


Key Points:
•Coelurosaurs are the most diverse clade of theropods (or dinosaurs, for that matter). Coelurosaurs began as small-bodied agile hunters, but quickly diversified into many distinct subgroups. All groups are known to have some form of fuzzy or feathery body covering.
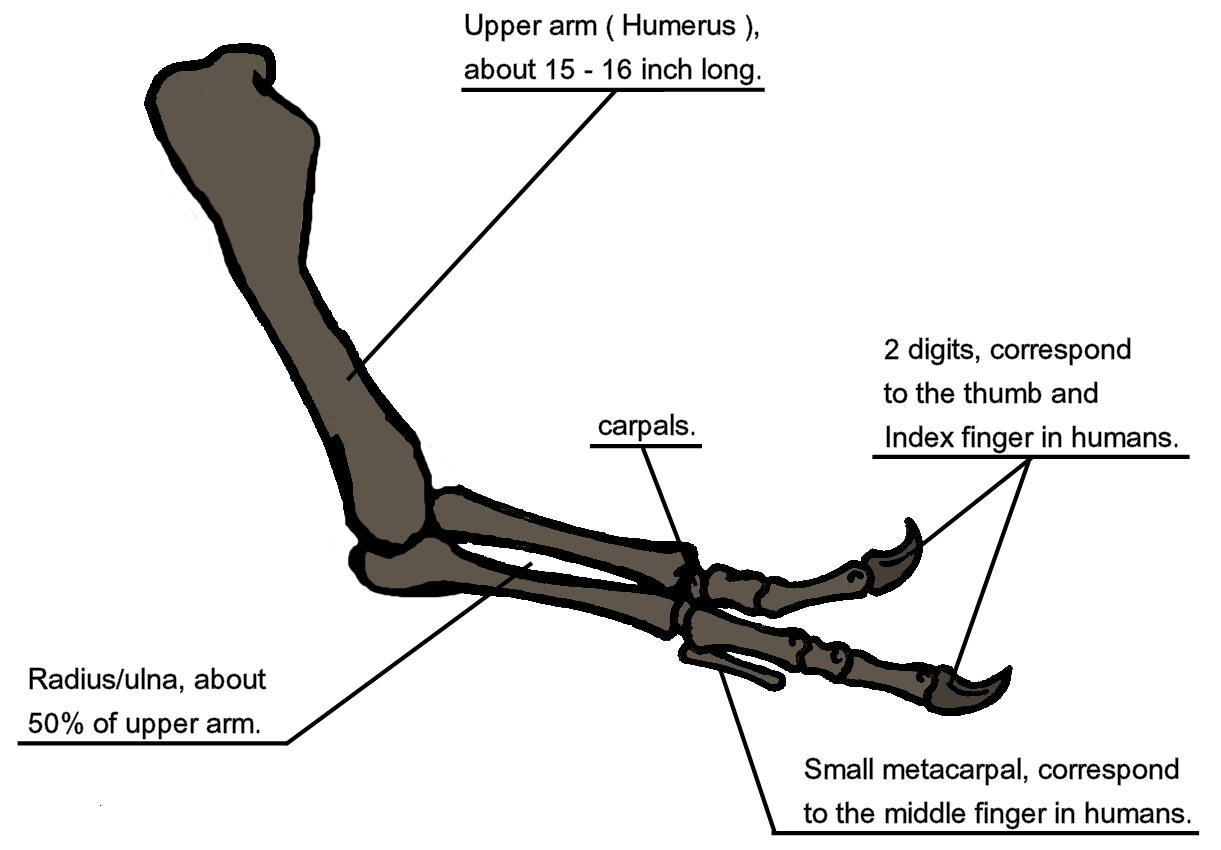
•Tyrannosauroids were a clade with enhanced bite and specialized teeth. Initially mid-sized predators, they culminated in the gigantic, short-armed, two-fingered, long legged, bone-crushing Tyrannosauridae.
•More derived forms typically had smaller skulls, long necks, and phyllodont dentition, showing a shift away from flesh-eating.
•Ornithomimosauria had long arms with clamping hands. This clade includes toothless forms, such as the large-to-gigantic Deinocheiridae and the slender-footed Ornithomimidae.
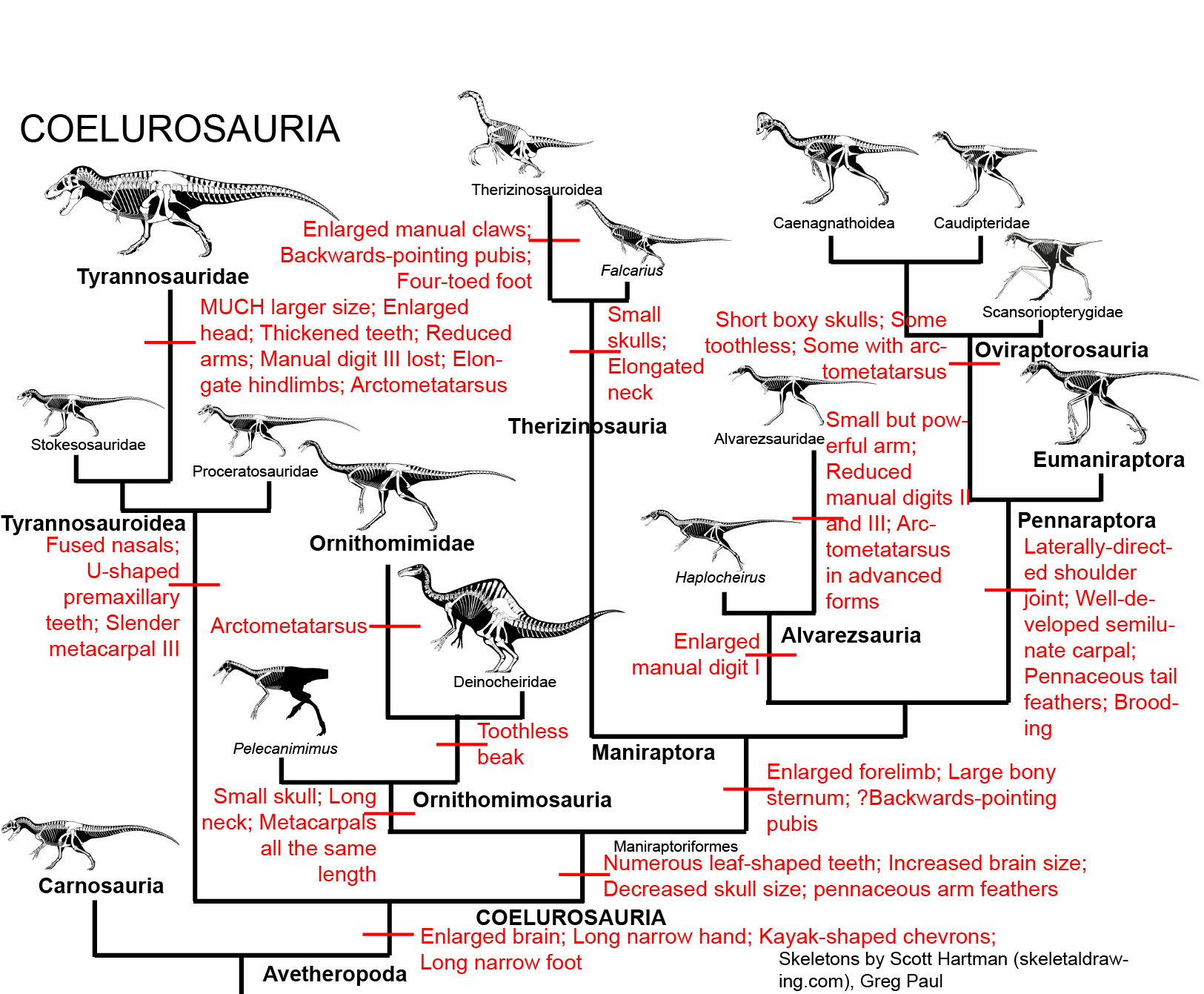
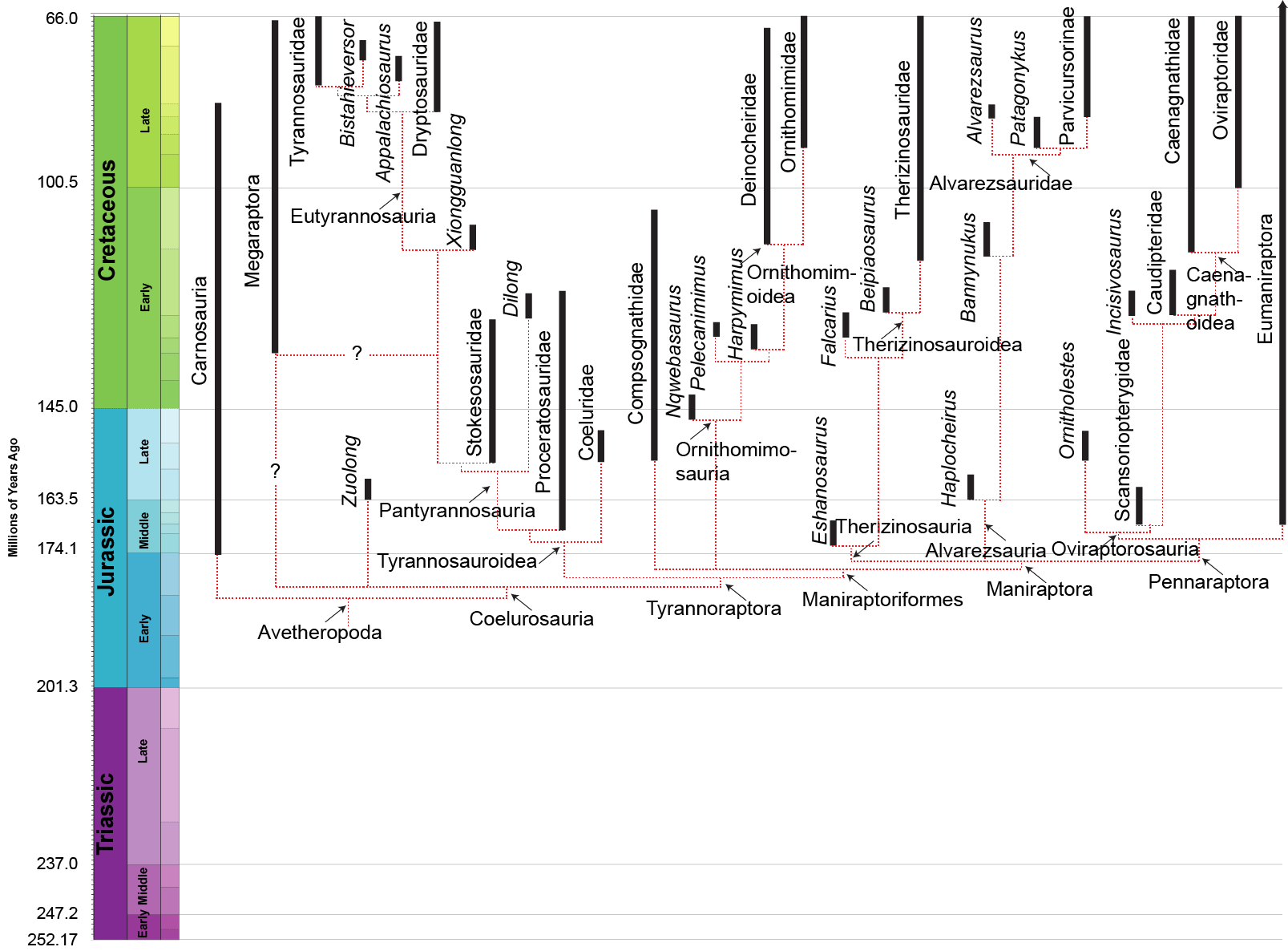
MAJOR GROUPS OF COELUROSAURS
Coelurosaurs are the sister group to Carnosauria within the avetheropods. The coelurosaurs differ from other theropods by possessing:
Additionally, where known, all coelurosaurs preserved in the appropriate-style sediment show at least some protofeathers or true feathers. However, as discussed previously, the discovery of the megalosauroid Sciurumimus pulls the origin of protofeathers much deeper into theropod (or dinosaur, or ornithodiran) history. At least some of the protofeathers of primitive coelurosaurs seem to be pennaceous (having a central shaft), rather than simply plumulose tufts.
Among the most primitive and oldest known coelurosaurs are the basal tyrannosauroids Proceratosaurus of the Middle Jurassic of England and Kileskus of Russia. Only the skull of the former, and skull, hand, and foot bones of the latter, are known at present. However, one of the most primitive known coelurosaurs is actual a relatively late one: Bicentenaria of the mid-Cretaceous of Argentina. It shares with basal tyrannosauroids and basal maniraptoriforms the same general body plan: relatively small (2-4 m) slender animals with skulls full of small ziphodont teeth. Their narrow grasping hands suggest they adapted to catching small prey; their light build, slender limbs, and narrow dynamic stabilizing tail suggests relatively agile animals (useful both in chasing prey and in avoiding predators). Additional basal coelurosaurs include the late Middle Jurassic Aorun (possibly an early alvarezsaur) and the early Late Jurassic Zuolong of China.
Some studies find the Late Jurassic western North American Coeluridae (Coelurus and Tanycolagreus) as basal members of Tyrannosauroidea and their neighbor Ornitholestes is a basal member of Maniraptoriformes (and possibly even of Oviraptorosauria). However, these do not show the derived features of their relative clades which will be discussed below (their position based on other skeletal traits not discussed in this course), so we won't address them there. (Furthermore, other research finds them in various other positions)
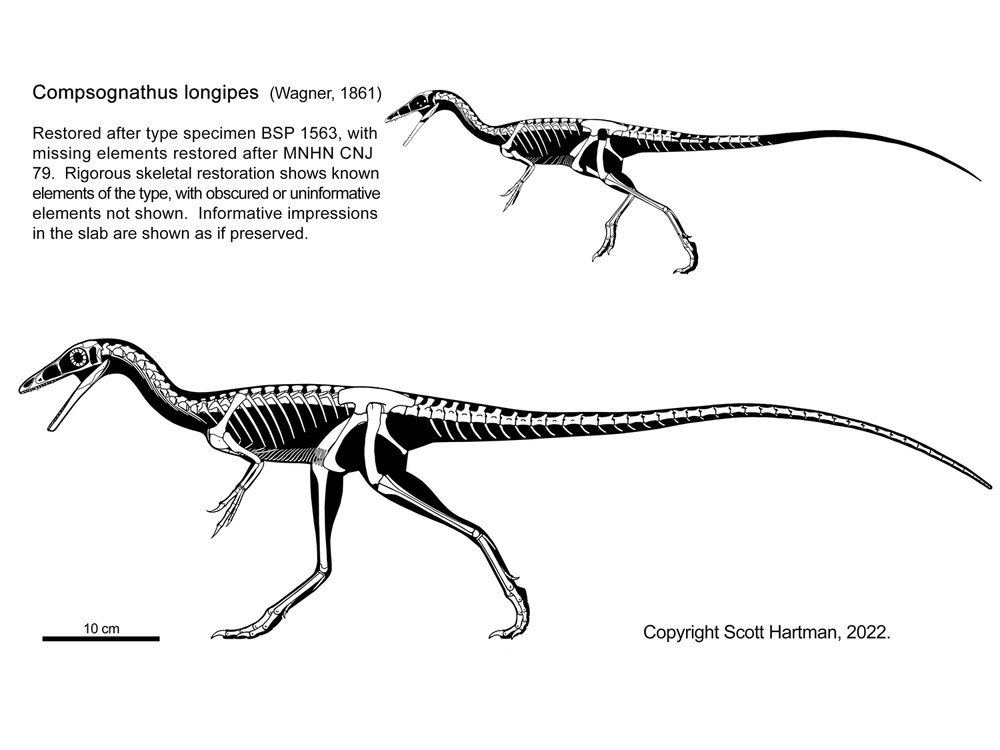
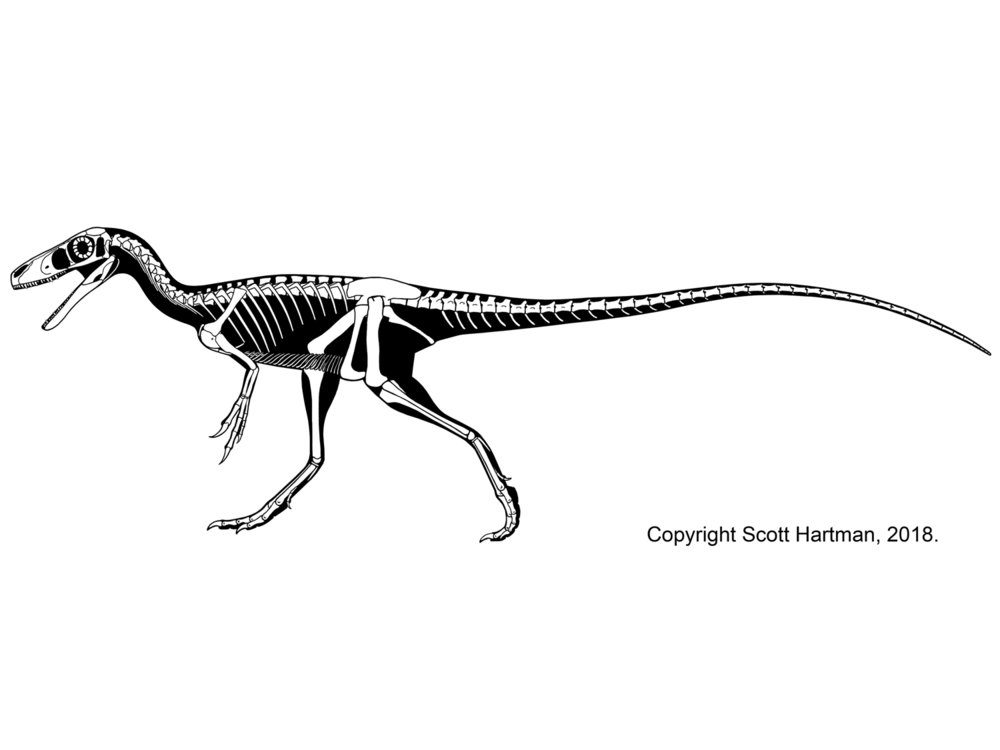
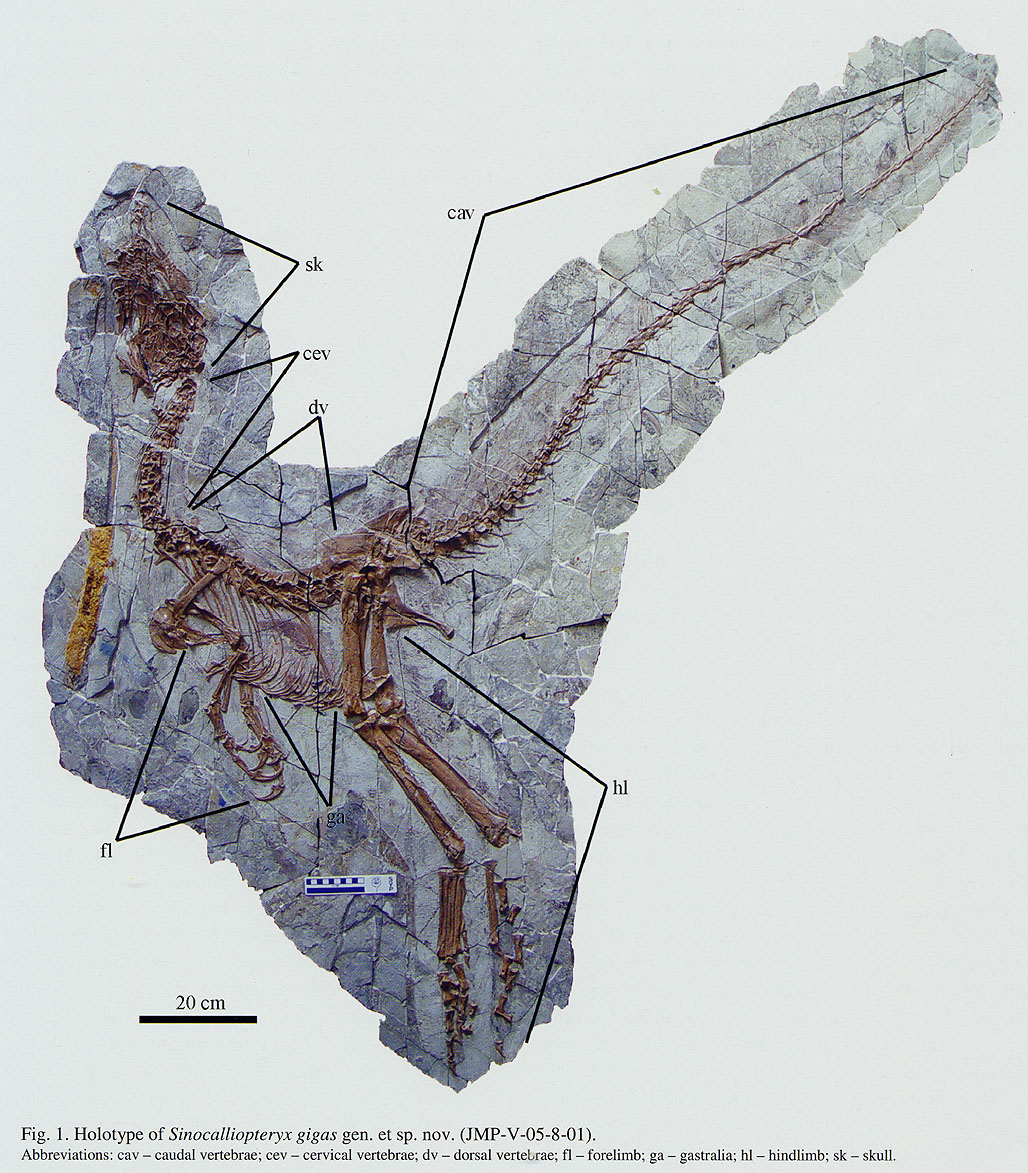
Traditionally, an important group of small primitive theropods was the Compsognathidae. This group ranges from the 1 m long Compsognathus of the Late Jurassic of Europe and Sinosauropteryx of the Early Cretaceous of China to 1.75 m long Huaxiagnathus of the Early Cretaceous of China to the "giant" Sinocalliopteryx of the Early Cretaceous of China at 2.5 m long. Compsognathids are also known from Early Cretaceous Europe and South America, and represented a minor radiation of small-bodied dinosaurs. Gut contents show that they ate lizards and small mammals. Being primitive and of generalized form, these dinosaurs show up in the phylogeny sometimes as basal coelurosaurs outside Tyrannoraptora (the tyrannosauroid-maniraptoriform clade); sometimes as basal maniraptoriforms (as shown here); and sometimes as basal maniraptorans. In some recent studies, though, there was no "Compsognathidae": instead some of these were megalosauroids, some carnosaurs, some basal coelurosaurs (the traditional position), and some basal tyrannosauroids. It does appear that all "compsognathid" specimens are really juveniles, and so it is quite likely that they are not a clade at all.
TYRANNOSAUROIDEA
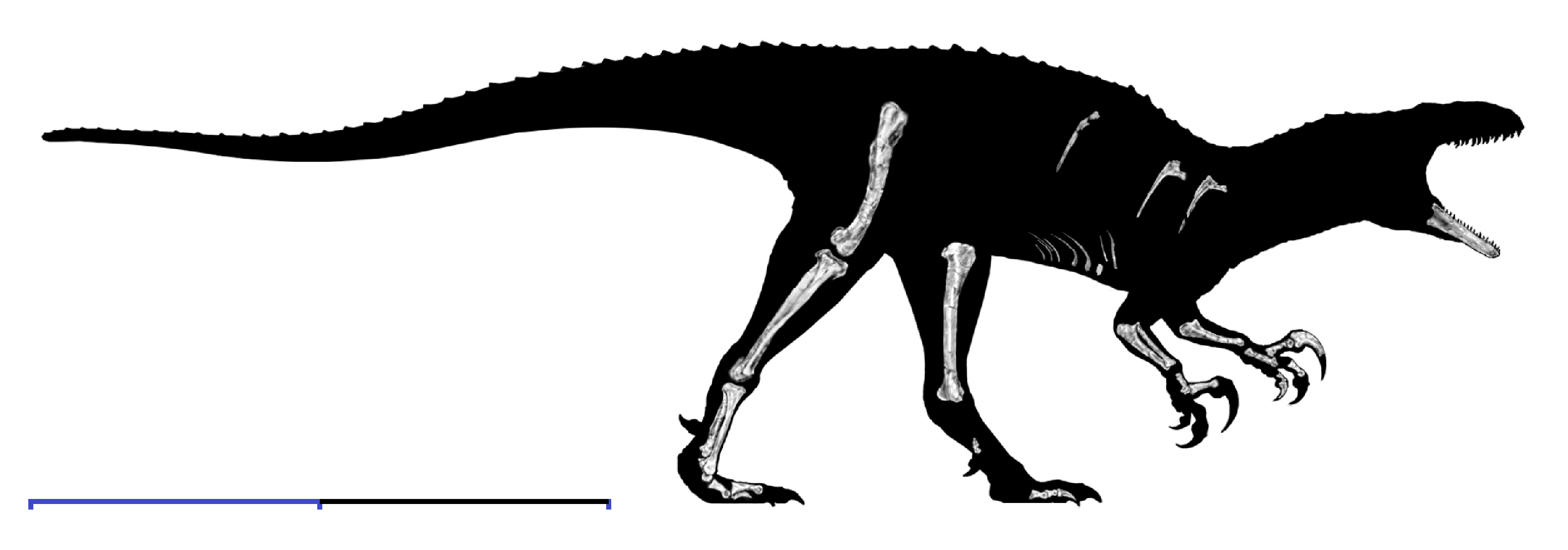
The most long-lived and ecologically significant group of primitive coelurosaurs was Tyrannosauroidea, the tyrant dinosaurs. Best known from the later Late Cretaceous Asia and North American Tyrannosauridae, recent discoveries reveal a long history of tyrant dinosaurs going back into the Middle Jurassic.
Basal tyrannosauroid specializations include:
A newly-discovered clade of specialized (and often quite slender) theropods is Megaraptora. Their phylogenetic position has been extremely difficult to pin down: initially considered megalosauroids close to spinosaurids, they were then found as carcharodontosaurians (specially neovenatorids), but are now best placed as either primitive coelurosaurs in general OR tyrannosauroids closer to tyrannosaurids than are proceratosaurids OR as the oldest branch of Tyrannosauroidea (as shown in the detailed phylogeny above). Megaraptorans get their name from Megaraptor from the Late Cretaceous of Argentina (originally thought to be a possible coelurosaur, and popularly (on the Internet, although not in the scientific literature!) considered a giant dromaeosaurid raptor). Others include Aerosteon of the mid-Late Cretaceous of South America; Fukuiraptor of Early Cretaceous Japan; Australovenator of the late Early Cretaceous of Australia; and mid-Late Cretaceous Orkoraptor and Murusraptor of Argentina. Maip from the latest Cretaceous of southern Argentina is currently the youngest and largest known (definite) megaraptoran. The discovery of the snout of a juvenile Megaraptor shows that this clade had relatively long and slender skulls. Argentine Gualicho and Aoniraptor (possibly the same taxon!) may also be megaraptorans; unlike other members of this clade, but like derived tyrannosauroids, their manus is reduced to two fingers. In Australasia and in southern South America the megaraptorans seem to be the apex predators, especially towards the end of the Cretaceous. Some studies find that the (normally considered carcharodontosaurian) Siats and Chilantaisaurus of the mid-Cretaceous represent enormous basal megaraptorans.
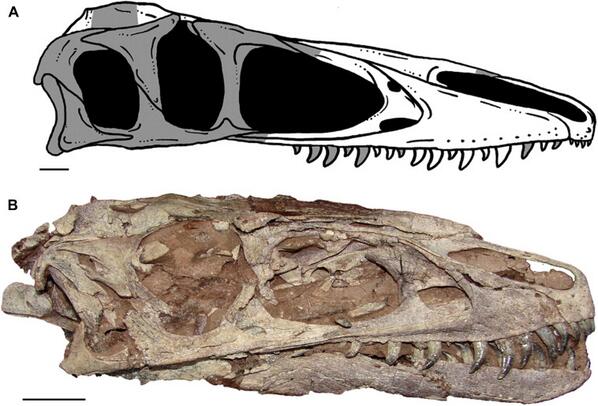
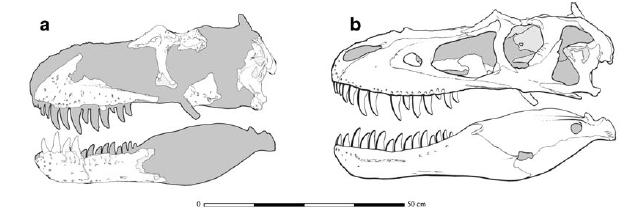
The oldest and most primitive (other than the coelurids) among the tyrannosauroids are the Proceratosauridae. The oldest known are Middle Jurassic English Proceratosaurus and equally old Siberian Kileskus. Slightly younger (and far more completely known) is Guanlong of the early Late Jurassic of China: a 3 m or longer crested proceratosaurid. Like other early coelurosaurs, the arms were fairly long. Long arms with tridactyl manus were likely found in all proceratosaurids, but we do not yet have the arms for Proceratosaurus, Kileskus, or Early Cretaceous (and possibly 8 m long!) Sinotyrannus, one of the youngest known proceratosaurids. Of similar age (and possibly the same genus) is Yutyrannus of Early Cretaceous of China. Known from three nearly-complete fossils, this is a 9 m long predator. It still retained the tridactyl manus with long claws of typical tetanurines. Because they were preserved in fine-grained ash-based sediments, the remains of long protofeathers were found over its body, indicating that even giant theropods had a fuzzy coat!
Dilong of the Early Cretaceous of China represents the next phase of tyrannosauroid evolution, the Pantyrannosauria. It was the first tyrannosauroid discovered with protofeathers. At 1.5 m length, it still indicated that some basal tyrannosauroids were small members of the predatory community.
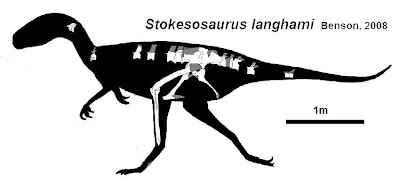
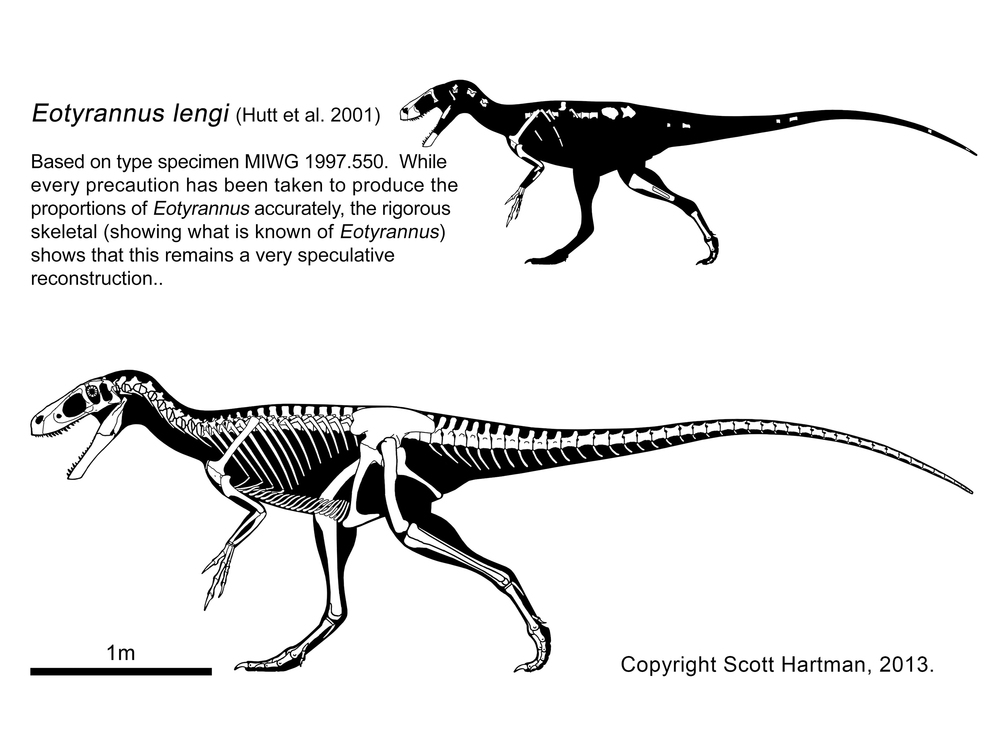
More robustly-built are the Stokesosauridae, including Jurassic tyrannosauroids such as Stokesaurus of North America, and Juratyrant (formerly considered a species of Stokesosaurus) of Europe, and Aviatyrannis of both (however, there is some evidence that Aviatyrannis is actually a Jurassic ornithomimosaur). A later stokesosaurid is Early Cretaceous Eotyrannus of Europe, with an adult size of possibly 4.5 m or more. It was dwarfed by other theropods in its community: the carnosaur Neovenator and the spinosaurid Baryonyx. (A new analysis suggests that stokesosaurids actually branched off below the Coleuridae and Proceratosauridae, and that Eotyrannus was actually a more derived tyrannosauroid closer to Xiongguanlong and later forms.)
Xiongguanlong of late Early Cretaceous China and Timurlengia of early Late Cretaceous Uzbekistan show an increase in body size and the development of increased bite forces.
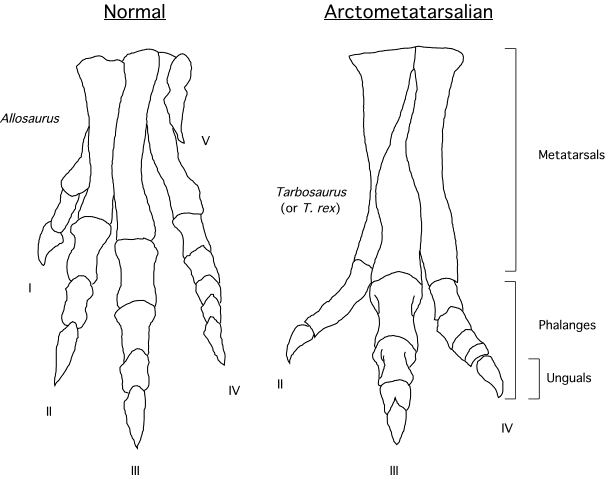
Two small-bodied tyrannosauroids with arctometatarsi are known from the early Late Cretaceous of western North America: incompletely known Moros and more completely known Suskityrannus. Both lived in environments when other forms of theropods were the dominant apex predators. While the arms of proceratosaurids and stokessaurids were primitively long, those in Suskityrannus show the later tyrannosauroid trend of reduced arm length. Similarly, these forms show elongation of the distal hindlimb and the development of the arctometatarsus, traits shared with their more derived relatives.
The more derived Eutyrannosauria increase size again. One branch of eutyrannosaurs is the eastern North American 6 m or longer slender-footed Dryptosauridae, best-known from Dryptosaurus. In Dryptosaurus the arm is very short but has a very large claw. It is confirmed as having an arctometatarsus. As with more derived tyrannosauroids, the distal hindlimbs (tibia, metatarsi) are elongated: an indication of cursorial (running) ability. Another Eastern North American eutyrannosaur is Appalachiosaurus of the Late Cretaceous of eastern North America. About the same age is deep-skulled Bistahieversor of the Late Cretaceous of the American Southwest. But the best understood and most derived eutyrannosaurs were Tyrannosauridae proper.
TYRANNOSAURIDAE
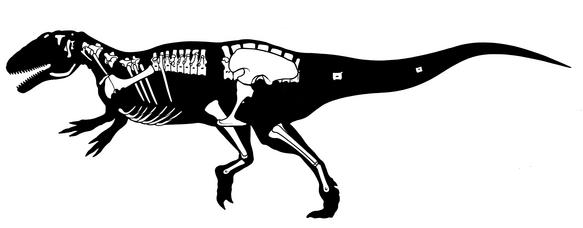
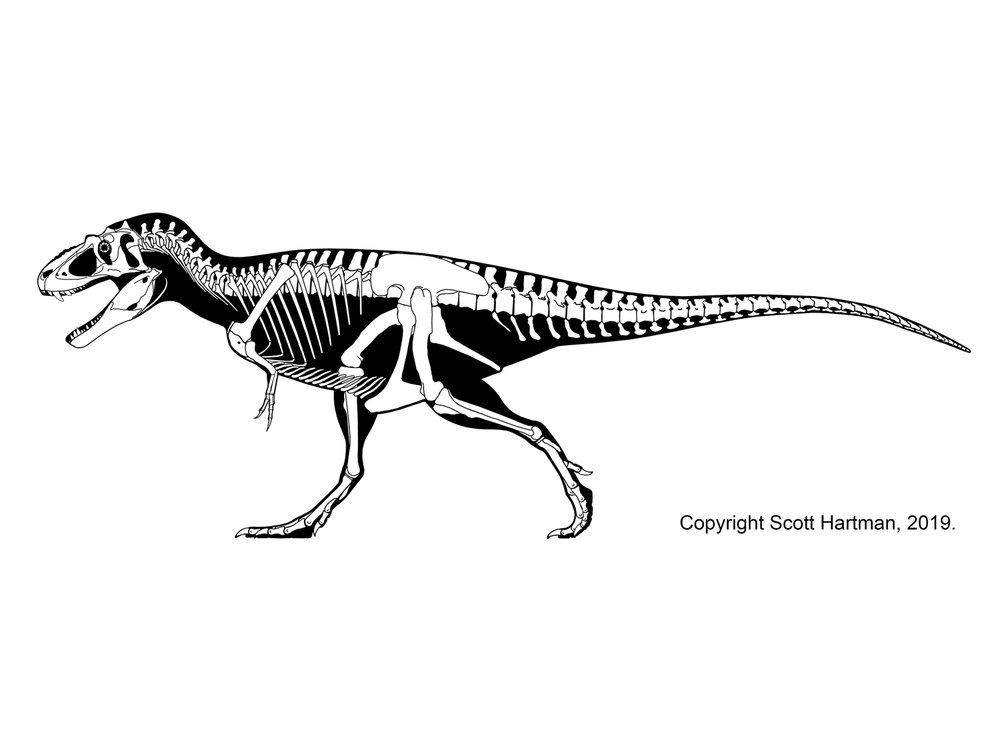
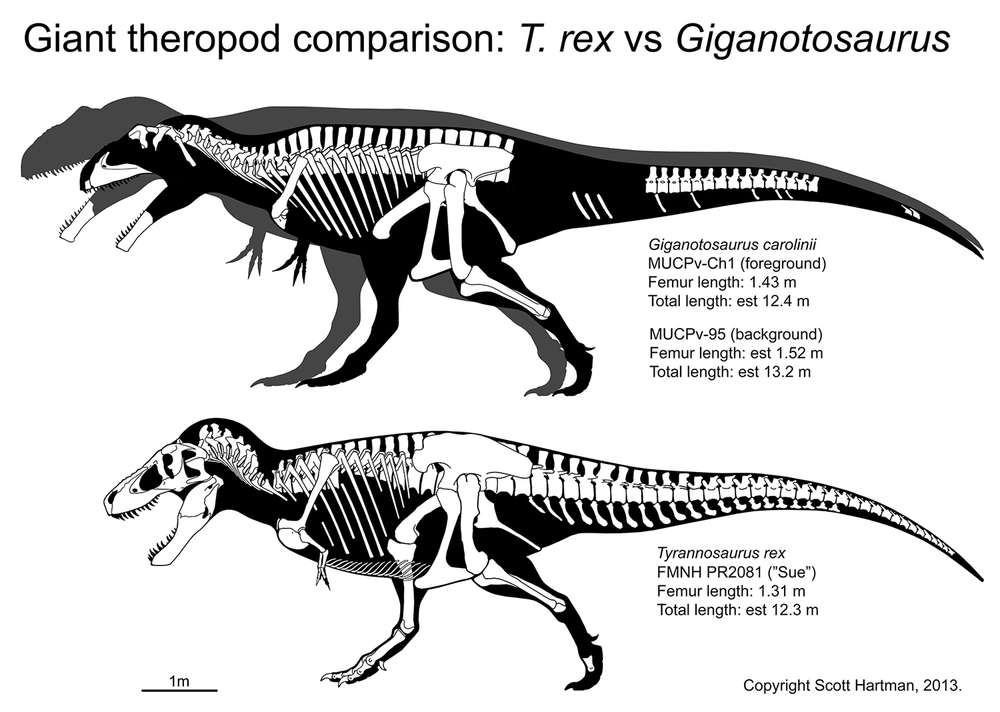
Tyrannosaurids proper are one of the last groups of large bodied theropods to evolve, showing up only in the last 20 million years or so of the Late Cretaceous of North America and Asia. (Consequently they have a similar range distribution to coronosaur ceratopsians, pachycephalosaurids, corythosaur-line lambeosaurines, and club-tailed ankylosaurines). Although for most of their history tyrannosauroids were minor predators in their habitats, tyrannosaurids were by far the largest flesh-eaters in their environments. Small tyrannosaurids were about 8 m long; most reached at least 10 m; and at least one genus reached 13 m.
Tyrannosaurids were specialized relative to their ancestors by possessing:
Tyrannosaurids include the relatively slender Albertosaurus and Gorgosaurus of western North America; slender long-snouted Alioramus and Qianzhousaurus of Asia; and more heavily built Lythronax, Daspletosaurus, and Teratophoneus of western North America and Tarbosaurus and Zhuchengtyrannus of Asia; and giant 13 m long, 8-10 ton Tyrannosaurus of western North America. Tyrannosaurus rivals the biggest carcharodontosaurs and spinosaurids in mass.
Tyrannosaurids seem to have relied solely on their jaws to kill their food. Their long legs meant that they were faster than their potential prey (hadrosaurids, ceratopsids), although adults of the 2 ton or greater size range may not have been fast runners. (Juvenile tyrannosaurids, though, would have been among the fastest dinosaurs). At least some tyrannosaurids have been found in groups of different ages: possibly family associations.
In communities where tyrannosaurids were present, mid-sized predatory dinosaurs were rare or absent. Furthermore, the differences in tooth form, skull shape, relative arm length, and hindlimb proportions strongly suggest that young tyrannosaurids fed upon different prey and hunted in a different style than the adults, as I discuss in this video:
MANIRAPTORIFORMES: Plant-Eating "Carnivorous" Dinosaurs
The remaining coelurosaurs (Maniraptoriformes) all have brains that are twice again as large or larger (based on skull size) as the more basal coelurosaurs. They also share a suite of unusual features that strongly suggest a move away from the strictly carnivorous diet of their ancestors and relatives. In particular, they typically:
A new discovery of broad pennaceous feathers (i.e., feathers with a shaft, branches off of that, subbranches off of that, etc.) in ornithomimosaurs seem to place this trait at the same part of the tree as this shift to non-carnivory. However, at present they are only known on the arms of ornithomimosaurs: the spread to other parts of the body seems to be further up the tree.
ORNITHOMIMOSAURIA
With the possible exception of Ornitholestes, the basalmost lineage of the maniraptoriforms are the Ornithomimosauria, the ostrich dinosaurs. Ornithomimosaurs differ from the ancestral state by:
Their adaptations suggest a move away from predation towards a more omnivorous or even herbivorous lifestyle.
Primitive ornithomimosaurs are known from the Early Cretaceous of Africa (Nqwebasaurus: currently the oldest and most primitive known ornithomimosaur, and the only one from the Southern Continents), Europe (Pelecanimimus), and Asia (Harpymimus, Shenzhousaurus, and Hexing). The remaining ornithomimosaurs fall into two clades: Deinocheiridae and Ornithomimidae. Both of these are dinosaurs larger than the basal members, and possess toothless beaks.
Deinocheirids are currently only known from Asia. They include large of the Early Cretaceous Beishanlong), Late Cretaceous of Asia (Garudimimus, and truly gigantic Deinocheirus, long known only from its arms and a few isolated bones (but new discoveries give us a better sense of the animal), is a Tyrannosaurus-sized primitive ornithomimosaur (lacking an arctometatarsus). The new discoveries show that Deinocheirus had a spinosaur-like sail over the hip region, an expanded blunt snout, a deep jaw, highly reduced supratemporal fenestrae, and oddly blunt toes. Deinocheirids retain the ancestral limb proportions of most theropods. The belly contents of Deinocheirus include numerous fish bones and scales, so it was probably omnivorous. At 6.4 tonnes, it was among the very largest theropods.
The Ornithomimidae did not produce any forms this large (although the largest rival Beishanlong). At present they are only known from the Late Cretaceous of Asia and North America. Ornithomimids are characterized by arctometatarsus (convergently evolved with Tyrannosauridae). Early Cretaceous Kinnareemimus of Thailand shows an incipient arctometatarsus form and may turn out to be the basalmost ornithomimid. Otherwise, the oldest and most primitive is Sinornithomimus of the early Late Cretaceous of China. Other ornithomimids include western North American Struthiomimus, Dromiceiomimus, and Ornithomimus, and Asian Gallimimus and Anserimimus. These dinosaurs were among the most cursorial of all theropods.
At least some ornithomimosaurs lived in herds/flocks. Recent discoveries reveal that adult ornithomimosaurs had pennaceous feathers on the arms and tail, but juveniles do not seem to have this.
EVOLUTIONARY PATTERNS IN BASAL COELUROSAURS
Feeding adaptation transformations:
Locomotory adaptations:
Gigantism and Miniaturization:
Niche partitioning:
Basal coelurosaurs represented the minor predators of many Jurassic and Early Cretaceous environments. Diversification into non-predatory modes allowed coelurosaurs to diversify into niches previously unoccupied by theropods. (In Late Cretaceous Asia, small non-predatory coelurosaurs are very common, while small ornithopods are absent.) In contrast, tyrannosauroids evolve into giant top predators in the Late Cretaceous of Asia and North America after the disappearance of carcharodontosaurids and spinosaurids.
.jpg)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}