


Key Points:
•Therizinosauria had long arms with large clawed hands. The derived Therizinosauroidea within it have expanded guts, retroverted pubes, and shortened metatarsi, showing that they had evolved into slow-moving herbivores.
•Alvarezsauria were small-bodied insectivorous coelurosaurs. Derived forms (Alvarezsauridae) had highly transformed forelimbs that had become short but powerful picks.
•The remaining coelurosaurs formed the clade Pennaraptora, characterized by sideways-oriented shoulder joints, long arms with a specialized folding wrist, and broad pennaceous feathers on the arms and tail.
•Among the pennaraptorans were the omnivorous short-skulled Oviraptorosauria (including the tiny gliding Scansoriopterygidae and the toothless Caenagnathoidea).
•Eumaniraptora is a clade of (mostly) small-bodied feathered coelurosaurs. Based on their body size and feathered limbs, at least ancestrally they could probably fly (at least to some extent).
•Relationships among the eumaniraptorans has been difficult to determine with certainty, because the primitive members (especially the Late Jurassic forms) are all so similar in form.
•Deinonychosauria (the raptor dinosaurs) is characterized by a hyperextensible sickle claw. It contains within it the diverse Dromaeosauridae and the Troodontidae.
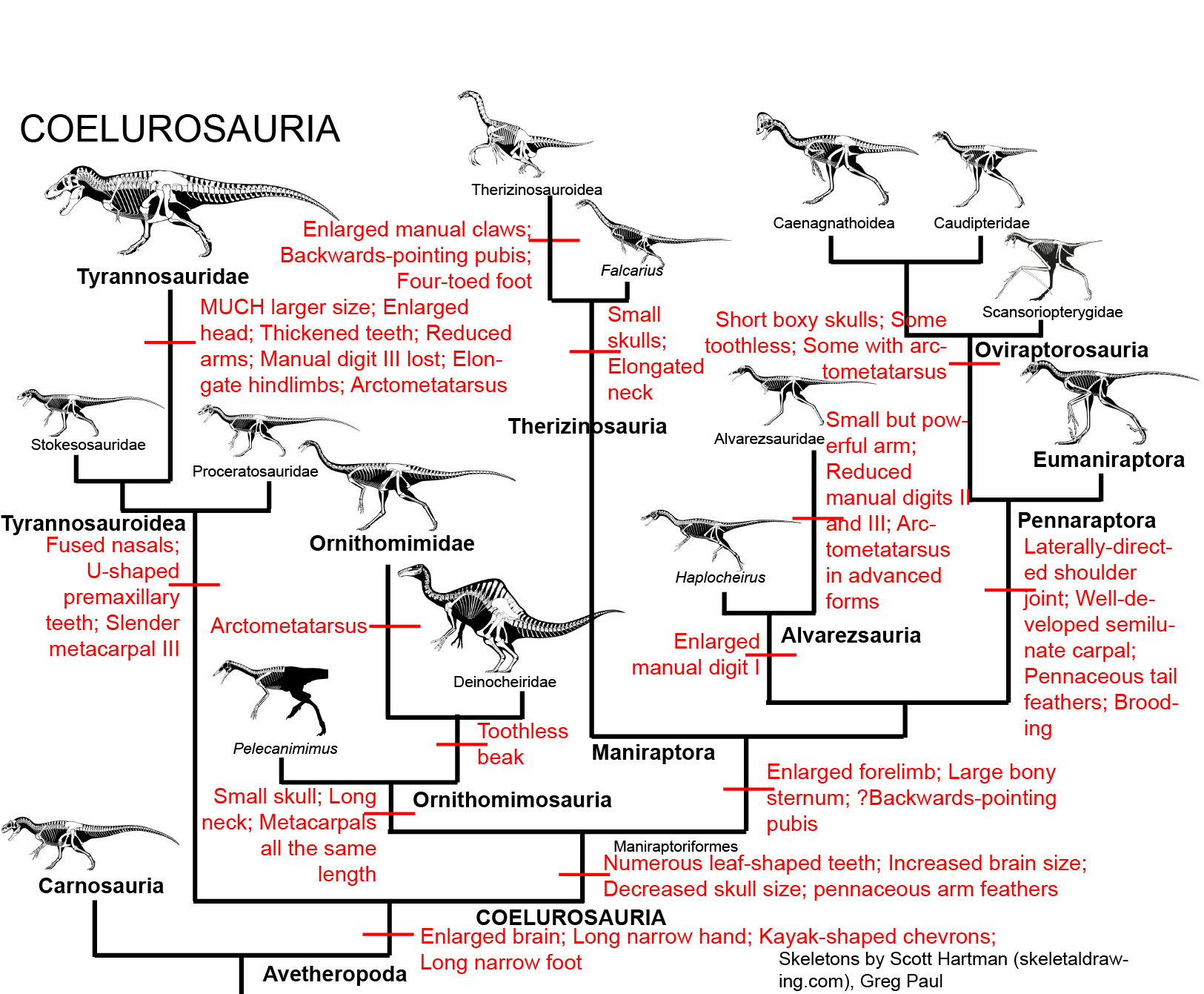
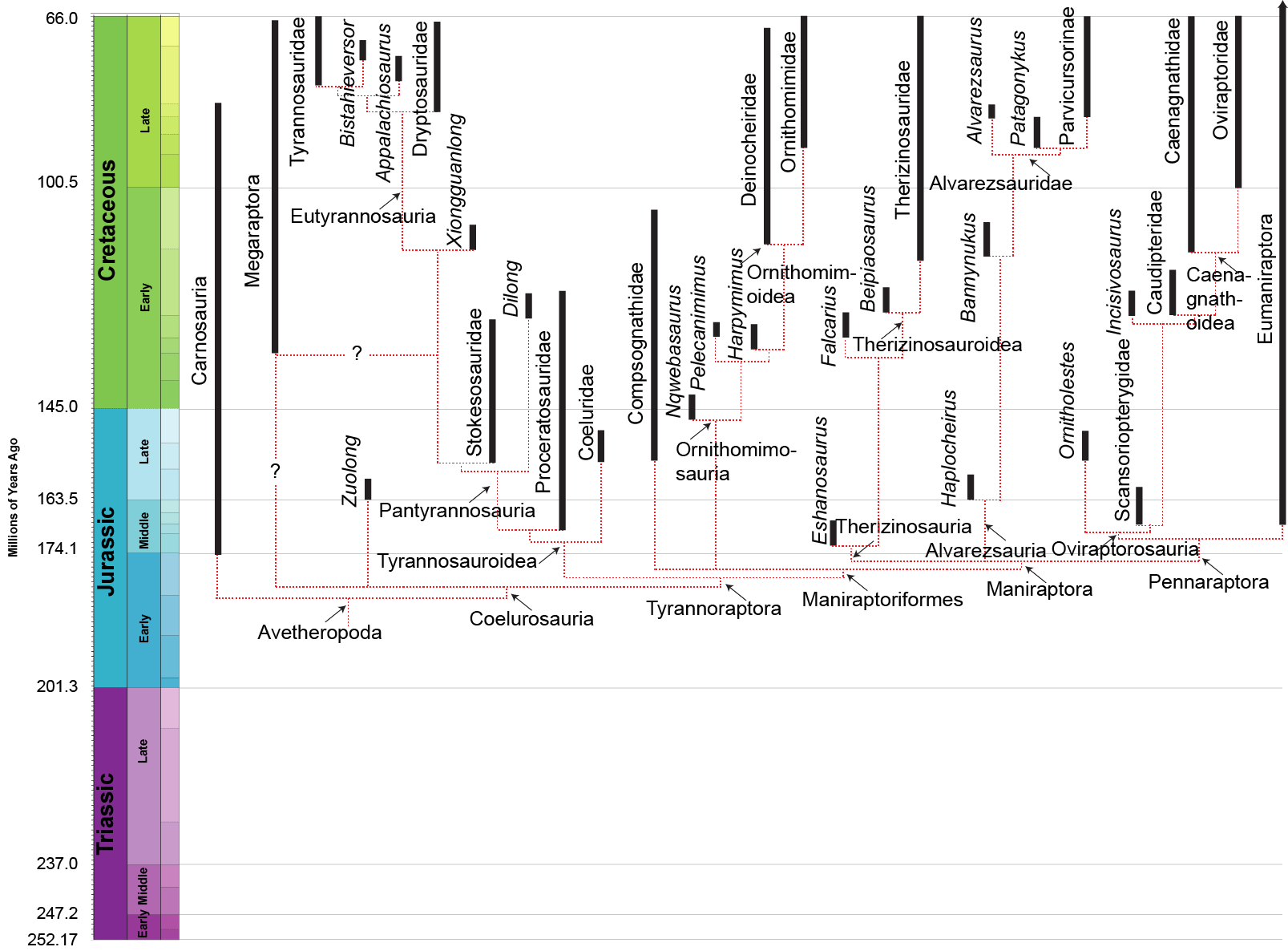
MANIRAPTORA
The remaining theropods form the clade Maniraptora ("hand grabbers"). Maniraptorans show numerous specializations:
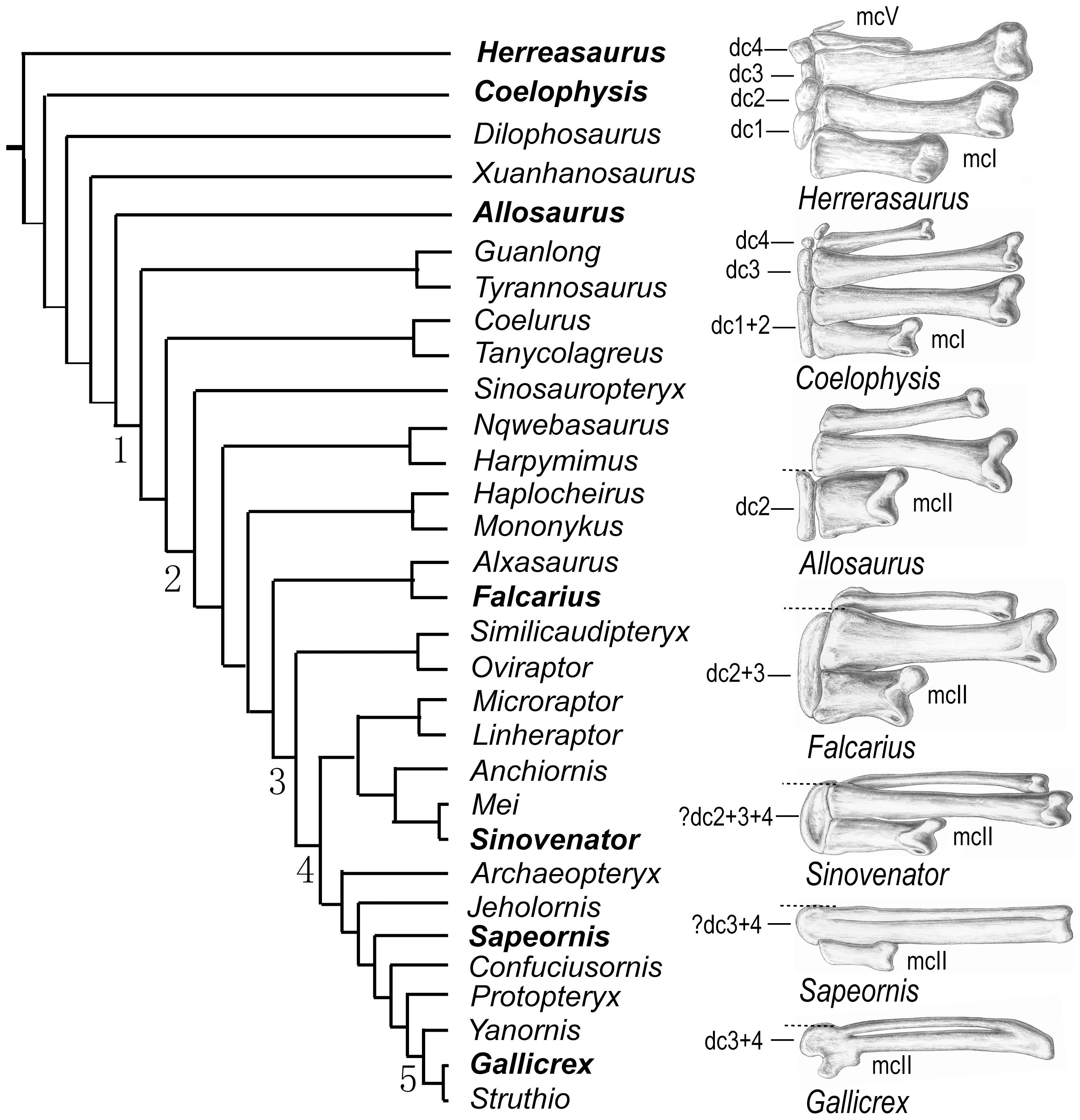
One possible problematic shared derived feature of Maniraptora is a backwards-pointing pubis. Most coelurosaurs (and saurischians in generally) have a vertically-oriented or anteriorly-oriented pubis. In therizinosauroids, alvarezsaurids, the basal troodontid Sinovenator and the derived troodontid Latenivenatrix, dromaeosaurids, Archaeopteryx, and avialians the pubis points backwards; in the basal therizinosaur Falcarius, the basal alvarezsauroid Haplocheirus, oviraptorosaurs, most troodontids, and the basalmost avialian Anchiornis it points vertically or anteriorly. So it is difficult to say which condition is found in the concestor of Maniraptora. (Regardless of the answer, there is a LOT of convergence going on!!)
Changes in the muscle attachments in the hindlimbs of maniraptorans show a switch from the femur-and-tail power stroke found in other dinosaurs (inherited from the early diapsids) to one where the flexion of the knee is more important.
Maniraptorans are the most diverse clade of dinosaurs. None retain a basal theropod form: indeed, very few retain the ancestral carnivorous condition. Major groups include the Therizinosauria, Alvarezsauria, Oviraptorosauria, and Eumaniraptora (which get their own lecture).
The oldest maniraptorans are some possibly Middle Jurassic eumaniraptorans, and definitely this clade is present by the Late Jurassic. A dentary from the Early Jurassic of China is considered by some authors to be a therizinosaur: however, it might simply be a derived sauropodomorph (which is honestly far more consistent with the stratigraphic range of other coelurosaurs, and with the known high diversity of various clades of sauropodomorph in the same unit.)
THERIZINOSAURIA
This group and the Oviraptorosauria were once thought to form their own clade (Oviraptoriformes). However, discovery of the primitive members of both Therizinosauria and Oviraptorosauria shows that many of the similarities between the derived members of these clades are convergences. More recent studies typically place therizinosaurs as the most basal branch of Maniraptora.
Therizinosauria ("scythe reptiles") have been considered sauropodomorphs and late surviving proto-ornithischians, but are in fact coelurosaurian theropods. Prior to the 1990s, they were often called the "segnosaurs." Similar to the ornithomimosaurs, this group is characterized by:
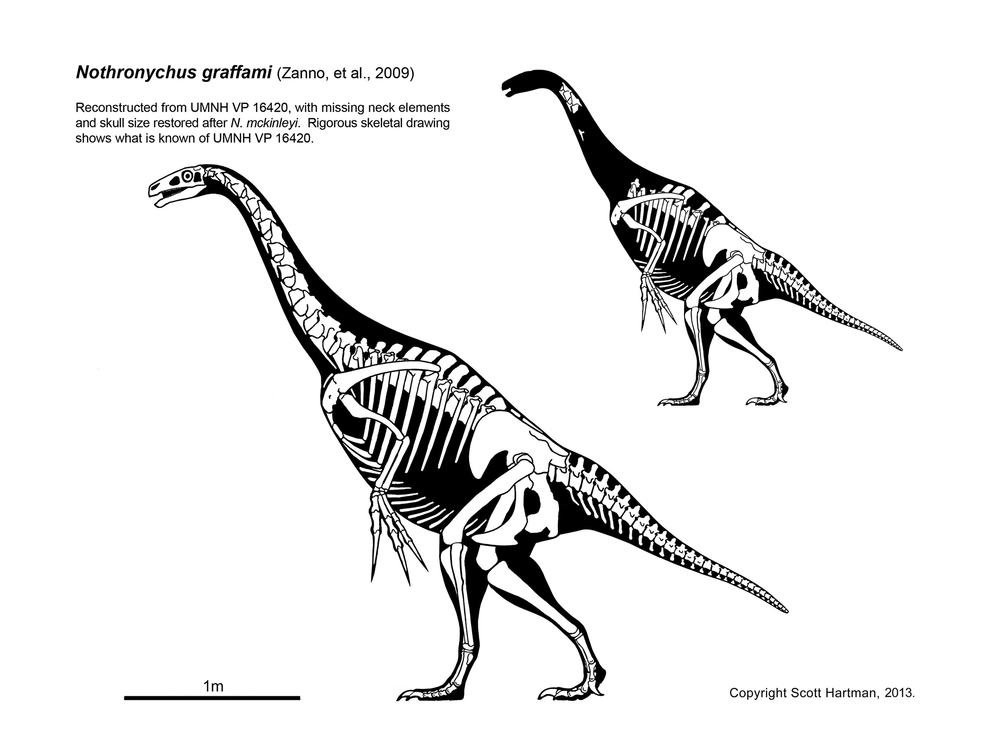
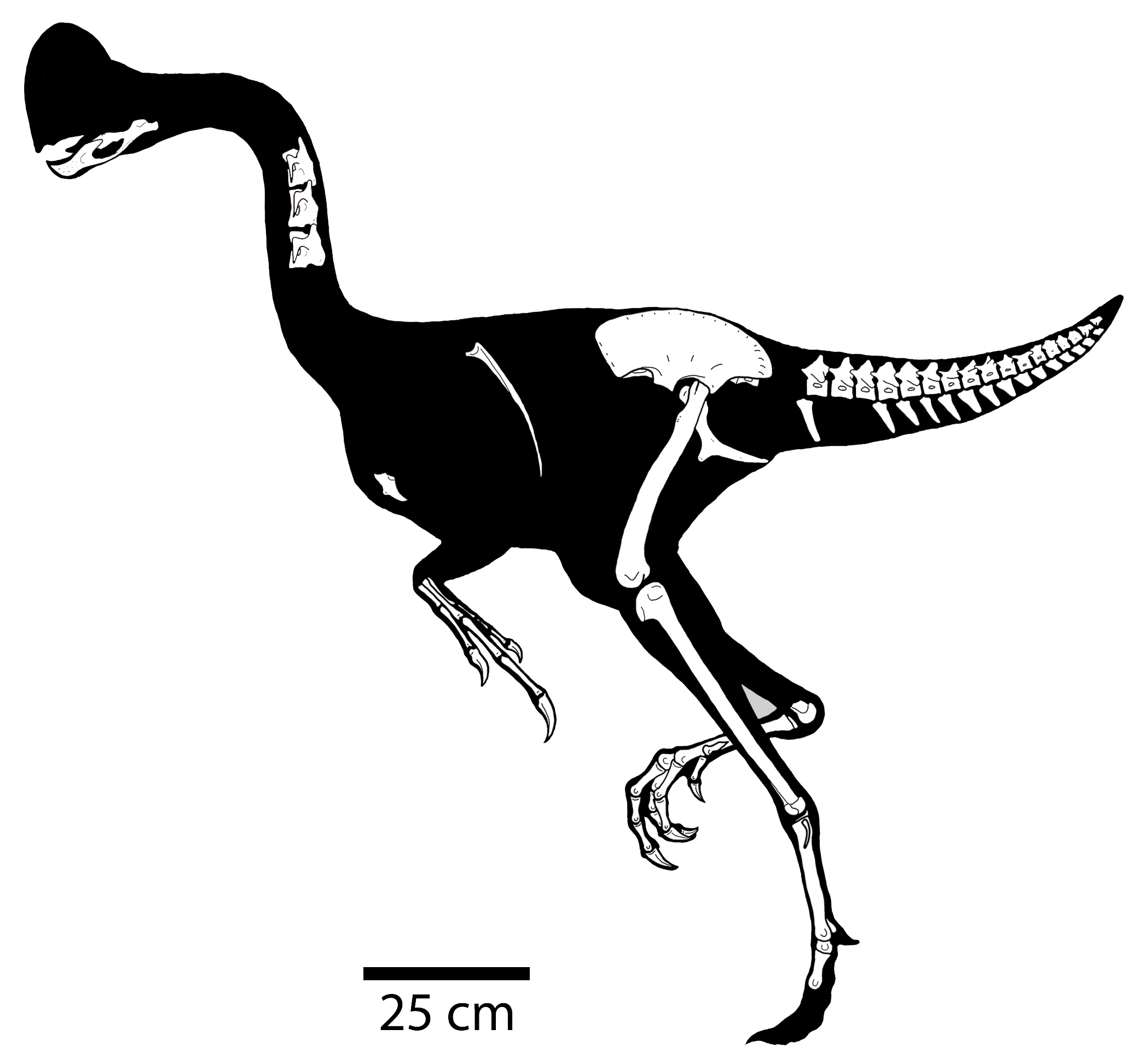
Other than the jaw Eshanosaurus, this group is known only from the Cretaceous, and only from Asia and North America at present. The basalmost form is Early Cretaceous Falcarius of western North America. It retains a relatively elongate metatarsus and a vertically-oriented pubis. The derived therizinosaurs form the clade Therizinosauroidea, and are characterized by shortened metatarsi in which all four toes touch the ground and backwards-pointing pubes. (In this case, like the ornithischians, this is almost certainly to accommodate a large gut for digesting plants.
Therizinosaurs seem to have been primarily, if not strictly, herbivores. Their stumpy feet and short legs show them to have been among the slowest theropods. To defend themselves (and possibly to help them feed) they had huge claws. They ranged from bear-sized taxa such as Erlikosaurus and Beipiaosaurus through Nothronychus to Tyrannosaurus-sized Therizinosaurus with 1 m long claws.
ALVAREZSAURA
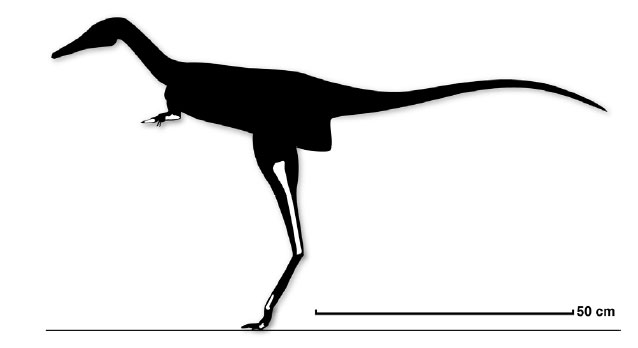
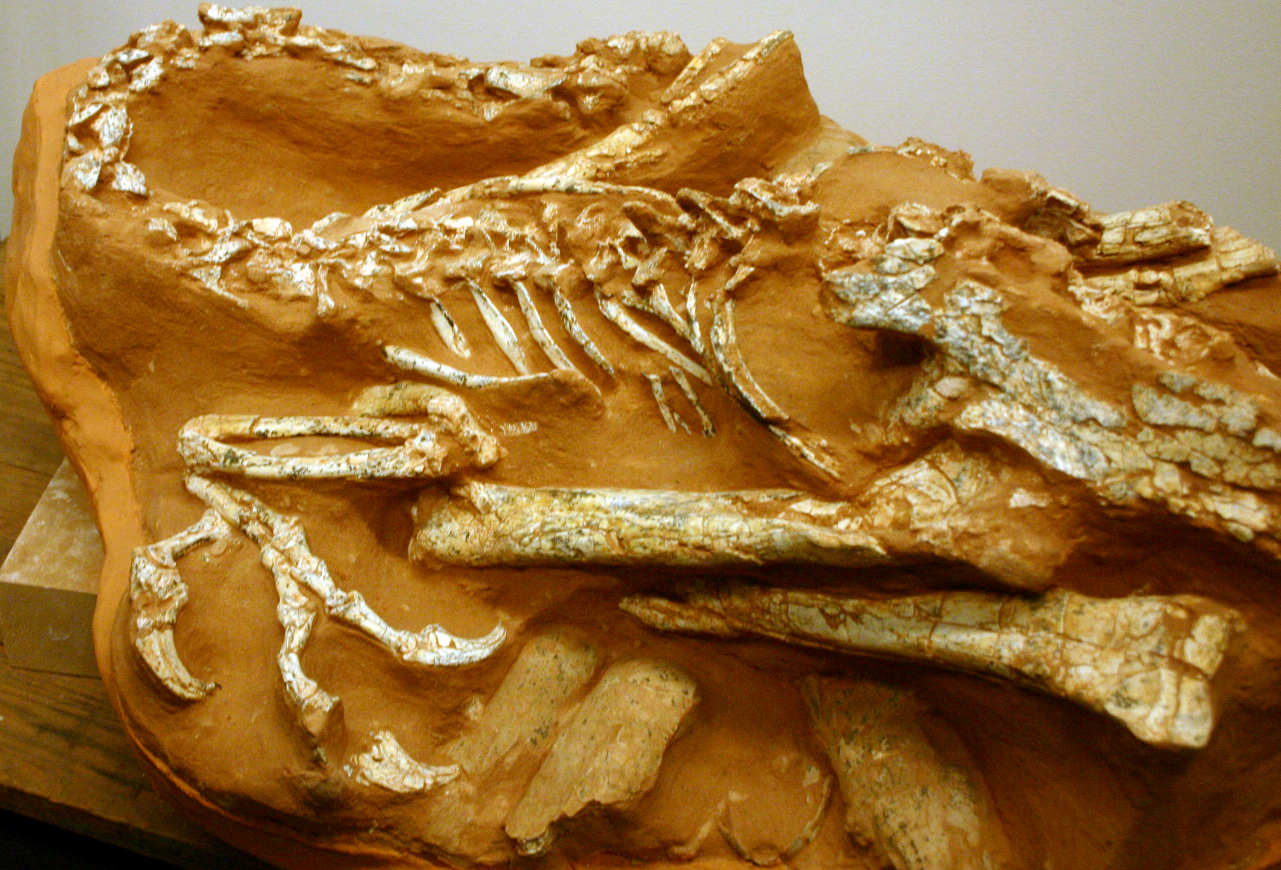
Alvarezsauria (sometimes called "Alvarezsauroidea") is a recently discovered, highly specialized group of maniraptoran theropods. Haplocheirus is the oldest well-known form, from the early part of the Late Jurassic of China. (Poorly-known Aorun from the latest Middle Jurassic might actually be an even more primitive alvarezsaur.) Until 2018, all the remaining alvarezsaurs (collectively the Alvarezsauridae) are from the Late Cretaceous. However, recently described are Early Cretaceous Xiyunykus and Bannykus, which document two different stages in transition relative to their Late Cretaceous relatives. Alvarezsaurids are known from South and North America, Europe, and Asia. They have numerous bird-like features, and were once thought to have been specialized flightless birds. Alvarezsaurids range in the chicken-to-rhea sizes.
Alvarezsaurids have small beaky skulls with tiny teeth and hands in which the thumb is much more powerful than the other fingers.
In the Cretaceous Alvarezsauridae, the forelimbs are further transformed into bizarrely powerful arms with a huge thumb claw and exceedingly small digits II and III. The alvarezsaurids have a backwards pointing pubis. Unlike the therizinosauroid and ornithischian situation, this backwards position of the pubis is more likely associated with changes in the locomotory muscles towards knee-driven power from the ancestral tail-and-femur driven power.
Only a little is known of Alvarezsaurus itself (the basalmost form); somewhat more is known for the more derived Patagonykus and Achillesaurus (all from South America). Only a foot is known of rhea-sized Kol of Asia.
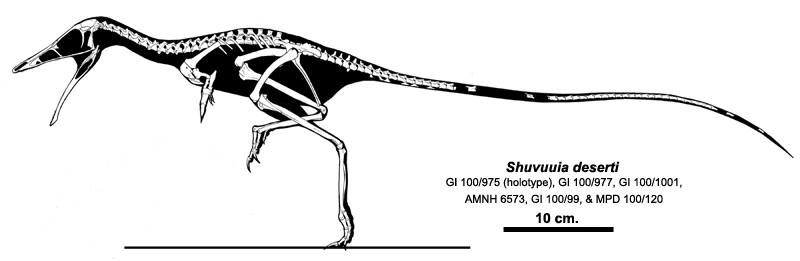
The highly derived Parvicursorinae (also called "Mononykinae"), in contrast, are known from many excellent specimens. The best studied are the Asian taxa Mononykus, Parvicursor, and Shuvuuia. More fragmentary Asian parvicursorines include tiny Albinykus, Linhenykus (in which digits II and III had entirely vanished), and Xixianykus. (However, North American forms such as Albertonykus and Trierarchuncus are known). Parvicursorines have an extreme version of the arctometatarsus, in which the upper portion of metatarsal III is entirely missing.
The parvicursorines show numerous cursorial adaptations (including a modified tail that allowed them to take quite sharp turns), but these were almost certainly defensive. They seem to have been insectivores, and their forelimbs may have been used to batter into ant and termite nests. They have been found from deserts to well-watered environments.
PENNARAPTORA: Fully Feathered Dinosaus
The remaining maniraptorans form the clade Pennaraptora ("feathered raptors"). These comprise the oviraptorosaurs, the scansoriopterygids, and the eumaniraptorans. These groups are united by several important characteristics:
OVIRAPTOROSAURIA
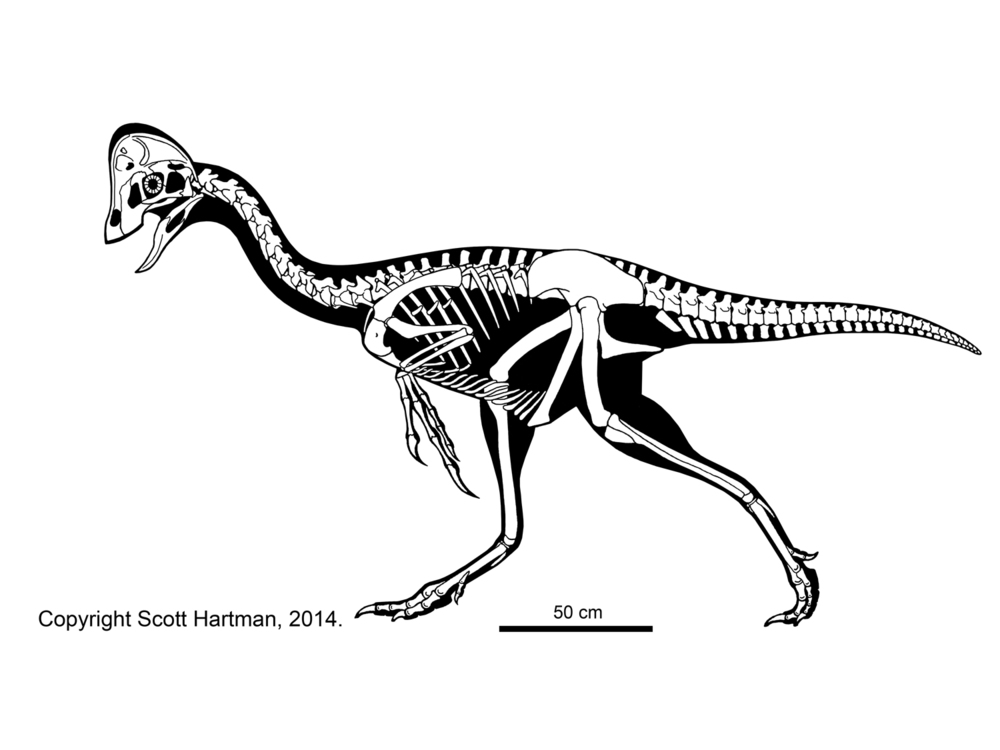
Oviraptorosauria is characterized by
An unusual clade of small pennaraptorans (and likely basal oviraptorosaurs) are Scansoriopterygidae. Known from Middle-early Late Jurassic Epidendrosaurus (also known as "Scansoriopteryx") and Epidexipteryx (which may just be the adult form of Epidendrosaurus!), and spectacular Yi and Ambopteryx these are among the smallest Mesozoic dinosaurs (pigeon-sized). (Early Cretaceous Zhongornis was once thought to be scansoripterygid, but is most likely a basal avialian.) As they are small enough, and have the distally-placed pedal digit I, it may be that they may have spent some time up in the trees. The skull shape and tiny teeth of the scansoriopterygids suggest that they might have been insect eaters or omnivores. The first phylogenetic studies place these dinosaurs as the basalmost members of Avialae, but newer analyses placed them outside Eumaniraptora proper, and the most recent place them as basal members of Oviraptorosauria (which would solve the problem of the lack of pre-mid-Cretaceous oviraptorosaurs.) Most specimens are juveniles, but Yi seems to be from an adult.
Very bizarrely, Yi shows an extra bone projecting from its wrist, and a membrane attached to this. Such accessory bones attached to membranes have evolved conversantly in some gliding and flying mammals. These were confirmed in Ambopteryx. This suggests that Yi (and maybe other scansoriopterygids) were flying squirrel-like gliders.
Among the rest of Oviraptorosauria, the basalmost are toothy Incisivosaurus, Protarchaeopteryx, and the Caudipteridae (sometimes spelled "Caudipterygidae": Similicaudipteryx and Caudipteryx), all from the Early Cretaceous of China. All these forms are about the size of a modern turkey, which makes them all much larger than the sparrow-to-mockingbird-sized scansoriopterygids.
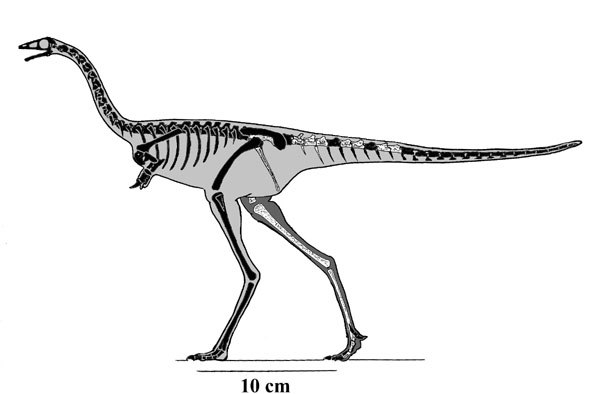
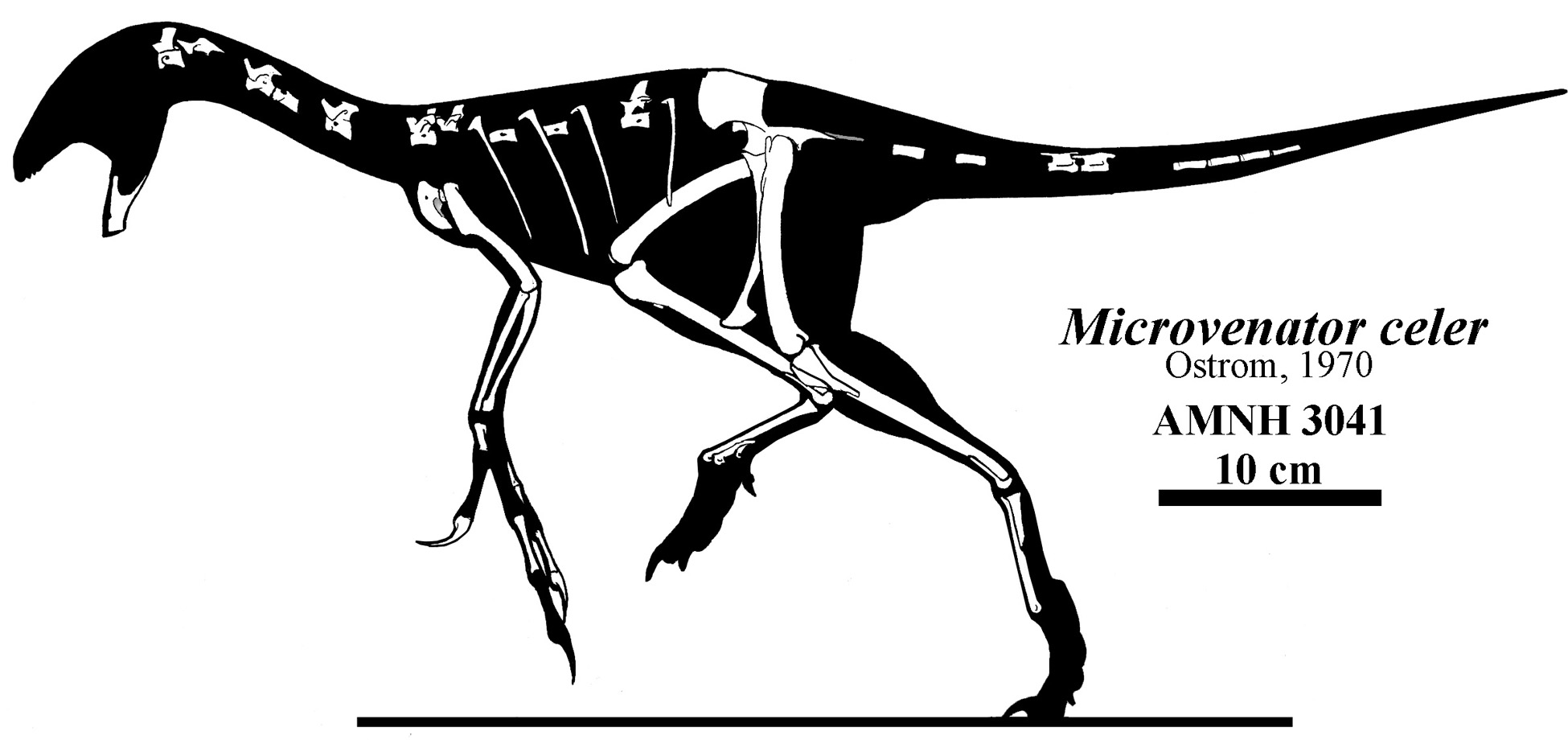
Other early branches of the oviraptorosaurs are Microvenator (probably a caenagnathid) of the Early Cretaceous of western North America and specialized Avimimus (with an arctometatarsus) of the Late Cretaceous of Asia. (The latter seems to have dwelt in herds/flocks/whatever).
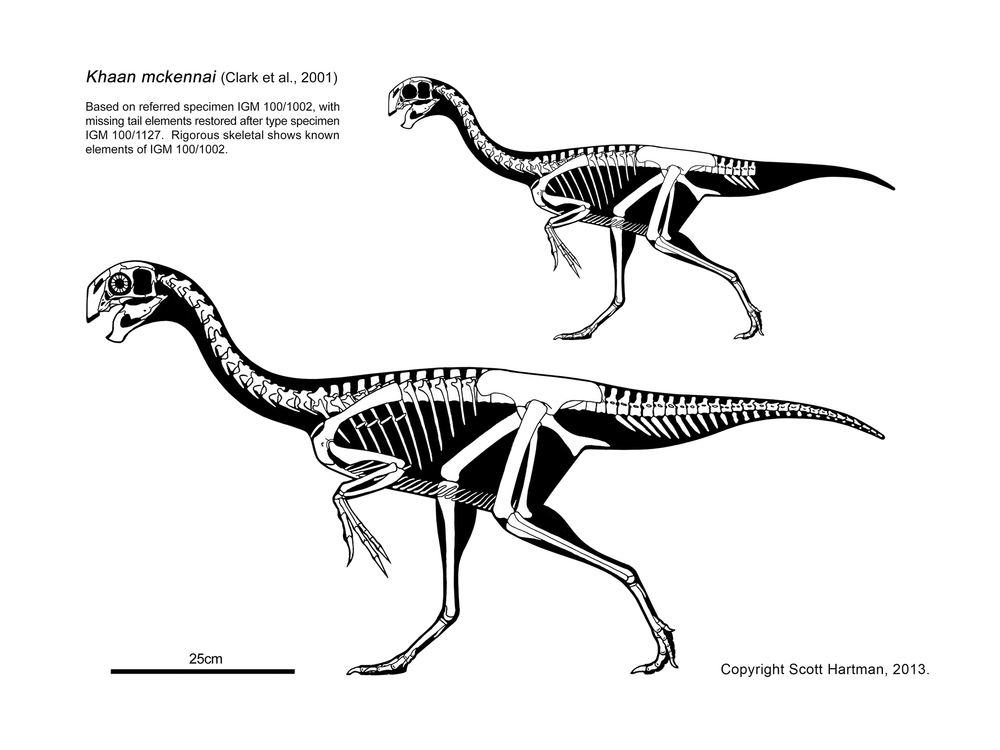
These basal branches of Oviraptorosauria are relatively small (sparrow-to-turkey sized). The more derived Caenagnathoidea contain forms that range from turkey to human to tyrannosaur size. Caenagnathoids are derived by loss of all teeth as well as other specializations. Most (but not all) phylogenetic studies divide the Late Cretaceous caenagnathoids into two branches: the Caenagnathidae (sometimes called the "Elmisauridae" or "Elmisaurinae": taxonomy on this remains in flux) which had slender mandibles, elongate hindlimbs, and sometimes even an arctometatarsus, and the deep-jawed, stout-footed Oviraptoridae. However, other studies mix and match the components of these groups. Oviraptoridae proper seems to be limited to Asia, while Caenagnathidae is found in both Asia and North America. Examples of oviraptorids include crested Oviraptor, Citipati and Rinchenia and Oksoko and crestless Khaan. Caenagnathids include Asian Elmisaurus and Caenagnathasia, and larger North American forms such as Chirostenotes, Leptorhynchos, Epichirostenotes, Apatoraptor, Hagryphus, and Anzu. By far the largest oviraptorosaur is the recently discovered caenagnathid Gigantoraptor of Asia: as large as an Albertosaurus or other mid-sized tyrannosaurid. Egg and some skeletal evidence points to the existence of Gigantoraptor-sized oviraptorosaurs in early Late Cretaceous North America.
The life habits of oviraptorosaurs are confusing. While the ancestral ones seem to be convincingly herbivorous, there are lizards in the gut contents of some oviraptorids: perhaps they were omnivorous? Many oviraptorosaurs have been found in desert environments, but others in forested regions.
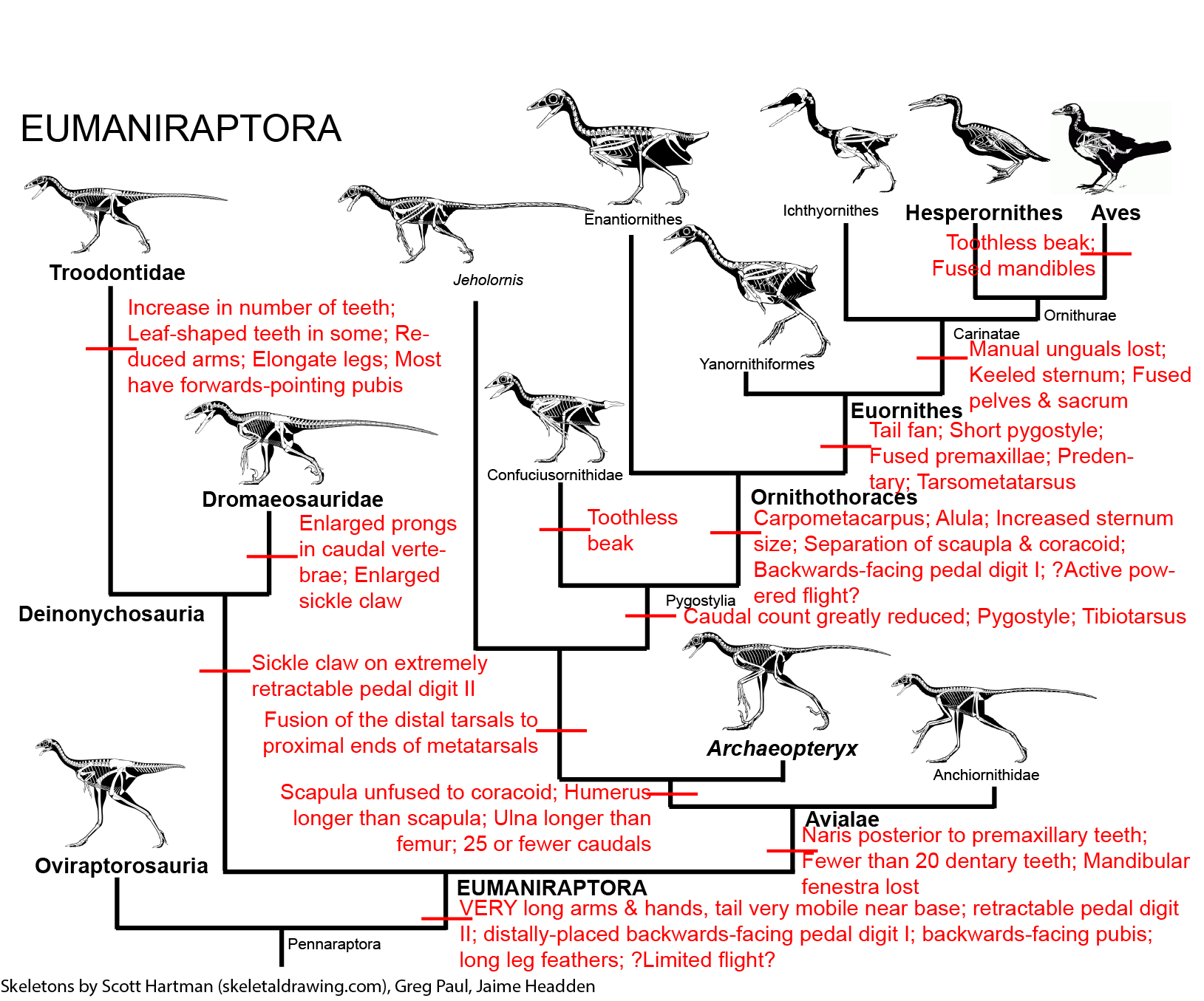
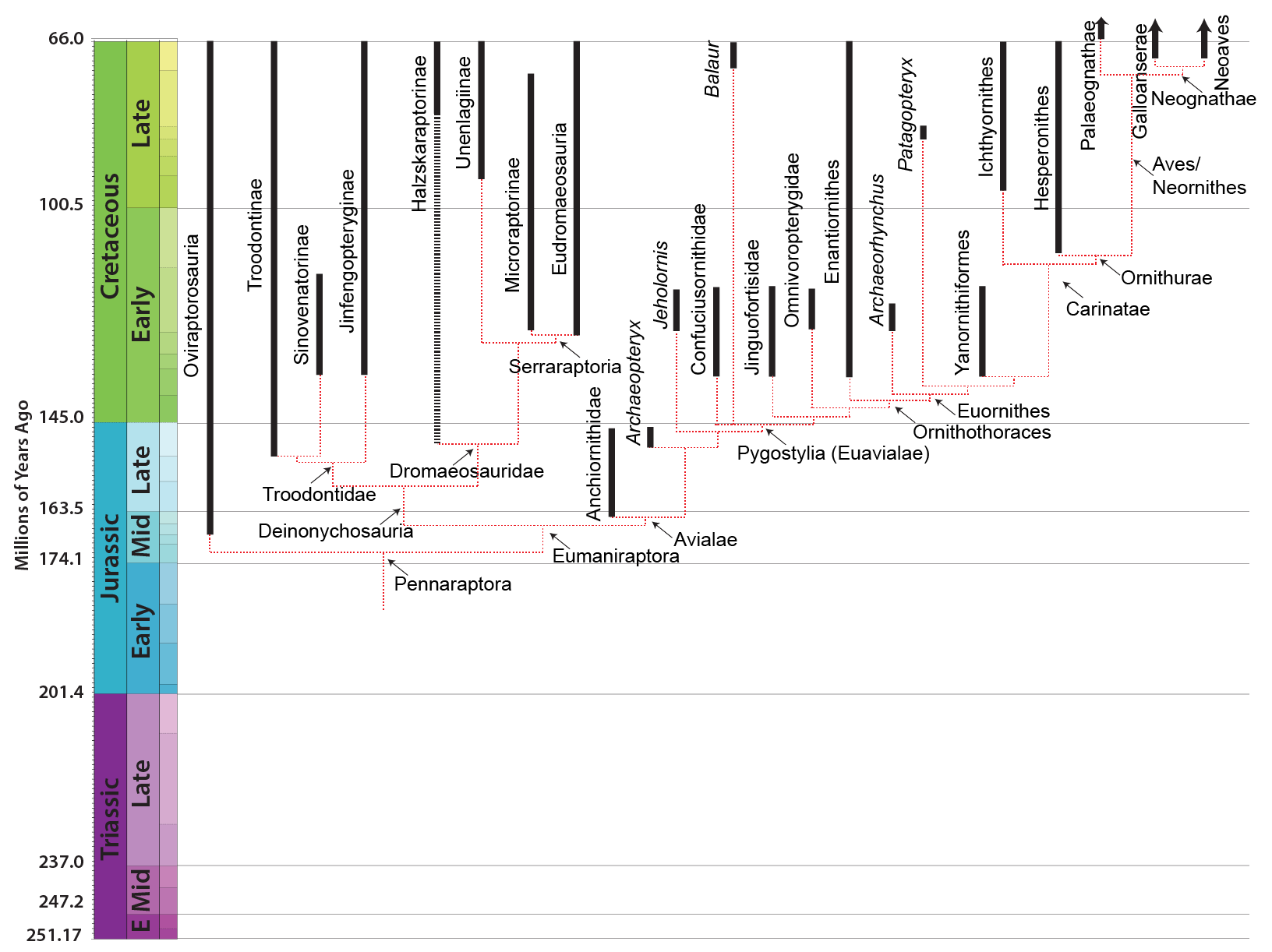
MAJOR GROUPS OF EUMANIRAPTORANS
Eumaniraptora is just one clade within Pennaraptora (and thus Maniraptora). The actual shared derived characters that unite them as a whole are a bit problematic, due to the uncertainty of the position of Archaeopteryx and other Jurassic taxa and the discovery that basal dromaeosaurids and troodontids are far more Archaeopteryx-like than are the "classic" dromaeosaurids and troodontids. Some probable transformations at the base of Eumaniraptora include:
Is "Bird" Really the Word?: A note on the use of the word "bird": this English term unfortunately carries with it a lot of intellectual baggage. If we hear that a fossil was from a "bird" we might assume that it had modern "bird" habits and adaptations (flight, song, tree nests, seed or insect diets, etc.). However, not all these characteristics are even true of all modern birds, much less of earlier branches of the lineage. So let's avoid calling Archaeopteryx a "Jurassic bird". There is no evidence that it was a seed eater; there is no evidence for song; it almost certainly nested on the ground; and as we will see it might not have been able to fly any better than early dromaeosaurids.
Eumaniraptora is traditionally divided into two clades: Deinonychosauria and Avialae. However, this situation has become complicated. On the one hand, there are several primitive eumaniraptorans whose exact placement is uncertain. For example, it is uncertain where the late Middle Jurassic (or early Late Jurassic) Pedopenna fits. It might be a basal avialian, or it might be outside Eumaniraptora. Similarly, the scansoriopterygids (discussed above) may be primitive avialians.
But more problematically, some new studies fail to recover the traditional grouping of Dromaeosauridae plus Troodontidae as a monophyletic "Deinonychosauria". And even worse, these studies disagree if it is Troodontidae or Dromaeosauridae which closer to Avialae, or even if Dromaeosauridae is monophyletic. (Furthermore, some studies in the 2010s pointed to an "Archaeopterygidae" comprised of Archaeopteryx and taxa here regarded as anchiornithids, and this whole group was uncertain as to where they fit.)
One comforting aspect of this uncertainty: the similarity of the morphology of anchiornithids, basal troodontids, microraptorine dromaeosaurids, and Archaeopteryx, shows us what our best model for what the common ancestor of all Eumaniraptora looked like. (Basically, something very similar to Anchiornis and Archaeopteryx.) The basal members of all three clades are crow-sized animals.
DEINONYCHOSAURIA
For many decades the "raptor dinosaurs" broadly defined (Troodontidae and Dromaeosauridae) were grouped together as Deinonychosauria ("Deinonychus lizards"). Some studies fail to recover this clade, and instead have either Troodontidae or Dromaeosauridae closer to birds. (In fact, some analysis find both groups paraphyletic with respect to birds!)
Deinonychosauria is united by a few traits, including features of the mandible. However, for this course we will focus on one in particular (and indeed one of the first features noted to be shared among them): a sickle claw on the extremely retractable pedal digit II.
At present all known deinonychosaurs are either dromaeosaurids or troodontids. However, some analyses in the past found archaeopterygids and/or anchiornithids as a basal deinonychosaur clade.
At present there is only one definite deinonychosaur from the Jurassic: the troodontid Hesperornithoides of western North America. Some additional teeth and skeletal material from the Late Jurassic of western North America and from the Middle Jurassic of England have been referred to either Troodontidae or Dromaeosauridae, but we can't definitely show these aren't from a basal avialian group instead. That said, two-toed footprints from the Middle Jurassic of Iran are pretty convincing evidence that deinonychosaurs were present.
DROMAEOSAURIDAE
Basal dromaeosaurs (Unenlagiinae and Microraptorinae) include some crow- to turkey-sized taxa: Rahonavis of the Late Cretaceous of Madagascar (and initially thought to be a bird); Microraptor of the Early Cretaceous of China (the first eumaniptoran for which the tail feathers were known); long-snouted Buitreraptor of Late Cretaceous Argentina; and others. Not all of these were small, however. One of the most unusual of these basal dromaeosaurids is giant Austroraptor of the mid Late Cretaceous of Argentina: a long-snouted, short-armed polar bear-sized unenlagiine. The very small size of unenlagiine teeth suggest that they ate only small bodied animals; we have definite evidence that at least some microraptorines ate their avialian cousins.
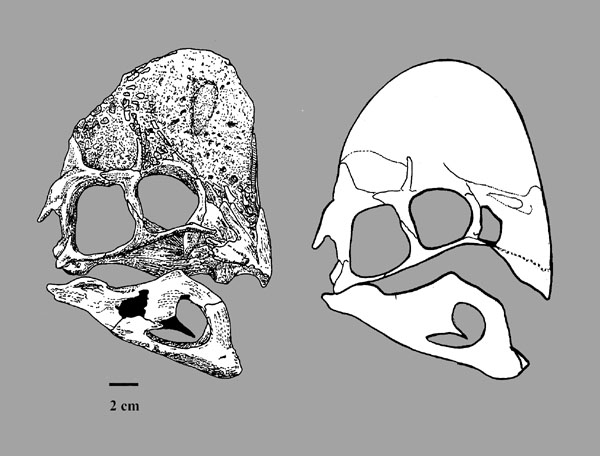
One group of primitive dromaeosaurid only recently recognized are the Halszkaraptorinae. Best known from little Halszkaraptor of the Late Cretaceous of Asia, these are short-armed dromaeosaurids with slender jaws with very tiny teeth: most definitely NOT rapacious predators of large prey! Even more bizarre, their bones are quite solid, suggesting that they may have been semi-aquatic. It has been suggested that they were like modern mergansers (fish-eating ducks) in terms of their ecology. Other Late Cretaceous Mongolian halskaraptorines include fragmentary Mahakala and Hulsanpes and nearly-completely known Natovenator. Teeth very much like Halskaraptor's are found in many parts of the world from the Late Jurassic onward, so it may be that these "pseudogeese" were actually a widespread successful group of dromaeosaurids.
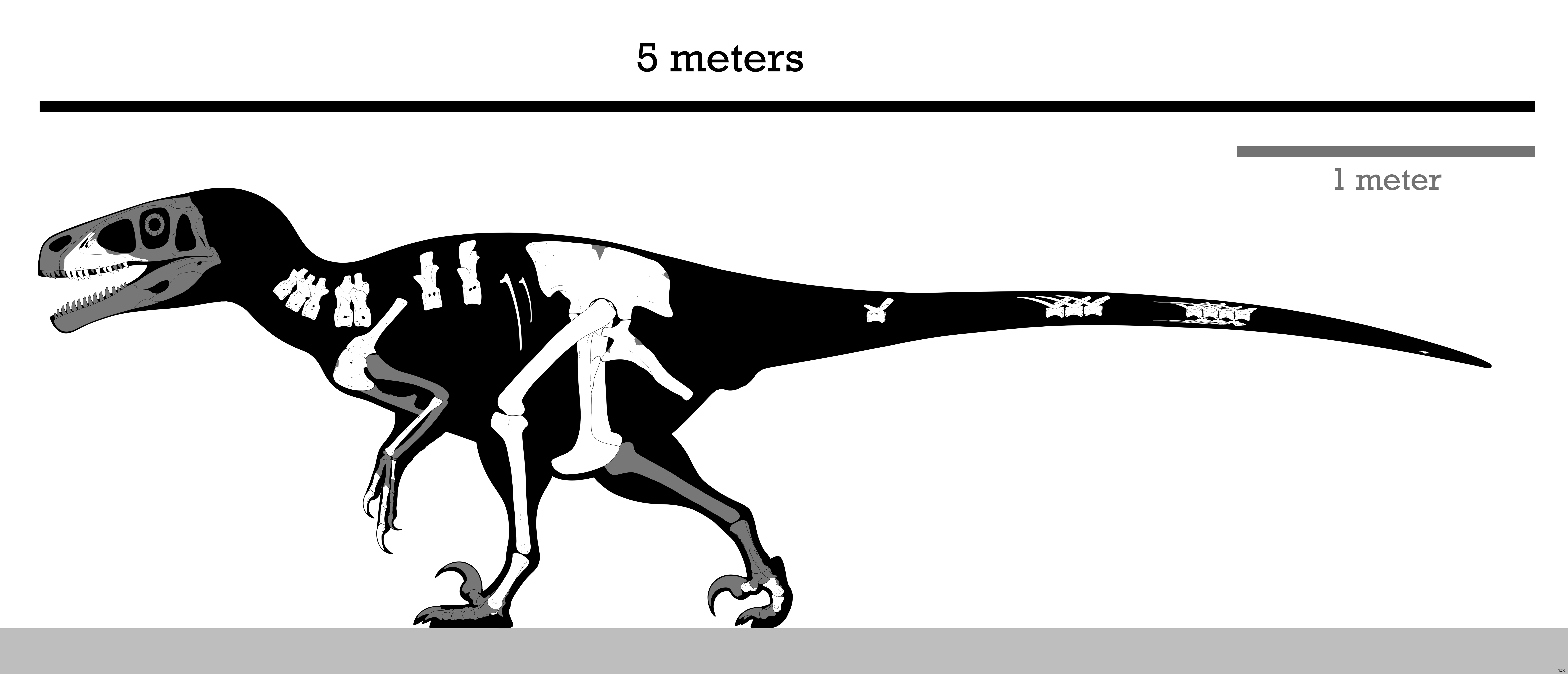
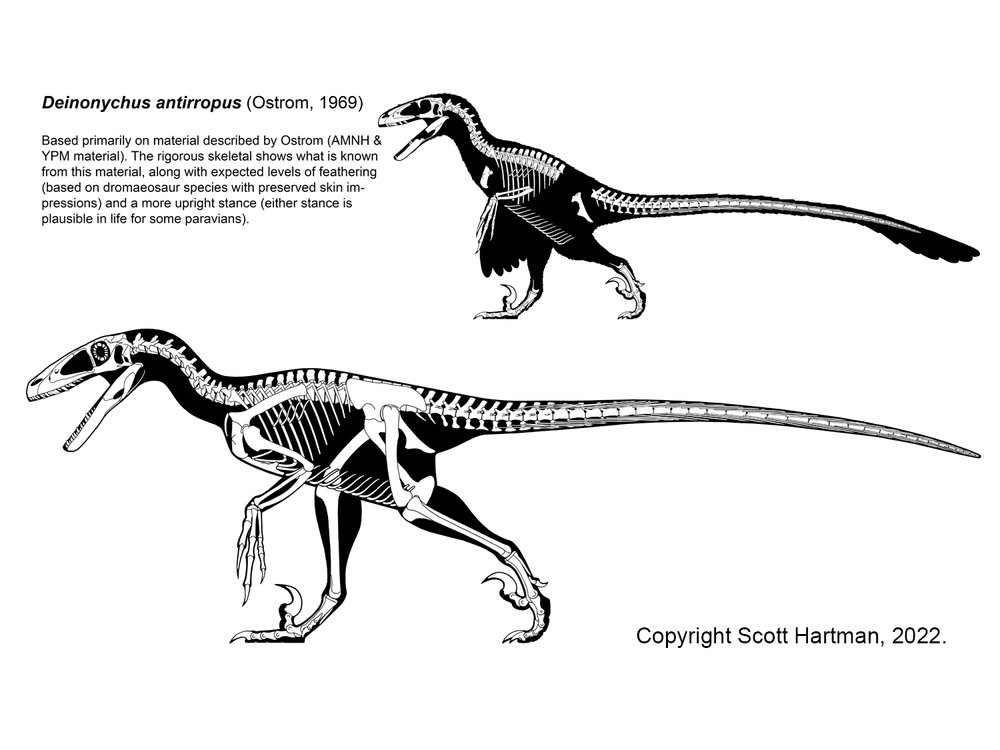
The more derived dromaeosaurids form a group called Eudromaeosauria. These were coyote-to-grizzly bear sized. Eudromaeosaurs include a major clade: robust Dromaeosaurinae. Dromaeosaurinae includes coyote-sized Dromaeosaurus of Late Cretaceous western North America; lion-sized Achillobator of mid-Late Cretaceous Asia; and grizzly-sized western North American Utahraptor of the Early Cretaceous and Dakotaraptor of the latest Late Cretaceous. At present Utahraptor and Dakotaraptor seem to be the largest known eumaniraptorans. The relationships among the remaining eudromaeosaurs are very uncertain. Traditionally the rest of them were grouped together as a clade Velociraptorinae; alternatively, some of the North American forms fall out as branches outside Dromaeosaurinae + Velociraptorinae. In general the non-dromaeosaurine eudromaeosaurs are more gracile than dromaeosaurines. Well-studied examples of include coyote-sized Velociraptor of the Late Cretaceous deserts of Mongolia; similar sized Saurornitholestes of Late Cretaceous North America; wolf-sized Deinonychus of the Early Cretaceous of western North America; and jackal-sized Acheroraptor of latest Cretaceous western North America.
Dromaeosaurid caudals were more tightly interlocked than in typical theropods, and in microraptorines and eudromaeosaurs extensions from the chevrons and neural arches grew extremely long. The tail was thus an extreme dynamic stabilizer.
Some basal microraptorines had elongate metatarsi (indeed, in some microraptorines there is a primitive form of the arctometatarsus), but eudromaeosaurs had relatively short stout metatarsi and tibiae. This suggests that they had sacrificed speed, perhaps for agility (better able to turn quickly while pursuing prey or escaping predators, especially with the help of the stiffened tail). Despite certain books and movies to the contrary, the dromaeosaurids show no signs of being speed specialists.
While the small primitive forms may have eaten small prey (skewering it with the sickle claw in the manner of modern secretary birds, perhaps), the larger forms were predators of dinosaurs. The "Fighting Dinosaurs" specimen of Velociraptor shows it in combat with Protoceratops, the hands used to grasp the head of the herbivore while the sickle claw was ripping into the throat: very similar to the attacks used by large cats. Some (controversial) evidence suggest that Deinonychus may have attacked the much larger iguanodontian Tenontosaurus in groups (packs or mobs).
Note that the sickle claw may have also been used to climb: either up trees, or up the sides of victims! Additionally, they may have been used to pin smaller prey down on the ground while the jaws and hand claws were used to kill it: the raptor prey restraint (RPR) model.
As discussed in a future lecture, at least some of the smaller basal forms may have had limited flight capability: indeed, they show somewhat better flight adaptations than early avialians such as Archaeopteryx and the anchiornithids! The majority of dromaeosaurids, however, probably lived their lives on the ground (although hiding/sleeping in the trees may have been possible, especially for juveniles).
In the Early Cretaceous dromaeosaurids were major mid-sized predators, and in the deserts of Late Cretaceous Asia they were among the largest carnivorous dinosaurs present.
[Hey, want to make eumaniraptoran systematics MORE confusing! Some studies suggest that microraptorines, unenlagiines, halszkaraptorines, and/or troodontids might be more closely related to Aves than to the eudromaeosaurs; this would make microraptorines, unenlagiines, halszkaraptorines, and/or troodontids members of Avialae!]
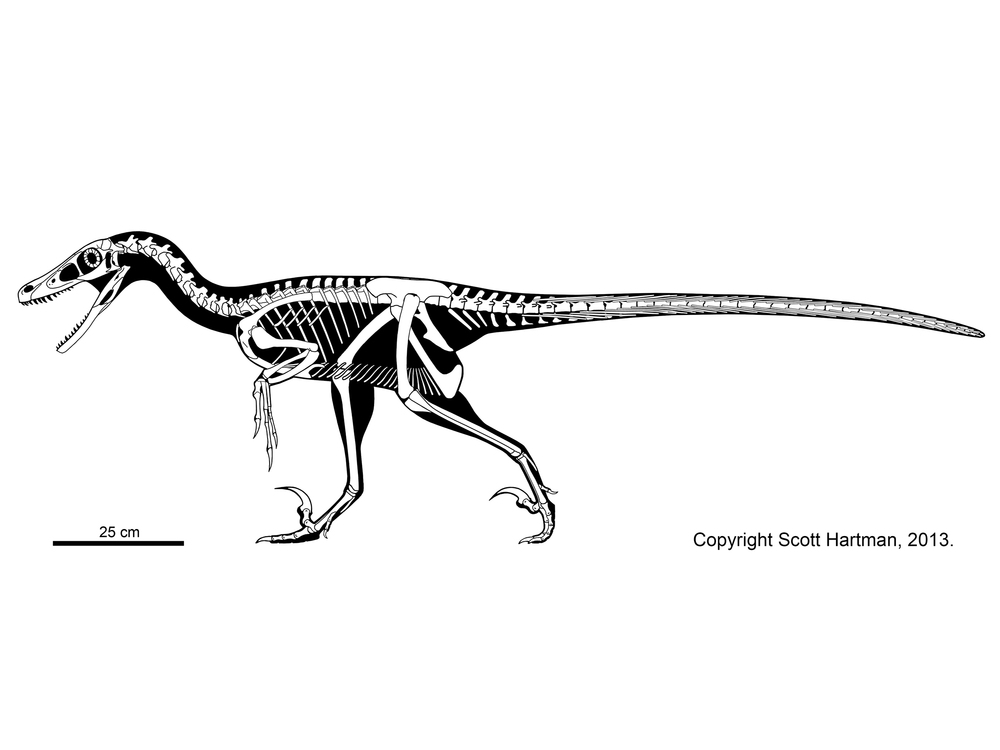
TROODONTIDAE
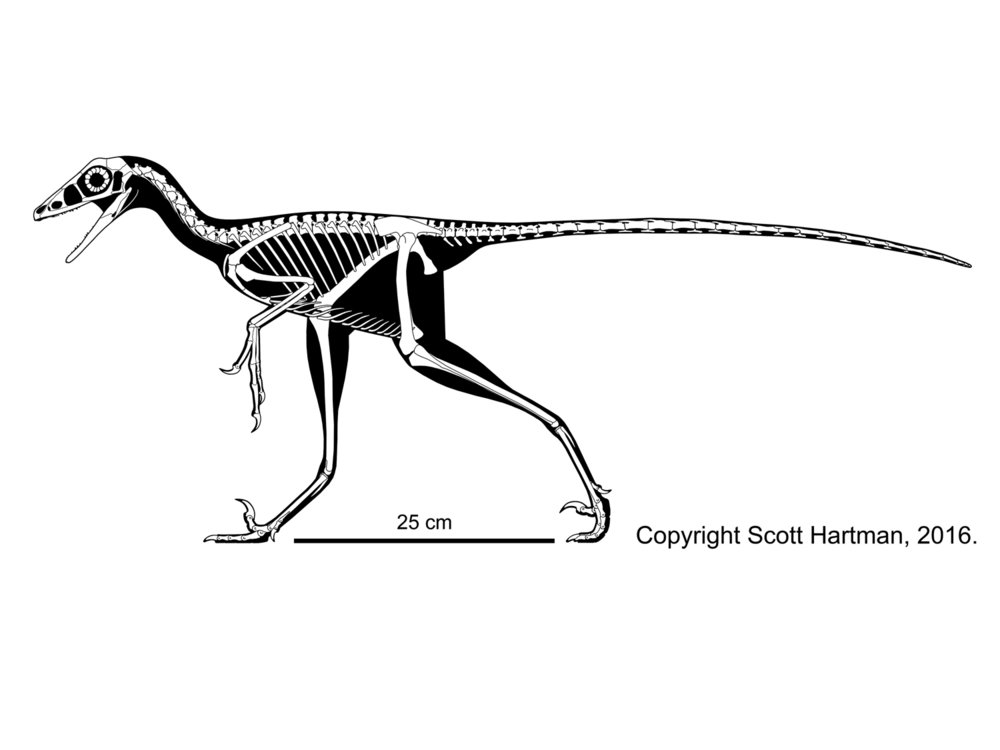
Troodontids were pigeon-to-deer sized theropods. The oldest known troodontid is the recently-described Hesperornithoides of the Late Jurassic of western North America. Somewhat younger is Jinfengopteryx of China: described originally as a bird, but now recognized as a troodontid. It is from the early Early Cretaceous (although for some time it was thought that the formation it is from was as old as the Jurassic). Its gut contents include small ovoid objects, that might be seeds. Like Archaeopteryx, Jinfengopteryx had long tail feathers all along its tail. (Various other troodontids, such as tiny Early Cretaceous Liaoningvenator, Late Cretaceous Almas, and latest Cretaceous Tamarro, form a clade Jinfengopteryginae, with Jinfengopteryx.) The last troodontids are from the very end of the Cretaceous. As discussed above, some studies once placed anchiornithids as the basalmost troodontids.
Troodontids differ from other eumaniraptorans by an increase in the number of teeth, reduced arm length, and increased distal hindlimb length. Tiny Sinovenator of the Early Cretaceous of China has a backwards-pointing pubis and lacks an arctometatarsus; most later troodontids show a reversal to a forward-pointing pubis and have an arctometatarsus. (But the newly recognized Latenivenatrix has a backwards-pointing pubis, just to be confusing...)
Troodontids have relatively lightly built snouts, suggesting that they did not tackle very large prey (particularly as their forelimbs were fairly short and lightly-built as well). The largest and most derived troodontids are not particularly large (turkey-to-deer sized), and had forward-facing eyes and leaf-shaped teeth with large denticles on the back. They may have had some plants in their diet, but tooth-marked bones show that they definitely ate meat (recent isotopic studies support the hypothesis that derived troodontids were omnivores). (NOTE: if troodontids were omnivores, this would make Dromaeosauridae the only primarily carnivorous group of maniraptoriform. If true, this would almost certain be an evolutionary reversal, and indicate that Velociraptor and Deinonychus were descendants of omnivores or even herbivores!)
The more completely known troodontids include Mei and Sinornithoides of the Early Cretaceous of Asia (and both known from specimens buried in sleeping position!); Saurornithoides, Xixiasaurus, and Zanabazar of the Late Cretaceous of Asia; and Talos and "Troodon" of the Late Cretaceous of western North America. (Okay, it is now getting complicated. The original specimens of Troodon were isolated teeth, and we can't be certain which skeleton-based taxon it belongs to. Some split the classic "Troodon" specimens into Stenonychosaurus and Latenivenatrix (the latter is largest known member of the Troodontidae); others consider these both Stenonychosaurus, and still more place them all in Troodon. A latest Cretaceous form known as Pectinodon exists, but we don't presently know what its skeleton looked like.
The large eyes of troodontids suggests to some that they may have been nocturnal. Possibly related to that, troodontids are by far the most common dinosaur in polar Late Cretaceous North American fossil sites (where light levels would have been very low during the winter), while they were no more common than dromaeosaurids in the rest of their range. "Troodon" (in the broad sense: the collection of Late Cretaceous western North American troodontids: Talos, Latenivenatrix, Stenonychosaurus, Latinevenatrix, and company) is particularly notable by having among the largest brain/body size of the non-pygostylian dinosaurs, and thus likely the "smartest" of the traditional dinosaurs.
As mentioned above, at least some phylogenetic analyses place Troodontidae as closer to extant birds than to Dromaeosauridae; in these studies, Troodontidae would be part of Avialae as traditionally defined rather than Deinonychosauria.
EVOLUTIONARY PATTERNS IN MANIRAPTORANS
Feeding adaptation transformations:
Terrestrial locomotory adaptations:
Miniaturization (and a little Gigantism):

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}