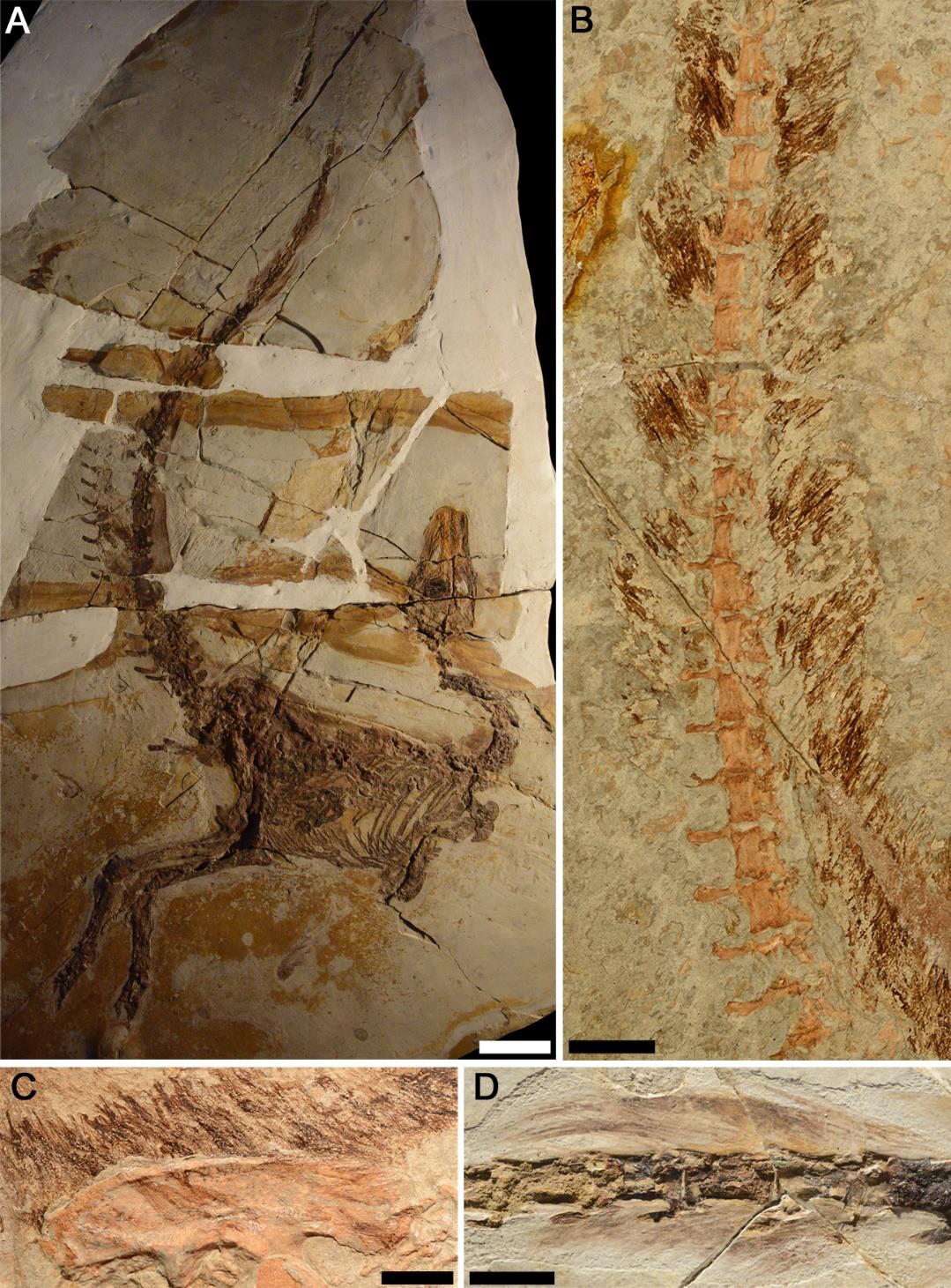
There is evidence that in basal theropods that there was significant sexual dimorphism. The bones of some members of the population were generally more robust, and had more pronounced muscle attachments, while others of the same length were more gracile. Based on observations of modern predatory birds, the robust forms are typically interpreted as female, but this is not certain. Additionally, in Coelophysis and Megapnosaurus there are sites where dozens or hundreds of individuals were found dead together, strongly implying that they were at least on occasion gregarious. This is further supported by the presence of display structures in the dilophosaurids and some basal tetanurines: such display structures are associated with within-species display behaviors, suggesting that at least on occasion they got together. Some coelophysoids, dilophosaurs, basal tetanurines, and even basal coelurosaurs had crests on their skull: these probably served as display structures.
Although very common in the Late Triassic and Early Jurassic, no basal theropods are known after the end of the Early Jurassic. They seem to have been displaced by the averostrans. Curiously, the range of these primitive theropods is very similar to that of "core prosauropods".
Tachiraptor is a theropod from just after the Triassic/Jurassic extinction which appears to the be the sister taxon to Averostra.
AVEROSTRA
The clade comprised of Ceratosauria and Tetanurae has sometimes been called "Neotheropoda"; however, that name has come to mean the more inclusive group that also contains Coelophysidae and "dilophosaurs". The ceratosaur-tetanurine clade, then, is now Averostra ("bird snouts").
Averostrans can be recognized by the following transformations:
The basal members of Ceratosauria and Tetanurae typically have mediolaterally narrow, dorsoventrally deep skulls: sometimes nicknamed "hatchet heads". This skull patterns is good for striking hard against a victim and slicing it up. However, it is not particularly strong if shaken back-and-forth, and so these dinosaurs probably did not hold onto their prey for very long with their jaws. This made primitive averostrans "bite-and-slice" feeders: they could carve chunks out of victims, or wound them, but could not hold onto them with their jaws. (We will see later examples of averostrans that evolved alternative forms of feeding.)
Many basal averostrans run in the 6-8 m range, like the "dilophosaurs". However, these more derived taxa are typically more robustly built, and likely tackled bigger prey. Indeed, their rise coincides with the rise of more advanced and larger herbivorous dinosaurs (thyreophorans, iguanodontians, eusauropods), and the more powerful build of averostrans may be a co-evolutionary "arms race" with the new-style herbivores.
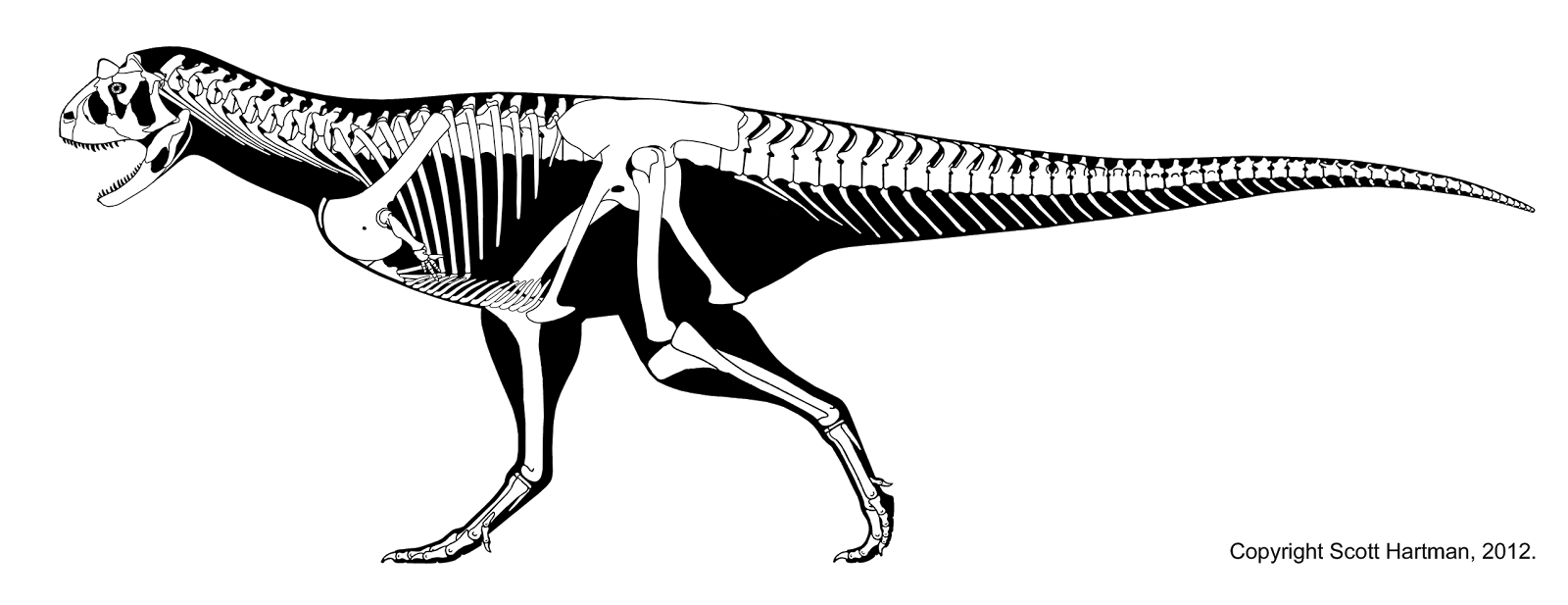
CERATOSAURIA
The ceratosaurs begin as a minor part of the theropod community, but in the Late Cretaceous dominate most of the world (particularly the southern continents and Europe). Ceratosaurs share the following specializations:
Late Early Jurassic Berberosaurus of northern Africa is one the oldest and most primitive ceratosaur; however, some analyses place it within the "dilophosaur" grade (or clade). Unfortunately the specimen is too fragmentary to get much sense of the proportions of this dinosaur. Even older is early Early Jurassic Saltriovenator. Also only partially known, it is among the oldest large-bodied theropod. Interestingly, the manual digits of Saltriovenator are not reduced as in more derived ceratosaurs (and indeed was long considered the oldest tetanurine before the detailed analysis of this specimen.)
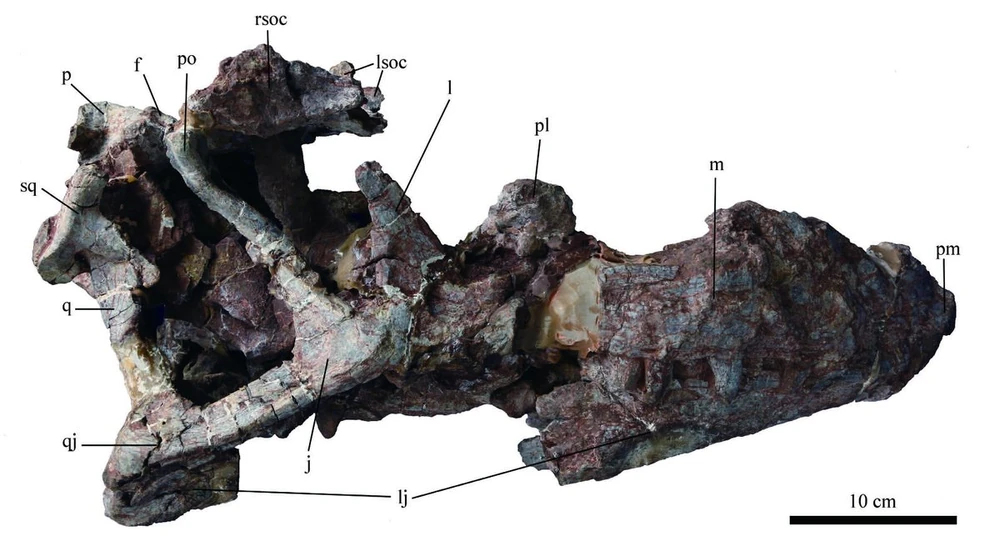
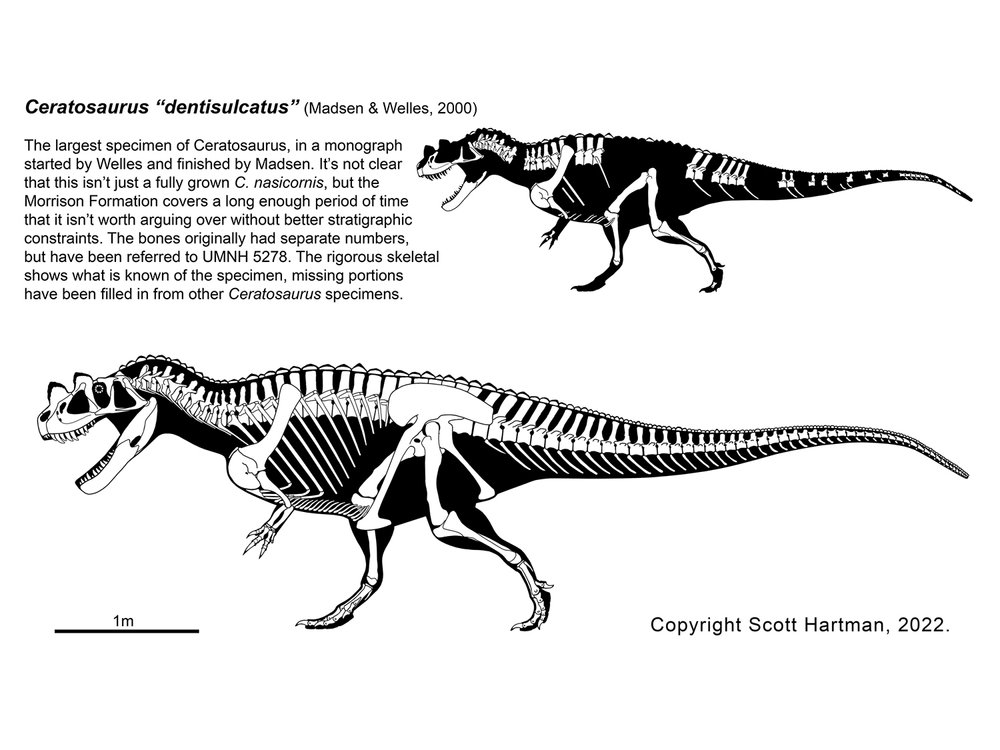
Most of the ceratosaurs belong to the clade Neoceratosauria. These include the larger ceratosaurs, and are typically relatively short-necked forms. Among these are the Ceratosauridae proper. These are best known from 6-8 m long Ceratosaurus of the Late Jurassic western North America and Europe.
The most successful group of ceratosaurs is the Abelisauroidea, a clade that includes the top predators of South America, India, Madagascar, and Europe (and for all we know continental Africa and Australasia/Antarctica) during the Late Cretaceous, the Abelisauridae, as well as a diverse paraphyletic series of "noasaurs".
Traditional phylogenies found the non-abelisaurid abelisauroids as a single clade "Noasauridae". However, the latest analyses find these as a paraphyletic series that encompasses a wide variety of body plans. Some were elongate cursorial (running) forms, such as Kiyacursor of Early Cretaceous Siberia, 6 m long Elaphrosaurus of Late Jurassic eastern Africa, and similar unnamed "elaphrosaur" forms from the same age in Africa and western North America.
A particularly bizarre group is a clade whose precise proper name is uncertain: possibly "Bahariasauridae", or perhaps "Berthasauridae". The most completely known member is early Late Jurassic toothless Chinese Limusaurus, for whom the ontogenetic series is represented. (Curiously, they had teeth as youngsters but lose them in favor of a toothless beak as adults, suggesting an ontogenetic change of diet.) Also toothless is short-snouted Berthasaura of Early Cretaceous Brazil. Tiny (< 1 m long) Ligabueino seems to belong here as well.
Most spectacular amonag these bahariasaurids are giants from the early Late Cretaceous of Argentina and Africa. The most completely known is African Deltadromeus and Bahariasaurus (which may be the same taxon, and has priority in terms of name). Bahariasaurus shows that these dinosaurs reached extremely large size, as it is almost as large as Giganotosaurus or Tyrannosaurus! Giant bahariasaurids show a mixture of different traits, making them extremely difficult to pin down phylogenetically. Once they were thought to be related to the neovenatorid allosauroids; or alternatively basal coelurosaurs (possibly members of Megaraptora.) The latest analyses put at least some of these (especially Deltadromeus/Bahariasaurus) as abelisauroids. At present we don't have any skull bones from them, so we know nothing of significance about their feeding ecology: were they armed with sharp teeth? Did they have toothless beaks? They are one of the big mysteries of theropod paleontology at the moment.
Closer to abelisaurds are some small Cretaceeous forms. Vespersaurus of Late Cretaceous Brazil was a desert-dwelling noasaurine: it is described as being functionally monodactyl (using only its pedal digit III to support its weight), with digits II and IV raised as cat-like retractable claws. Afromimus from Niger was originally interpreted as an ornithomimosaur, but re-analysis shows that it is likely the sister taxon to Vespersaurus. The (now much reduced) proper Noasauridae includes 1.5 m long Velocisaurus and 2-3 m long Noasaurus of Late Cretaceous South America and Masiakasaurus of Late Cretaceous Madagascar.
Middle Jurassic Eoabelisaurus of Argentina appears to be a true abelisaurid. However, in some phylogenies appears to be in the same general region of the phylogeny as the ceratosaurids. Its arms are not as strongly reduced as the later true abelisaurids.
More derived abelisaurids such as (such as early Late Cretaceous Rugops) and Kryptops of early Late Cretaceous northern Africa were minor predators compared to their neighbors the spinosaurids and carcharodontosaurids (about whom see below). With the extinction of those two groups, however, the abelisaurids came into their own.
Abelisaurids are further specialized from other abelisauroids by:
The particulars of their forelimbs show that they were useless in grappling; their tough skulls and stout teeth suggest that they may have used their skulls to hold onto prey with their jaws in order to kill it.
Notable Late Cretaceous abelisaurids include Rajasaurus of India; Majungasaurus (formerly "Majungatholus") of Madagascar; and Abelisaurus, Aucasaurus, Skorpiovenator, Spectrovenator, Exkrixinatosaurus, and Carnotaurus of South America.
Abelisaurids make it all the way until the end of the Cretaceous. Interestingly, their stratigraphic range and geographic distribution closely matches that of lithostrotian titanosaurs.
TETANURAE
The remaining theropods form the Tetanurae ("stiff tails"). Tetanurines (some prefer the form "tetanurans") are specialized from earlier theropods in possessing:
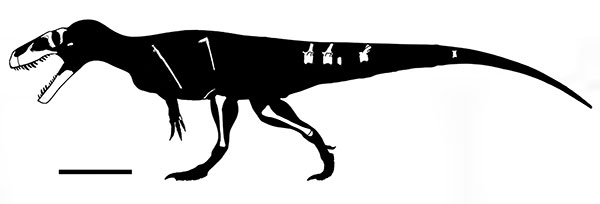
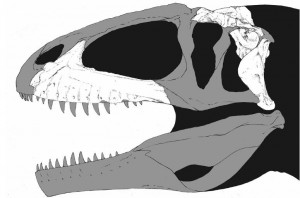
Basal tetanurines tended to be large (5-8 m long) hatchet-headed carnivores. Some analyses place Early Jurassic Sinosaurus of China and Cryolophosaurus at the base of Tetanurae. A basal tetanurine is Middle Jurassic Monolophosaurus of the Middle Jurassic of China (once considered one of the oldest and most basal carnosaurs, or alternatively as a primitive megalosauroid). Note that all of these have some form of crest on the head: this was apparently the "fashion" for Early and Middle Jurassic big theropods.
The three major clades within Tetanurae (Megalosauroidea (also known as Spinosauroidea), Carnosauria, and Coelurosauria) are united into the clade Orionides ("hunters"). Both megalosauroids and coelurosaurs are confirmed to have protofeathers (not yet demonstrated in carnosaurs). Protofeathers are simple, apparently hollow, down-like tufts on the body. They represent the evolutionary precursors to true feathers. In this primitive state, may have helped to insulate; for display; for brooding; or some other function. Note that if these do prove to be homologous with the fuzz of the heterodontosaurid Tianyulong than protofeathers would be shared derived features of Dinosauria (at least!) in the Saurischia model, or Ornithoscelida if that model is correct, and not just Orionides. At present, though, the lack of positive evidence of any such structure in non-tetanurine theropods or in sauropodomorphs means that this is not the simplest explanation.
Until 2012 the only definite protofeathers known in theropods were in primitive coelurosaurs. However, the discovery of Sciurumimus (a possible megalosauroid known only from a juvenile specimen) shows that at least small megalosauroids were fuzzy. (It is true that some think that Sciurumimus may turn out to be a primitive coelurosaur rather than a megalosaur.)
It is not yet certain if the protofeathers found in Sciurumimus are all simple strands or tufts (plumulose, or downy, feathers) or if some might not have a central shaft (pennaceous feathers).
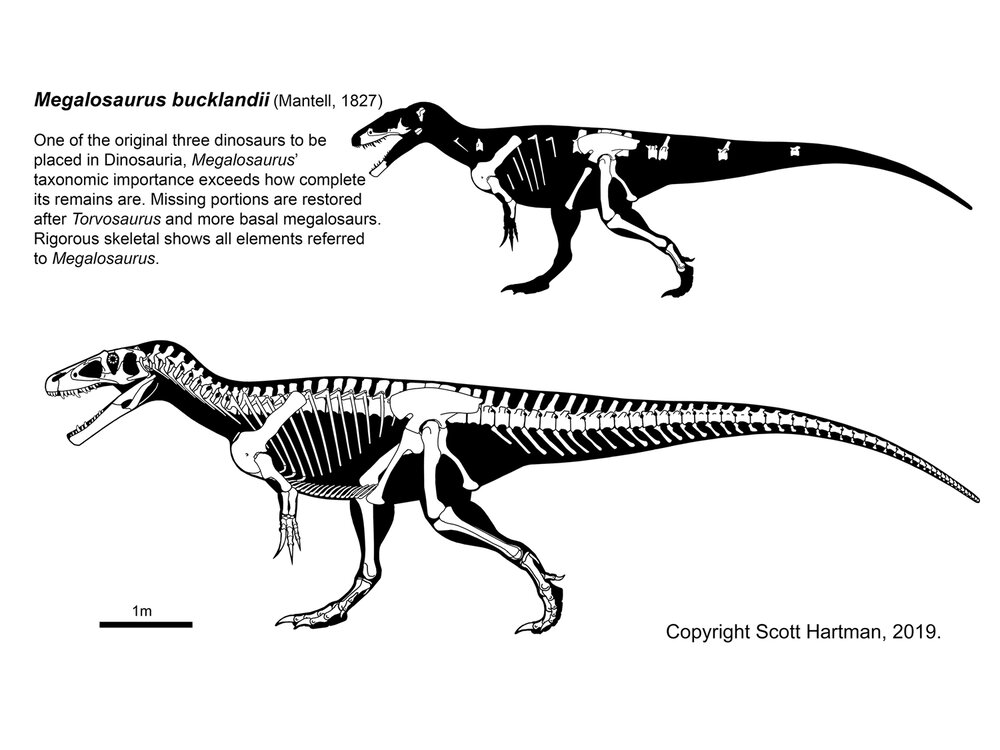
One of the first major clades of tetanurines are the Megalosauroidea (often also known as Spinosauroidea). Megalosauroids share elongate skulls. Primitive megalosauroids include the Piatnitzkysauridae (such as Piatnitzkysaurus of Middle Jurassic Argentina and Marshosaurus of the Late Jurassic of the western US). More specialized were the Megalosauridae. This group contains Megalosaurus, Duriavenator, Poekilopleuron, and Dubreuillosaurus of Middle Jurassic Europe; Middle Jurassic Afrovenator of northern Africa; Eustreptospondylus of Late Jurassic Europe; and massive Wiehenvenator of Middle Jurassic Germany and its even larger close relative Torvosaurus of Late Jurassic North America and Europe. One trait that unites the megalosauroids is that the maxillary fenestra becomes a fossa (that is, the sinus no longer punches all the way through the maxilla bone, but forms a 'divot' on the lateral surface).
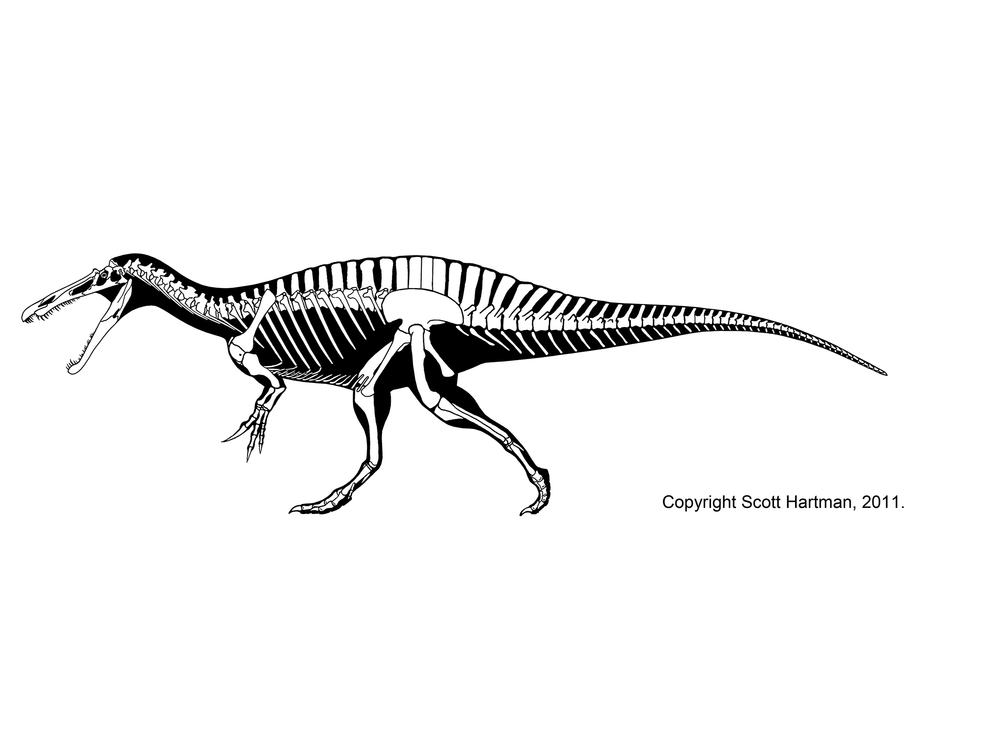
Giant Torvosaurus shares its enormous size and powerfully-developed forelimbs with the Spinosauridae. The spinosaurids are a group of Late Jurassic to mid-Late Cretaceous giant (8-14 m long) theropods characterized by:
Additionally, spinosaurids share with at least some of the megalosaurids an enormously enlarged thumb claw (even by saurischian standards).
The adaptations of the crocodile-like spinosaurid jaws and teeth (as well as their gut contents) suggest that they added large fish as well as dinosaurs to their diet, and chemical analyses of their bones show that they ate substantial amounts of food from the water. All spinosaurids have been discovered in environments in which large fish are common.
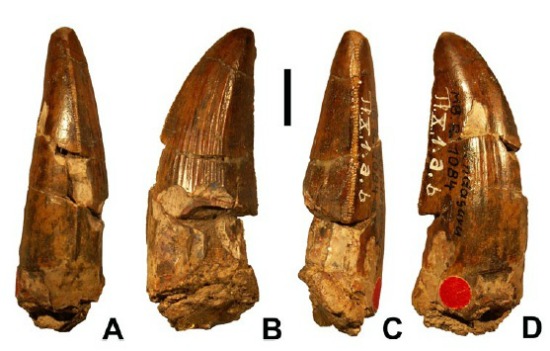
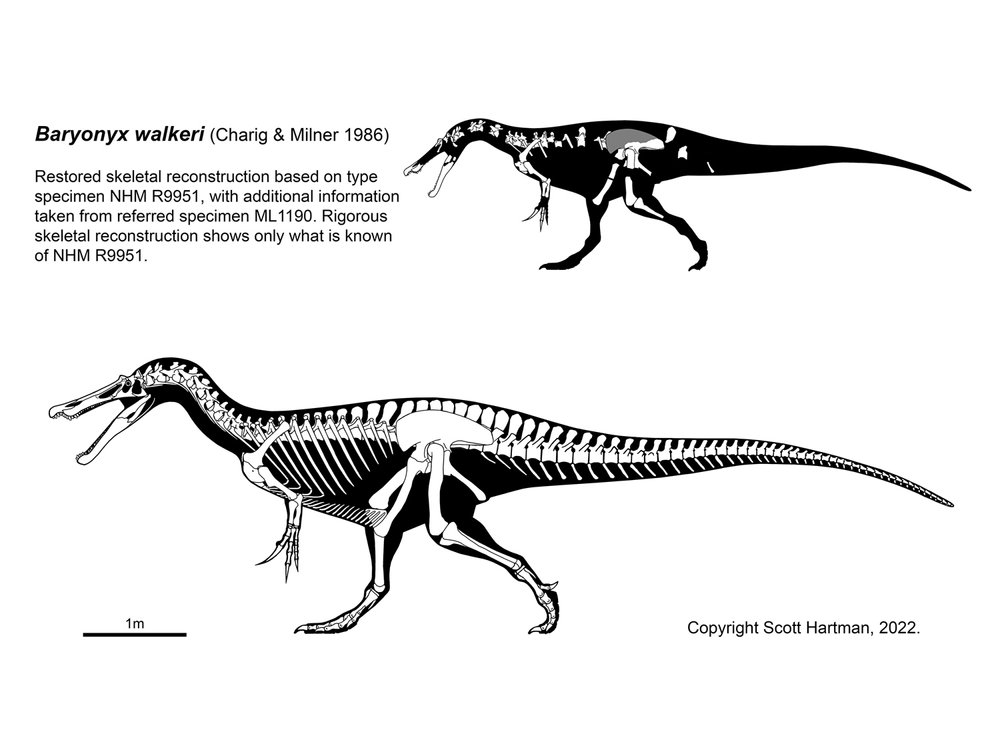
Once thought to be a Jurassic spinosaurid is Ostafrikasaurus of the Late Jurassic of (not surprisingly) eastern Africa, known only by its teeth and now considered a probable ceratosaurid rather than a spinosaurid. More complete spinosaurids include Baryonyx of Early Cretaceous Britain (less complete are the closely-related Riparvenator and Ceratosuchops also of Early Cretaceous Britain and Protathlitis and Riojavenatrix of the same age in Spain); Suchomimus of the Early Cretaceous of northern Africa; Irritator of the Early Cretaceous of Brazil; Ichthyovenator of Early Cretaceous Thailand; and giant (14 m long) Spinosaurus of the early Late Cretaceous of northern Africa. Spinosaurus is one contender for the largest known theropod of all time. It was also the youngest named spinosaurid (and indeed megalosauroid), although possible spinosaurid material is known from middle Late Cretaceous of China. However, none are yet known from the later Late Cretaceous, and thus this large clade is long gone before the end of the Cretaceous. Curiously, although this clade has been found on nearly every continent, at present there are no known North American spinosaurids.
New skeletal material shows that Spinosaurus was even more bizarre than previously thought. The hind limbs (at least) were solid (in other theropods, even giants, they are hollow). The hind limbs and pelves are proportionately shorter than expected in a dinosaur of this size. The only known tail shows very tall neural spines. Many have interpreted this suite of features as suggesting that perhaps Spinosaurus may have been aquatic: more like a crocodile than a heron in terms of the way it approached food. If these discoveries are correct, it may have actually spent relatively little time on land. (By the way, the super-large forelimbs in these reconstruction is almost certainly wrong, as the humerus used to scale the rest of the arm is almost certainly that of the diplodocoid sauropod Rebbachisaurus, and not Spinosaurus at all! Thus, claims that Spinosaurus was a quadruped are not yet supported.) However, other evidence shows that it could not have been a fast swimmer, and thus most likely did not chase after prey in the water. Instead, it likely was a shoreline feeding opportunist, fishing and probing for food, and swimming from location to location to feed.
NEOTETANURAE and AVETHEROPODA
The remaining groups of theropods comprise the Avetheropoda ("bird theropods") (sometimes "Neotetanurae", or "new tetanurines"). Avetheropods share the following transformations from the ancestral tetanurine condition:
Note: in pre-1990s literature, these terms were often used as synonyms for "big theropod" and "little theropods", respectively. So larger coelophysoids, ceratosaurids, abelisaurids, spinosauroids, and tyrannosaurid coelurosaurs were considered by many to be "carnosaurs", while small coelophysoids were included with the "coelurosaurs". Since the rise of cladistic studies, however, these names are restricted to two branches of the derived tetanurines.
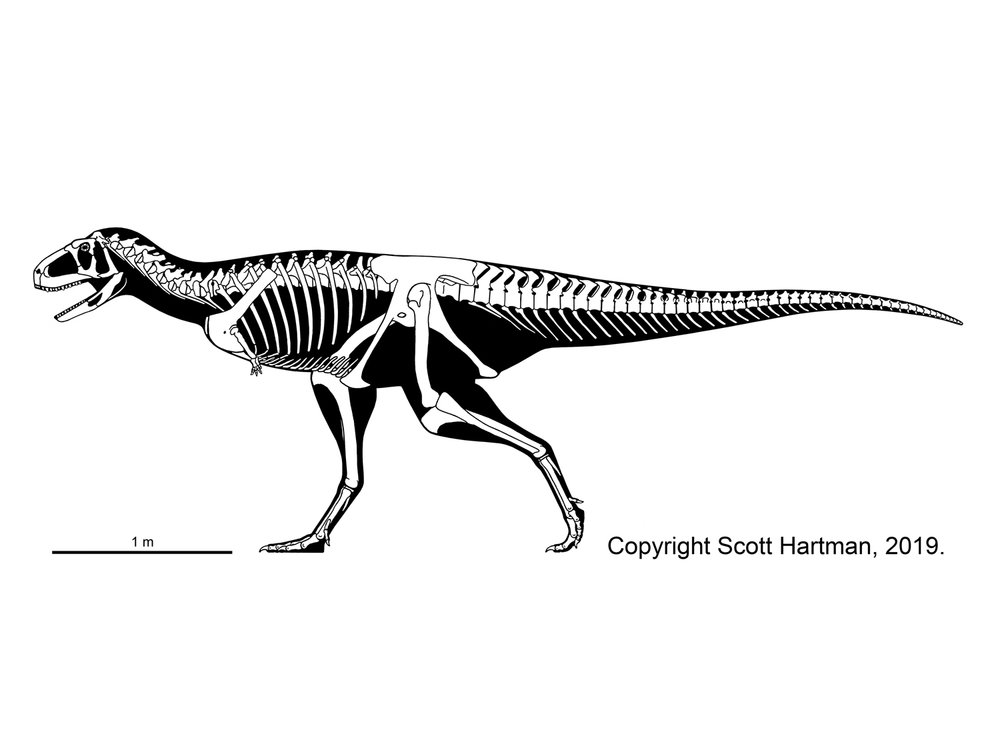
CARNOSAURIA
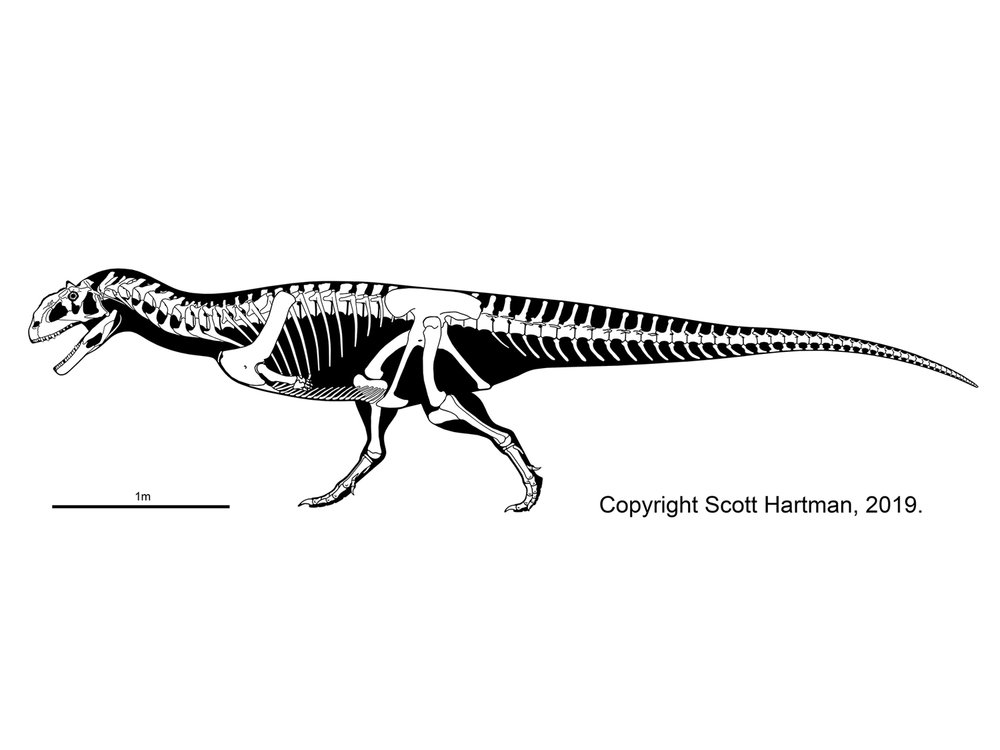
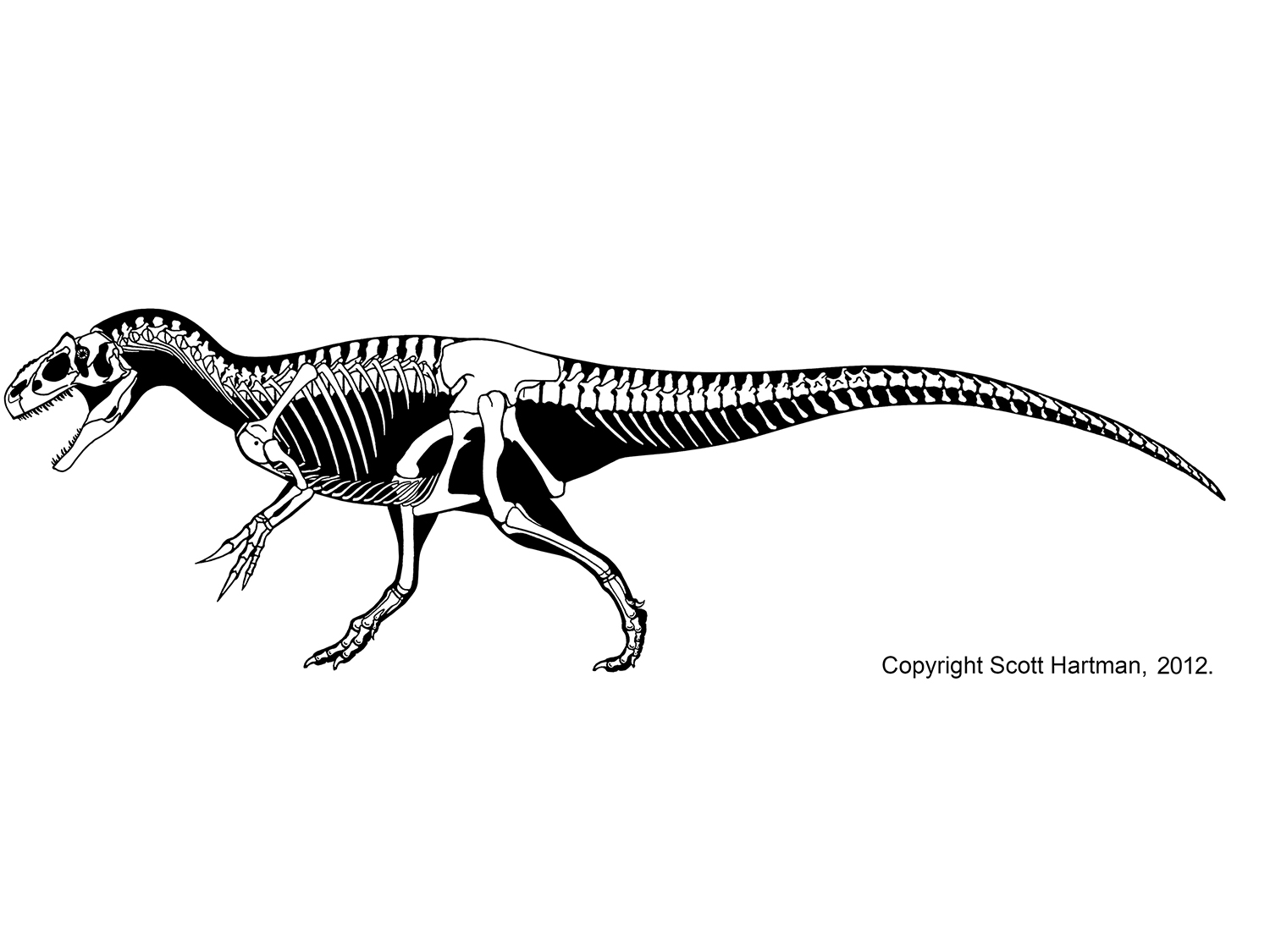
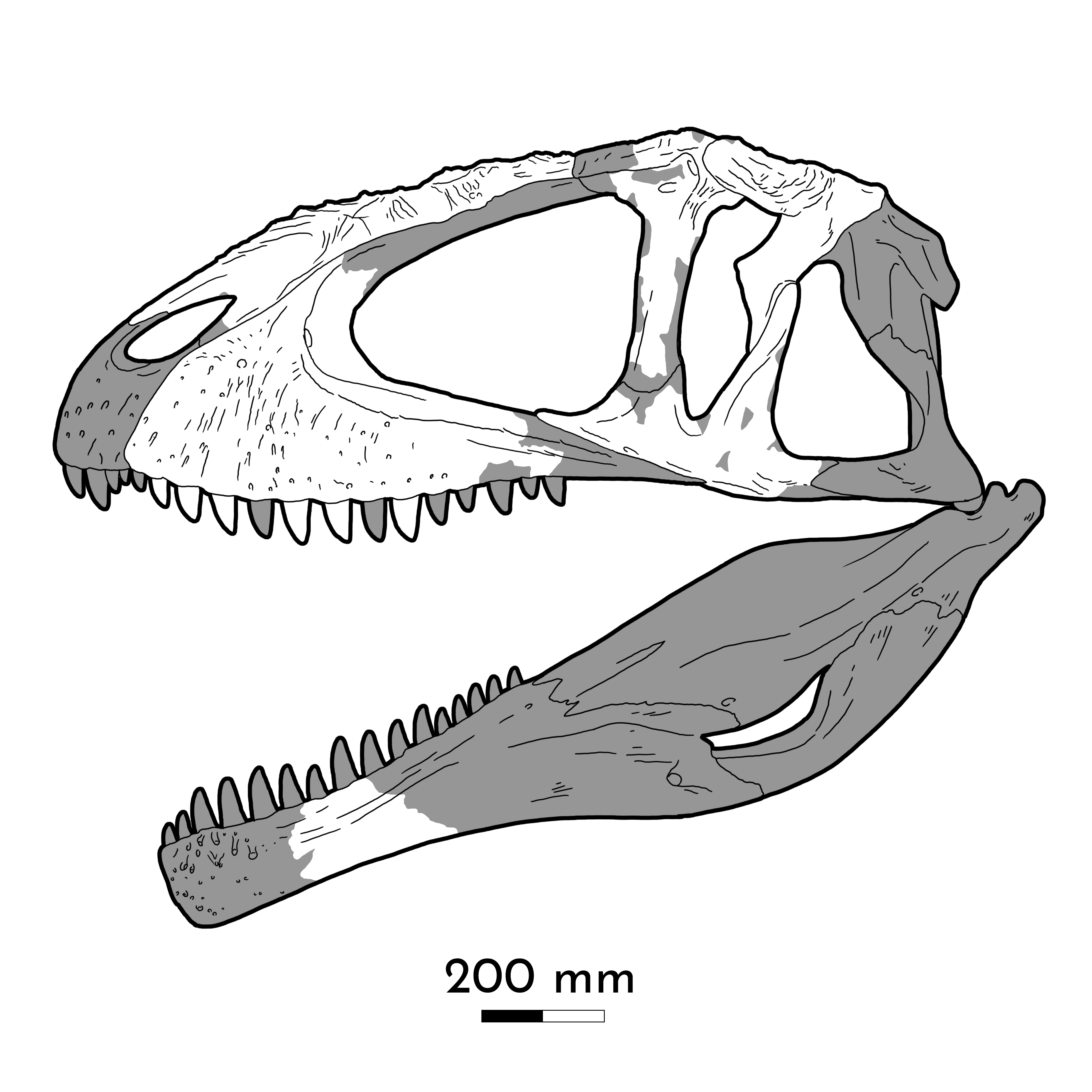
The dominant group of large theropods from the Middle Jurassic through the Early Cretaceous, although a few persist until the middle part of the Late Cretaceous. Carnosaurs (sometimes called Allosauroidea) are best known in the form of Late Jurassic North American and European Allosaurus. Carnosaurs are characterized by:
(However, some of these traits are also shared with Monolophosaurus, and may actually be basal tetanurine rather than carnosaur traits!)
One primitive clade of carnosaurs the Metriacanthosauridae (formerly Sinraptoridae) of the Middle Jurassic to Early Cretaceous of Asia (such as Jurassic Sinraptor and Yangchuanosaurus of China and Cretaceous Siamotyrannus of Thailand) and Europe (such as Metriacanthosaurus of early Late Jurassic England, and possibly Lourinhanosaurus of the Late Jurassic of Portugal (which some studies suggest is a basal coelurosaur). Larger are the Allosauridae of Late Jurassic North America and Europe, known primarily from various species of Allosaurus. The largest (13 m long) specimens once were given the name of "Saurophaganax", but these are now considered as a species Allosaurus anax.
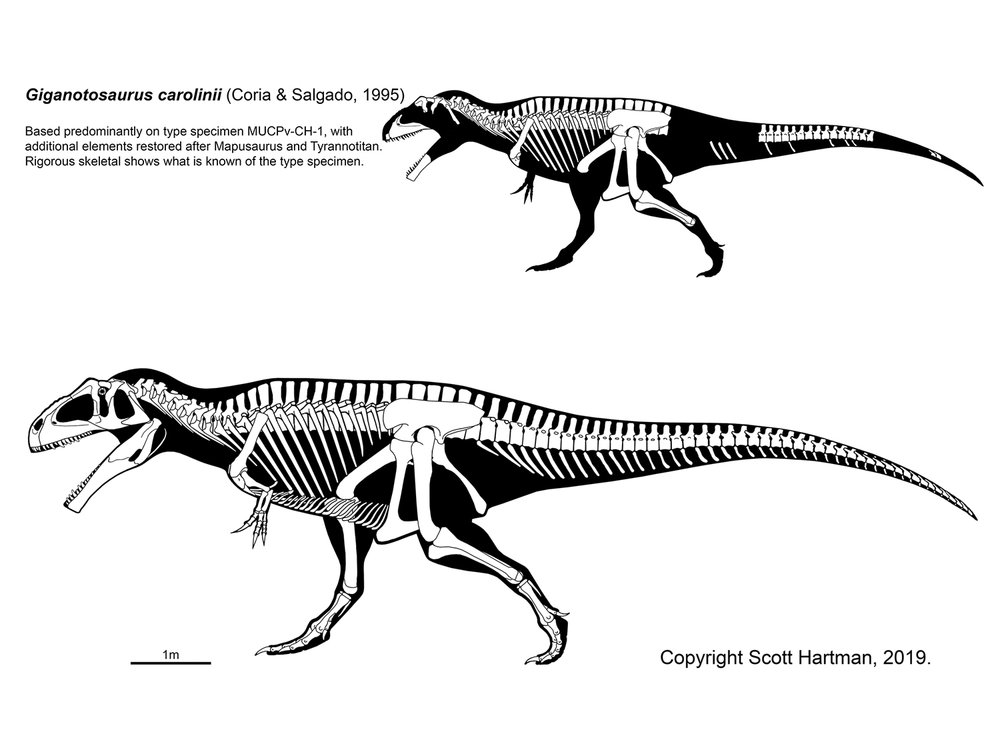
The Cretaceous carnosaurs mainly consist of the clade Carcharodontosauria. Most studies show that the carcharodontosaurs contain two major groups: the more massive, powerfully built Carcharodontosauridae and the Neovenatoridae. (A Late Jurassic genus Veterupristisaurus from the Late Jurassic of eastern Africa may be an early primitive carcharodontosaur, or even cacharodontosaurid.) Primitive carcharodontosaurids include giant (12-13 m long) Acrocanthosaurus of the later Early Cretaceous of North America, Concavenator of Early Cretaceous Spain; Eocarcharia of the late Early Cretaceous of northern Africa and Shaochilong of China. The most specialized carcharodontosaurids are those of the late Early Cretaceous and early Late Cretaceous of South America and Africa. These include Tyrannotitan and Giganotosaurus, of the late Early Cretaceous of South America; Meraxes and Mapusaurus of the early Late Cretaceous of the same region; and Carcharodontosaurus of the same age in Africa (making these the youngest of the carcharodontosaurids proper.) Carcharodontosaurids may have been sauropod-eating specialists, and indeed many co-occur with particularly large titanosaur or brachiosaurid sauropods. Carcharodontosaurids are among the largest theropods known: in particular, Mapusaurus, Tyrannotitan, and Giganotosaurus just about equal the largest individuals of Tyrannosaurus rex in size, and rivaled the largest Spinosaurus specimens in mass (although the latter was probably longer, given the relatively long snout and neck of spinosaurids). It seems that these dinosaurs and the spinosaurids disappear around the same time, for reasons as yet uncertain. (Some recently described teeth and jaws from the end of Cretaceous in Brazil were initially thought to be from carcharodontosaurids, but new study shows these are more likely abelisaurid fossils.)
The Neovenatoridae are a clade (or grade?) closely related to the carcharodontosaurids. It includes forms such as Neovenator of the Early Cretaceous of Europe and giant Chilantaisaurus of early Late Cretaceous China. At least some models include the Megaraptora (see next lecture) in this clade. The last giant non-tyrannosaurid theropod known in North America, early Late Cretaceous Siats of Utah might be a neovenatorid, but might be a megaraptorid instead.
Re-Expanding Carnosauria: As noted above, "Carnosauria" once included all the large theropods. A potential version of this--namely, a clade uniting Megalosauroidea and Allosauroidea to the exclusion of Ceratosauria and Coelurosauria--has been proposed as early as the late 1990s. The discovery of Middle Jurassic Argentine Asfaltovenator has reenergized this hypothesis. This dinosaur shares traits with both primitive allosauroids and primitive megalosauroids, and when included in some phylogenetic hypotheses has united this expanded Carnosauria. If this is supported in future work, "Orionides" and "Avetheropoda" wind up being the same clades.
EVOLUTIONARY PATTERNS IN BASAL THEROPODA
Feeding adaptation transformations:
Like the Jabberwock, the theropod predatory armament consisted of "jaws that bite" and "claws that catch". Each of these were modified in different ways among the Theropoda:
Locomotory adaptations:
Gigantism and Miniaturization:
Co-Evolution:
Increased mass and "fire power" of basal averostrans occurs in time and space with the rise of advanced larger herbivorous dinosaurs: ankylosaurs, stegosaurs, iguanodontians, eusauropods. It may be that there was an co-evolutionary arms race between predators and prey: new types of offensive weaponry in the former, new types of defenses (increased body armor, social behaviors, and size) in the latter.
Niche partitioning:
In many environments several different theropods shared the same habitat. In some cases they may have partitioned the resources by body size (although the juveniles would still overlap). But in the case of the spinosaurids there seems to have been evolution of the ability to access meat that other theropods couldn't: fish. Similarly, spinosaurids could travel more easily from lake to lake and also capture food more easily on land than the giant crocodyliforms that were their main competitor for fish.

{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}