


Key Points:
•Among the primitive ornithischians are big-handed, deep-skulled Heterodontosauridae and slender Eocursor; the rest form the clade Genasauria
•Thyreophorans represent the armored dinosaurs, and are a clade of (predominantly) quadrupedal ornithischians.
•There are characterized by the presence of osteoderms (armor plates) in their skin. Different clades of thyreophorans express these osteoderms in different patterns.
•Beyond a few basal taxa, thyreophorans are divided into the plated Stegosauria and the tank-like Ankylosauria.
•Armor in thyreophorans seem to have functions beyond simple defense: they served as display structures and (in the case of the stegosaurs and the club-tailed ankylosaurine ankylosaurs) as active weapons.
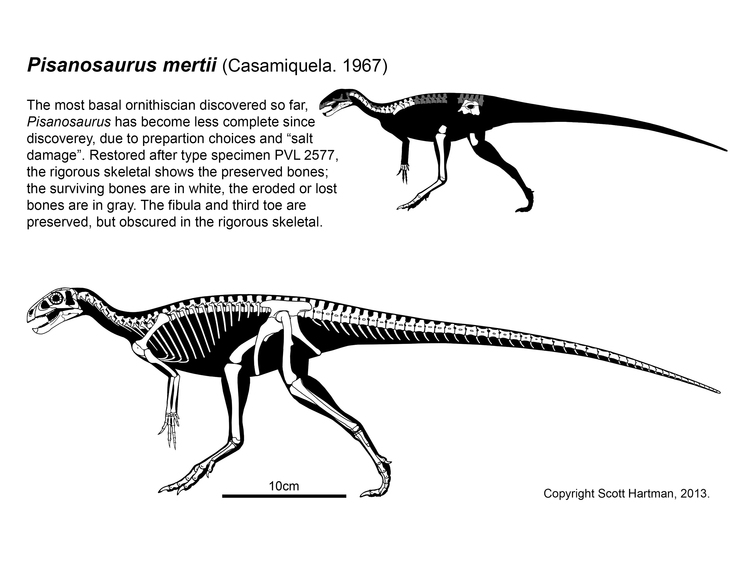
As we saw in the last lecture, the only likely Triassic non-"silesaur" ornithischian is Pisanosaurus of the early Late Triassic Argentine Ischigualasto Formation. The fossil is incomplete, so many aspects of its anatomy are uncertain.
As previously discussed, Pisanosaurus plus Prionodontia is characterized by the following traits:
Prionodontia itself is characterized by:
Primitive Ornithischian Groups: Heterodontosaurids & Eocursor:
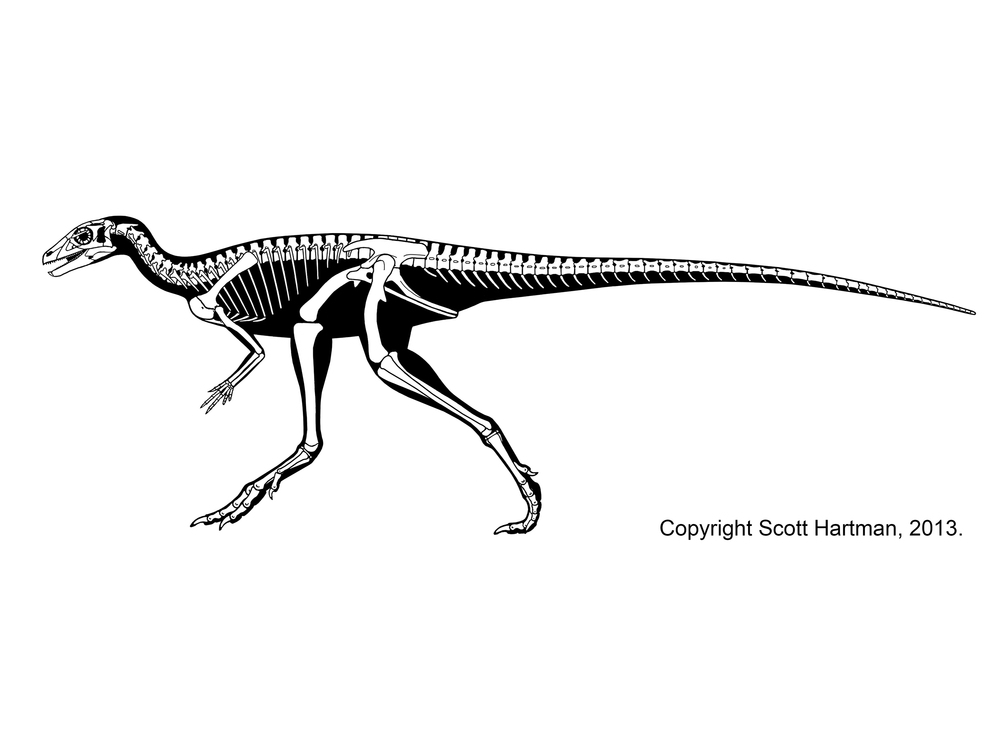
Heterodontosauridae was a primitive group of ornithischians. Although one fragmentary specimen was thought to be from the Late Triassic, redating shows it was younger; the oldest heterodontosaurids are known from Early Jurassic, and persist into the mid-Early Cretaceous. They had skulls which are relatively deep and powerfully built, indicating that they ate fairly tough food. Advanced heterodontosaurids also had premaxillary tooth row that were ventral to the maxillary tooth row and jaw joints that were ventral to the dentary tooth row: the result were jaws that brought the teeth together all at once (like a nutcracker), and not slicing (as in scissors, or as in most dinosaurs). These latter jaw adaptations evolved convergently in Ornithopoda, and so for a long time Heterodontosauridae was considered a clade within Ornithopoda. However, primitive heterodontosaurs lack these convergent adaptations.
Most heterodontosaurids are quite small. Some are only about 1-1.5 m long, and Fruitadens of the Late Jurassic of western North America may have been no more than 80 cm long as an adult (most of which length is tail) and Manidens of the Middle Jurassic of Argentina only 65-75 cm; that makes these the smallest known ornithischians.
Interestingly, the early Late Jurassic Chinese heterodontosaurid Tianyulong had a fuzzy body covering over at least part of its body! If this is found to be homologous to the protofeathers of tetanurine theropod saurischians it would suggest that the concestor of all dinosaurs was fuzzy, and that dinosaurs were thus fuzzy ancestrally! At present, however, there is enough uncertainty to make the homology between Tianyulong's fuzz and tetanurine protofeathers suspicious. (But do not be terribly surprised if in the future we discover that most dinosaurs were fuzzy to some degree or another! All we need is a fuzzy primitive sauropodomorph, and it is basically a done deal!) (By the way, the initial reports placed Tianyulong in the Early Cretaceous, but the formation in which it was found has been redated to the earliest part of the Late Jurassic, around 160 Ma.) (A few years ago a radical new hypothesis for the position of Heterodontosauridae was proposed: that "heterodontosaurids" were a paraphyletic grade of Jurassic and Early Cretaceous pachycephalosaurs! However, the latest most comprehensive studies have not supported this hypothesis.)
The similarly aged neornithischian Kulindadromeus of Siberia also shows simple filimentous fuzz, as well as scales, plates, and additional bizarre tufted plates, showing that primitive ornithischians had a wide variety of integumental features.
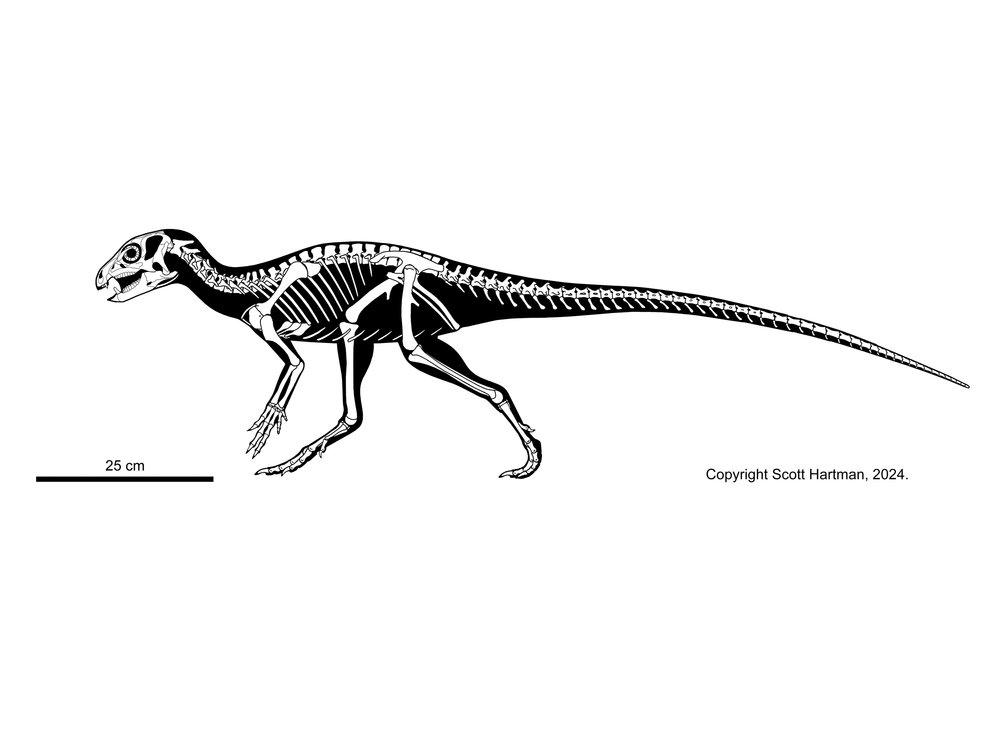
Once thought to be from the Late Triassic, Eocursor of South Africa is from the earliest Early Jurassic.
Ornithischians more derived than Heterodontosauridae, Eocursor, and Laquintasaura had greatly reduced hands, losing most of their grasping ability. This suggests a switch to jaws-only while obtaining food.
The remaining ornithischians (Genasauria, the "cheeked reptiles") include the armored Thyreophora and the highly diverse Neornithischia (especially beaked Ornithopoda and ridge-headed Marginocephalia).
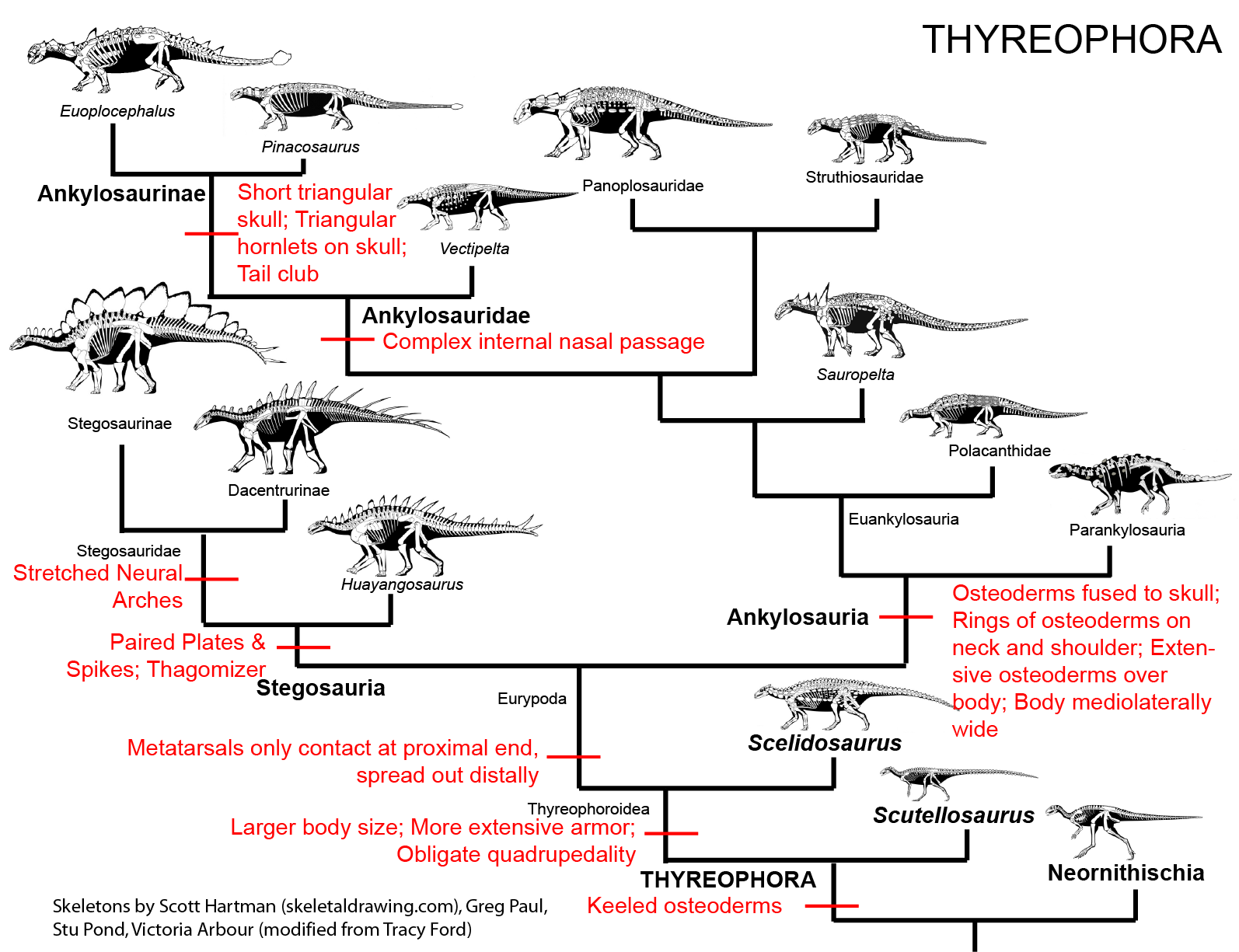
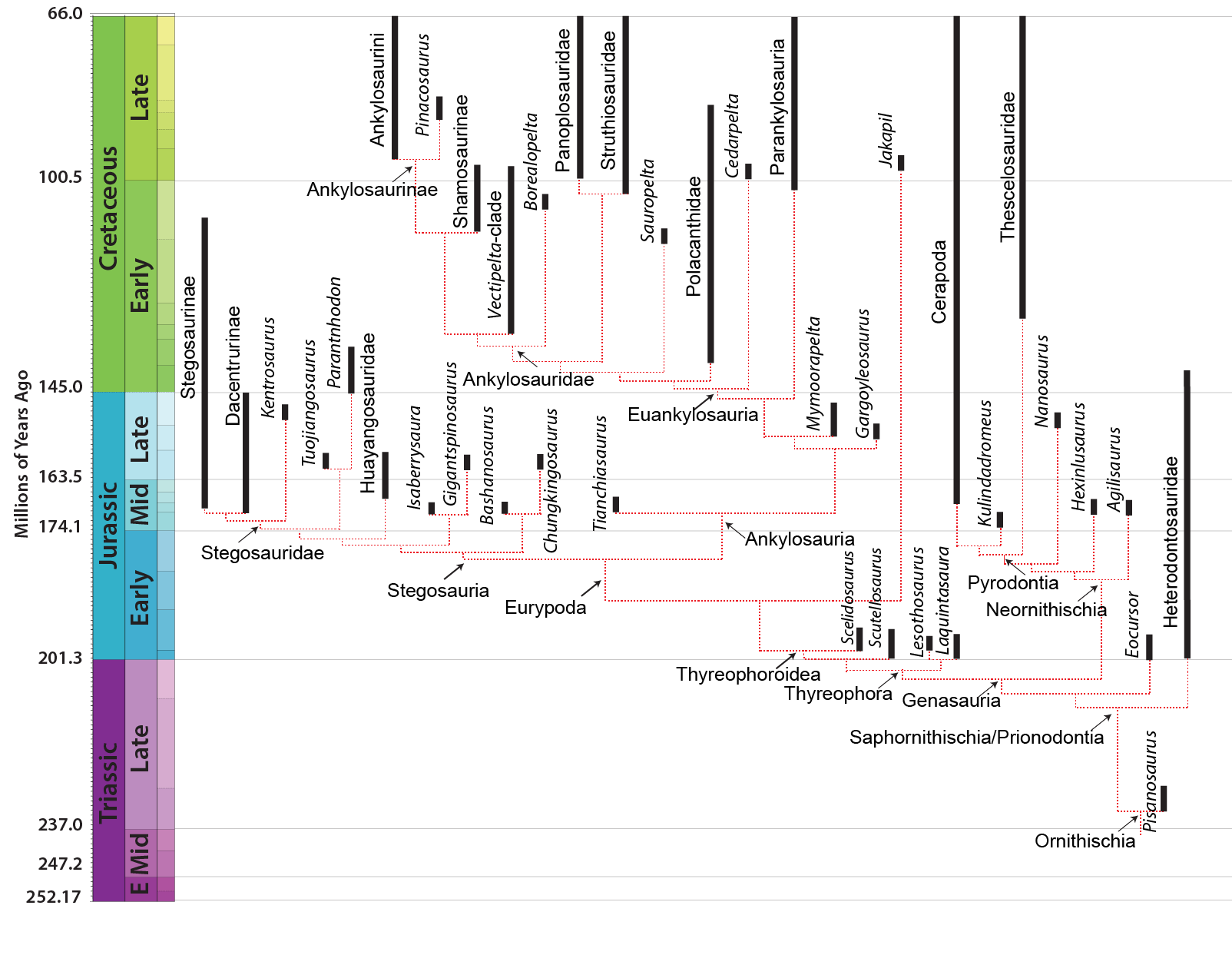
MAJOR GROUPS OF THYREOPHORANS
Thyreophora are united by various skeletal attributes, the most obvious of which is:
The derived thyreophorans are the plated Stegosauria (shingled lizards) and the heavily armored Ankylosauria (fused lizards). Both clades are present by the Middle Jurassic. There are several Early Jurassic taxa which fall outside either of the two advanced clades.
BASAL THYREOPHORANS
Lesothosaurus of the Early Jurassic of southern Africa and similarly-aged Laquintasaura appear to be the oldest and earliest branching thryeopohiorans. Laquintasaura is the oldest and most primitive ornithischian known from group assemblages, indicating that at least some of these lived in groups during life. These are united with the later thyreophorans on various subtled cranial and skeletal traits that are outside the scope of this course. Importantly, these are the only known unarmored thyreophorans. (But we would expect the oldest ones to have been unarmored.) To be fair, other analyses find one or both to be the oldest and most primitive neornithischian(s) or in their traditional positions as non-genasaurian ornithischians.
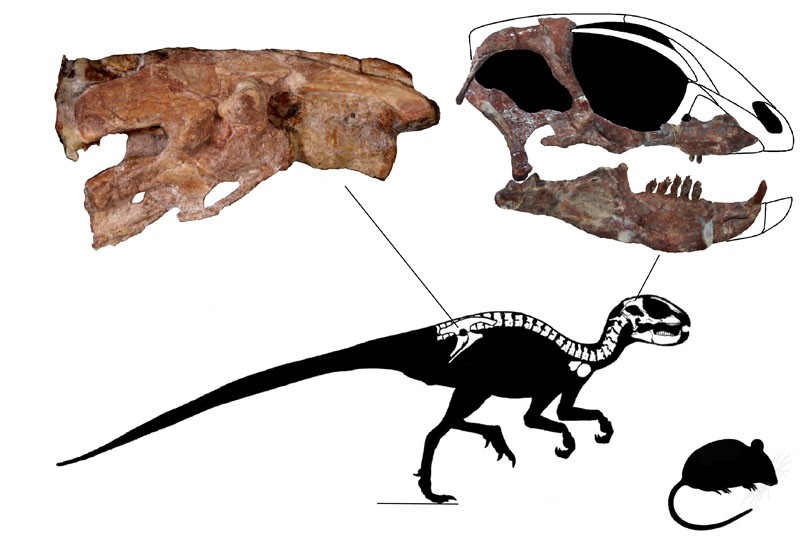
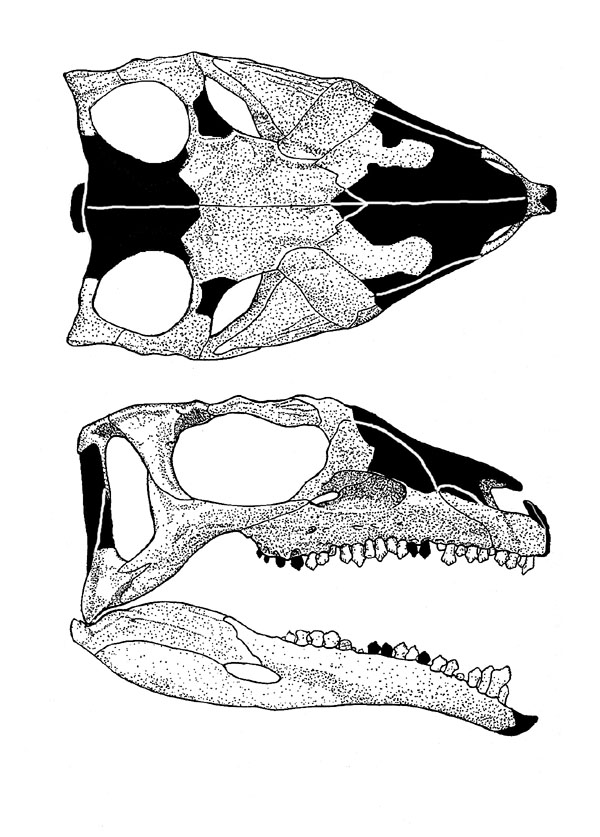
The oldest and most primitive definite (i.e., armored) thyreophoran is Scutellosaurus of the Early Jurassic of western North America. It was a 1.5 m long biped (possibly facultative biped) not very dissimilar to other primitive ornithischians like Lesothosaurus or Hexinlusaurus: small herbivores with small hands. The primary distinction of Scutellosaurus is the presence of a great many small osteoderms over the body. These would protect against small-bodied predators, but might not help against the new larger theropods that had begun to appear in the Early Jurassic.
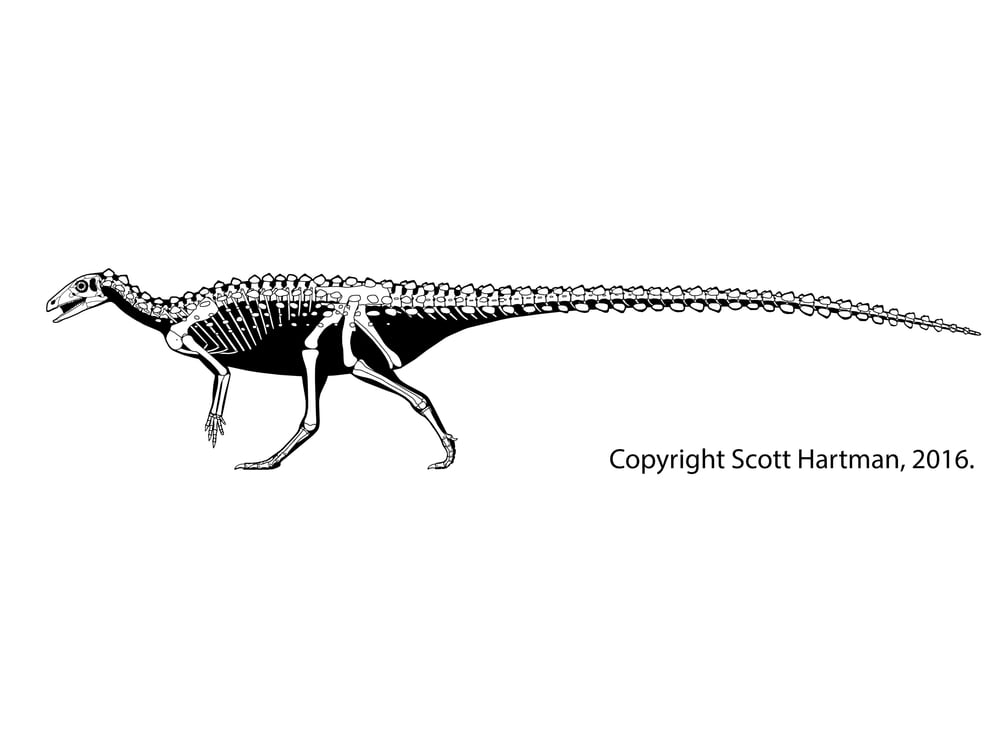
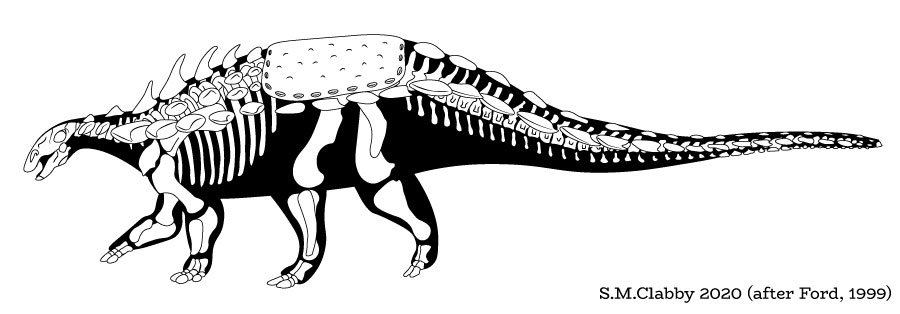
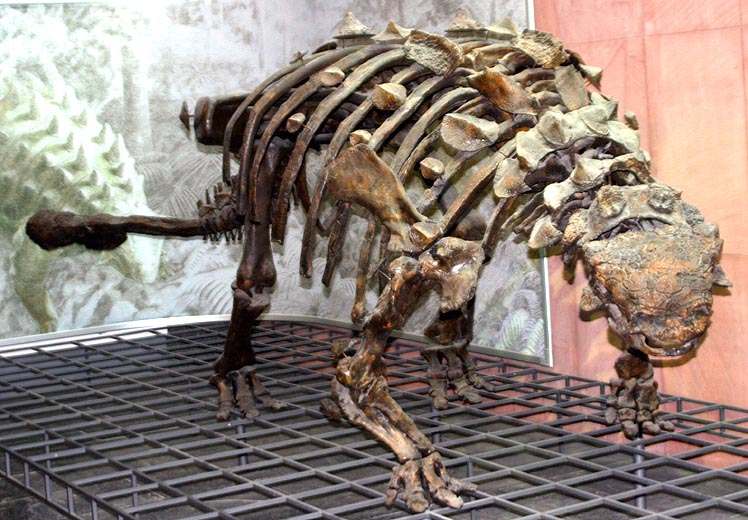
In response, thyreophorans evolved larger size and heavier armor, as seen in Emausaurus and Scelidosaurus (new specimen shown here), both of Europe, and comparable-aged Yuxisaurus of China. The larger body size (3-4 m long) and proportionately larger osteoderms may have been more effective defense against attacking predators, but forced them onto all fours (at least for Scelidosaurus): in other words, they were obligate quadrupeds.
(Note that the hypothesis shown here is that Scutellosaurus, Emausaurus, and Scelidosaurus were progressive closer to the Stegosauria-Ankylosauria clade (Eurypoda). However, some paleontologists have considered Emausaurus to be a primitive stegosaur, and others that Scelidosaurus was the oldest and most primitive ankylosaur. However, eurypods share a number of transformation not found in these Early Jurassic taxa: these include:
A possible fly in the ointment of this scenario is recently (2022) described Jakapil of the early Late Cretaceous of Argentina. Known only from a single partial skeleton, this seems to be a late survivor of the non-eurypodan thyreophorans, even closer to Eurypoda than Scelidosaurus. However, the remains of the forelimb and hindlimb indicate it was almost certainly a biped! Obviously we need more of the skeleton to make certain, but Jakapil is a good reminder that Evolution is not a planned directional phenomena, and that some lineages will buck the trend that the rest of their clade are following if that is the most successful set of adaptations at that place and time. Also, there is the definite non-zero chance that Jakapil belongs to some entirely non-thyreophoran group that convergently evolved osteoderms. Indeed, its mandible is very much like that of a neornithischian, so perhaps it is the first discovery of a previously-unknown clade of Gondwanan marginocephalian!
STEGOSAURIA
From a Scelidosaurus-like ancestor, the stegosaurs evolved armor that was less covering and more concentrated. While they had some small osteoderms in their skin (particularly around the neck and the hips), most of their armor was specialized as:
In general, the stegosaur armament suggests active defense: the dinosaur probably turned in response to attacking predators, trying to keep the tail towards the theropod so that it could use its thagomizer. Damaged thagomizer spikes and theropod bones with thagomizer-generated puncture wounds confirms their use in defense.
Early stegosaurs were only about 2.5-3 m long, but the most derived forms ranged up to 9 m or more. Their narrow snouts suggests that they were rather picky eaters (that is, instead of munching a lot of plants at once, they were selective as to which ones they chomped.) Biomechanical analysis shows that their bite was stronger than many herbivorous saurischians, but still weaker than many specialized ornithischians. Although they were obligate quadrupeds in terms of locomotion, they may have been able to rear on their hind legs in order to feed higher in trees.
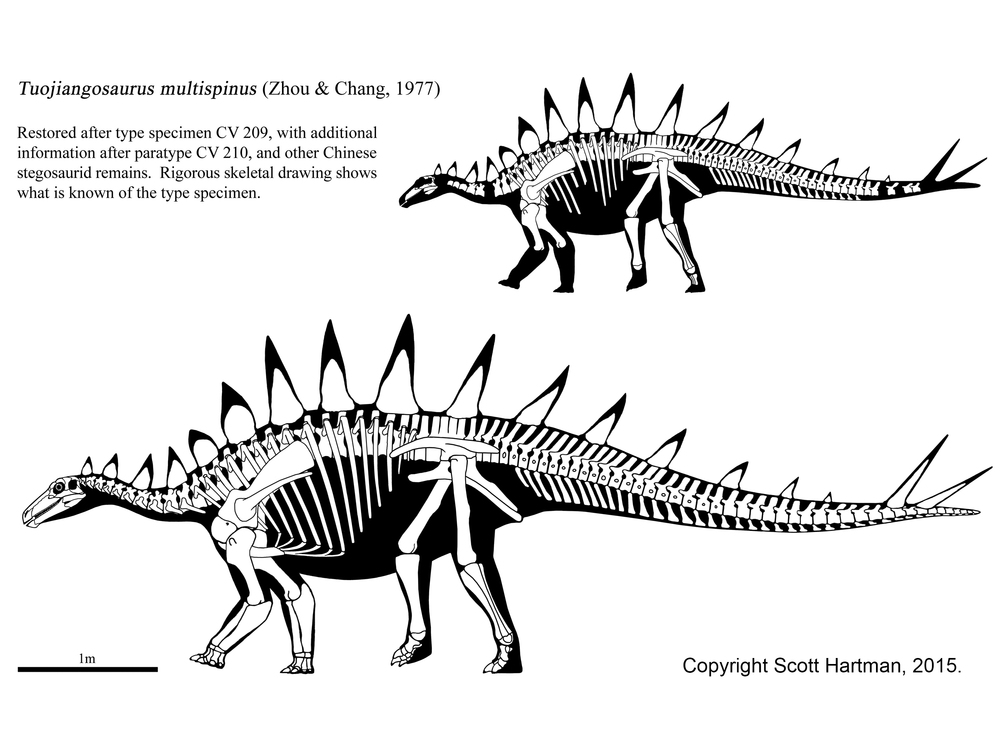
Some tantalizing footprints suggest possible Early Jurassic stegosaurs, but these may be from a more basal Scelidosaurus-like thyreophoran instead. Primitive stegosaurs include Middle Jurassic Isaberrysaura of Argentina (first reported as being an ornithopod, and being Early Jurassic) and Bashanosaurus and Huayangosaurus of China; and Late Jurassic Chungkingosaurus, Gigantspinosaurus and Tuojiangosaurus (all three from China), western North American , Alcovasaurus [figures A, B, E & F], and Early Jurassic Paranthdon of South Africa. More derived stegosaurs form the clade Stegosauridae, and include Middle Jurassic Adratiklit of Morocco, long-necked Late Jurassic Dacentrurus (which may include the smaller stegosaur typically considered its own genus, Miragaia) of Europe, Kentrosaurus of eastern Africa, and the Stegosaurinae.
Stegosaurids had dorsoventrally stretched neural arches and disproportionately short forelimbs, giving them an odd profile.
The most derived stegosaurids (Stegosaurinae) lacked shoulder spines (also missing in Tuojiangosaurus), had only plates rather than spikes along the back (again, shared with Tuojiangosaurus), and had alternating rather than parallel plates. This advanced group includes Middle Jurassic Loricatosaurus of Europe, Late Jurassic Jiangjunosaurus of China, the two Late Jurassic western North American genera (Hesperosaurus and famous Stegosaurus [also known from Portugal]), and Early Cretaceous Wuerhosaurus and Yanbeilong of China, the last of the [distinctive] stegosaurs. (Note: some paleontologists consider Wuerhosaurus, and Hesperosaurus to belong within the genus Stegosaurus.)
Stegosaurs are relatively common in Middle and Late Jurassic formations (especially so in China), are present but rare in some Early Cretaceous Asian, African, and European assemblages, and vanish before the end of the Early Cretaceous (at present, Yanbeilong of China and Mongolostegus of Mongolia are the youngest known). Claims of later stegosaurs have so far turned out to be either mis-dated or misidentified.
ANKYLOSAURIA
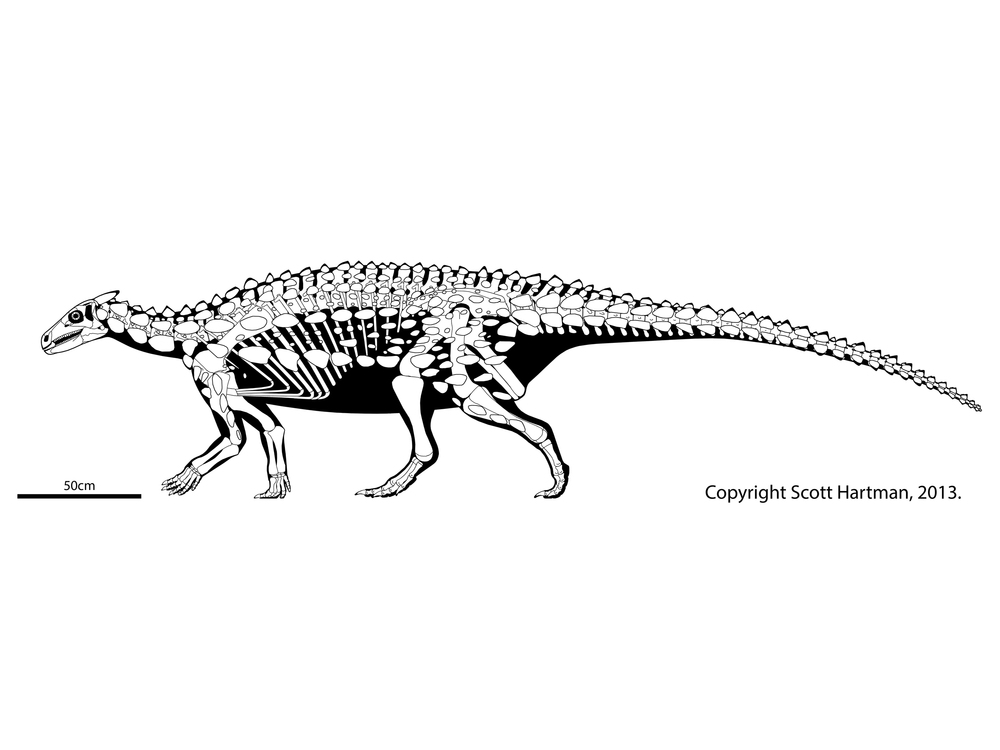
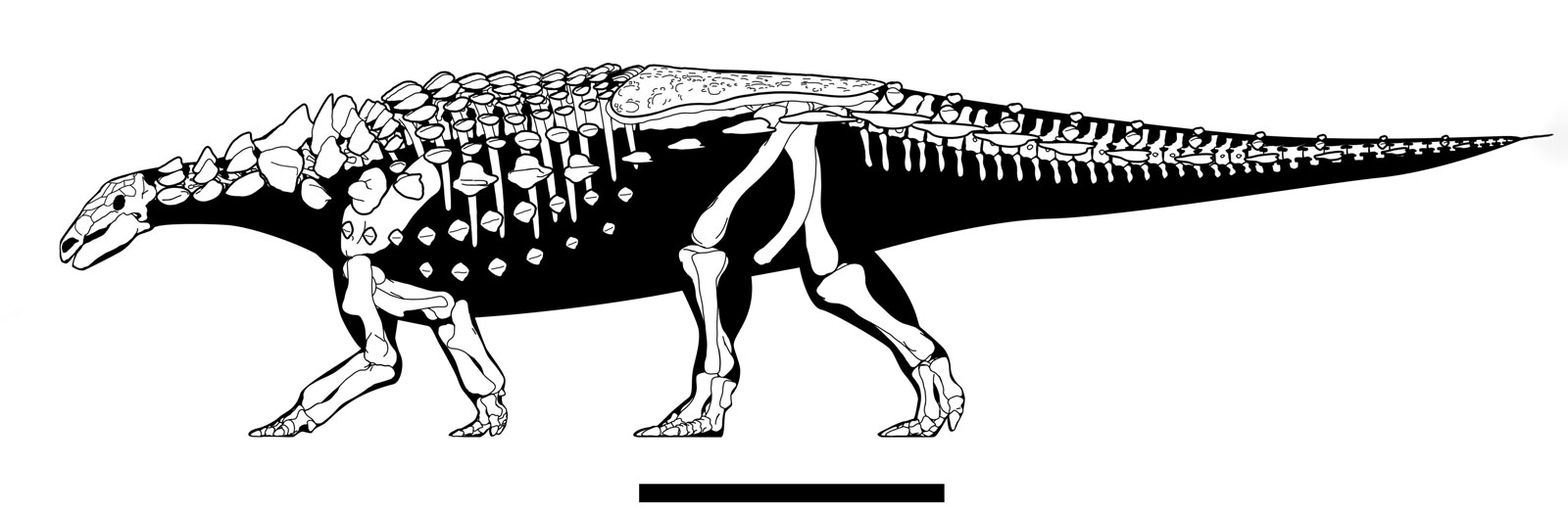
While the stegosaurs evolved active defense, the ankylosaurs (at least at first) seem to have been selected for passive defense: the ability to stay put and absorb damage from an attack. They were even more extensively armored than Scelidosaurus, and were characterized by:
Additionally, in ankylosaurs the predentary bone is reduced, and the jaws arranged so that they would have more extreme rotation (and also be pulled back further) when chewing then in most ornithischians. This evolved convergently (and to a far more extreme form) in the advanced ornithopods. This motion allowed ankylosaurs to more effectively chew up their food.
The oldest ankylosaurs are from the Middle Jurassic: Tianchiasaurus of China, Sarcolestes of Britain, and newly discovered Spicomellus of Morocco. None of these are known from anything approaching a complete skeleton, and it is not inconceivable that one or more of these are actually more basal thyreophorans (in the Scelidosaurus-part of the tree). Alternatively, as mentioned previously, some analyses place Early Jurassic Scelidosaurus as the basalmost ankylosaur rather than a non-eurypod thyreophoran.
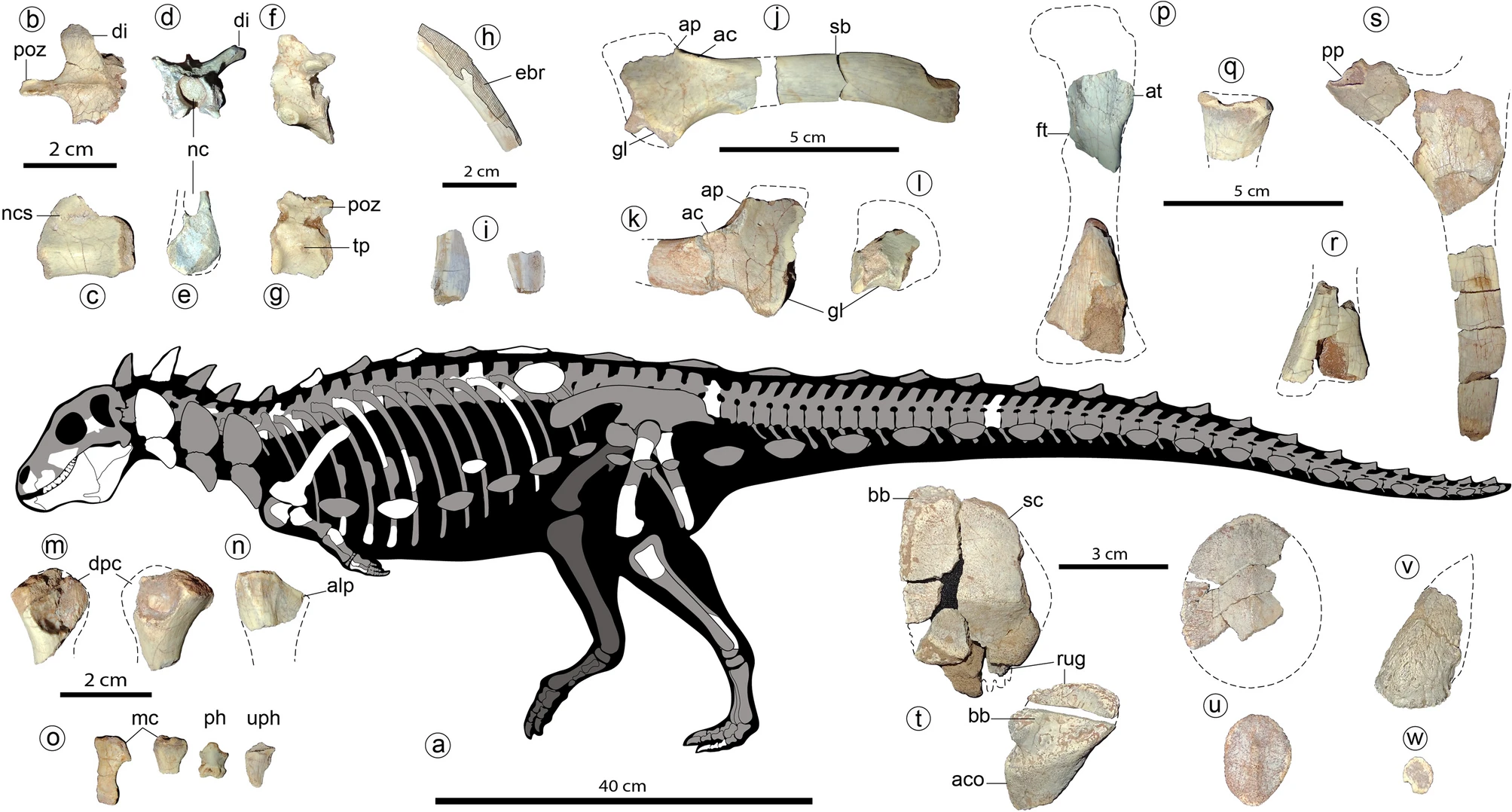
Although still only incompletely known, newly described specimens of Spicomellus show it to be EXCEPTIONALLY long spined, with many of the shoulder and dorsal spines fused directly to ribs! Furthermore, it has a spiked thagomizer-style tail weapon, differing from later ankylosaurs. Its phylogenetic position remains uncertain, but its aberrant anatomy and extremely early age suggests it was a very basal branch within Ankylosauria.
Ankylosaur systematics is at present quite complicated. Traditionally Ankylosauria was divided into the club-tailed Ankylosauridae and the club-less Nodosauridae. Almost all recent studies, however, show that the "nodosaurids" are a paraphyletic grade comprised of several subgroups. The problem is that the different analyses don't agree what genera belong to which subgroup, nor do they agree on the arrangement of these subgroups relative to each other or to Ankylosauridae. An attempt at consensus is shown here, but this particular arrangement doesn't confirm to any one particular study and will almost assuredly change with future studies.
Late Jurassic North American Mymoorapelta and Gargoyleosaurus have uncertain positions: possibly basal branches of Ankylosauria, or basal ankylosaurids, or basal polacanthids, or some combination thereof.
In late 2021, there was the announcement of a relatively complete skeleton of a Late Cretaceous ankylosaur from southern Chile, given the name Stegouros. This taxon showed traits which united it with Early Cretaceous Kunbarasaurus of Australia and poorly-known Late Cretaceous Antarctopelta of Antarctica and Patagopelta of Argentina formed a clade Parankylosauria, found to be the sister group to Euankylosauria (the remaining "nodosaurids" plus Ankylosauridae). Stegouros at least has an interesting slashing tail weapon: an expanded series of laterally-oriented blade-like osteoderms.
Polacanthidae includes Early Cretaceous Gastonia of North America and Polacanthus of Europe, and possibly Hylaeosaurus (the first discovered thyreophoran). A number of other Early and Late Cretaceous ankylosaurs appear to be polacanthids, including gigantic early Late Cretaceous Peloroplites and smaller mid-Late Cretaceous Niobrarasaurus, both of North America. They tend to have extremely large blade-like osteoderms in their shoulder regions and a shield over the sacral region formed by a series of large osteoderms surrounded by many smaller ones.
The North American Early Cretaceous Sauropelta and poorly known Tatankacephalus show some similarities to polacanthids, but also to the other "nodosaurid" clades.
The Struthiosauridae are a radiation of primarily European forms such as Europelta of the Early Cretaceous, Pawpawsaurus and Stegopelta of the Early-Late Cretaceous boundary (the only North American struthiosaurids), and Struthiosaurus and Hungarosaurus of the Late Cretaceous.
Struthiosaurids may form a clade with Panoplosauridae, a group of primarily North American Late Cretaceous forms. Edmontonia, Denversaurus, and Panoplosaurus are the best known examples. These have broader, smoother skulls than most "nodosaurs", and often have huge laterally-oriented spines along their sides in the shoulder region. Some of these are among the largest ankylosaurs. Early Late Cretaceous Nodosaurus itself may belong to this group (if so, then either "Panoplosauridae" or "Panopolosauridae + Struthiosauridae" would then properly be "Nodosauridae"!)
There remain, however, a number of "nodosaur"-grade ankylosaurids which are quite difficult to place. Early Late Cretaceous North American Animantarx and Cedarpelta do not have stable positions in these studies. Tiny Liaoningosaurus of Early Cretaceous China is known only by probable juveniles and only in lake deposits: some have speculated it was a semiaquatic fish eater!
ANKYLOSAURIDAE
The remaining ankylosaurs do seem to form the clade Ankylosauridae; however, that group is not limited to the club-tailed forms of the Late Cretaceous as once thought. Among the most primitive known member of Ankylosauridae is spectacularly-preserved Borealopelta. There is a newly discovered clade of shield-hipped ankylosaurids (in which the shield is formed of fused osteoderms of all the same size) which includes European Early Cretaceous Vectipelta and Early/Late Cretaceous Dongyangopelta and early Late Cretaceous Zhejiangosaurus of China.
The remaining ankylosaurids have distally-stiffened tails (although to be fair, complete tails are not recovered in most of the earlier forms.)
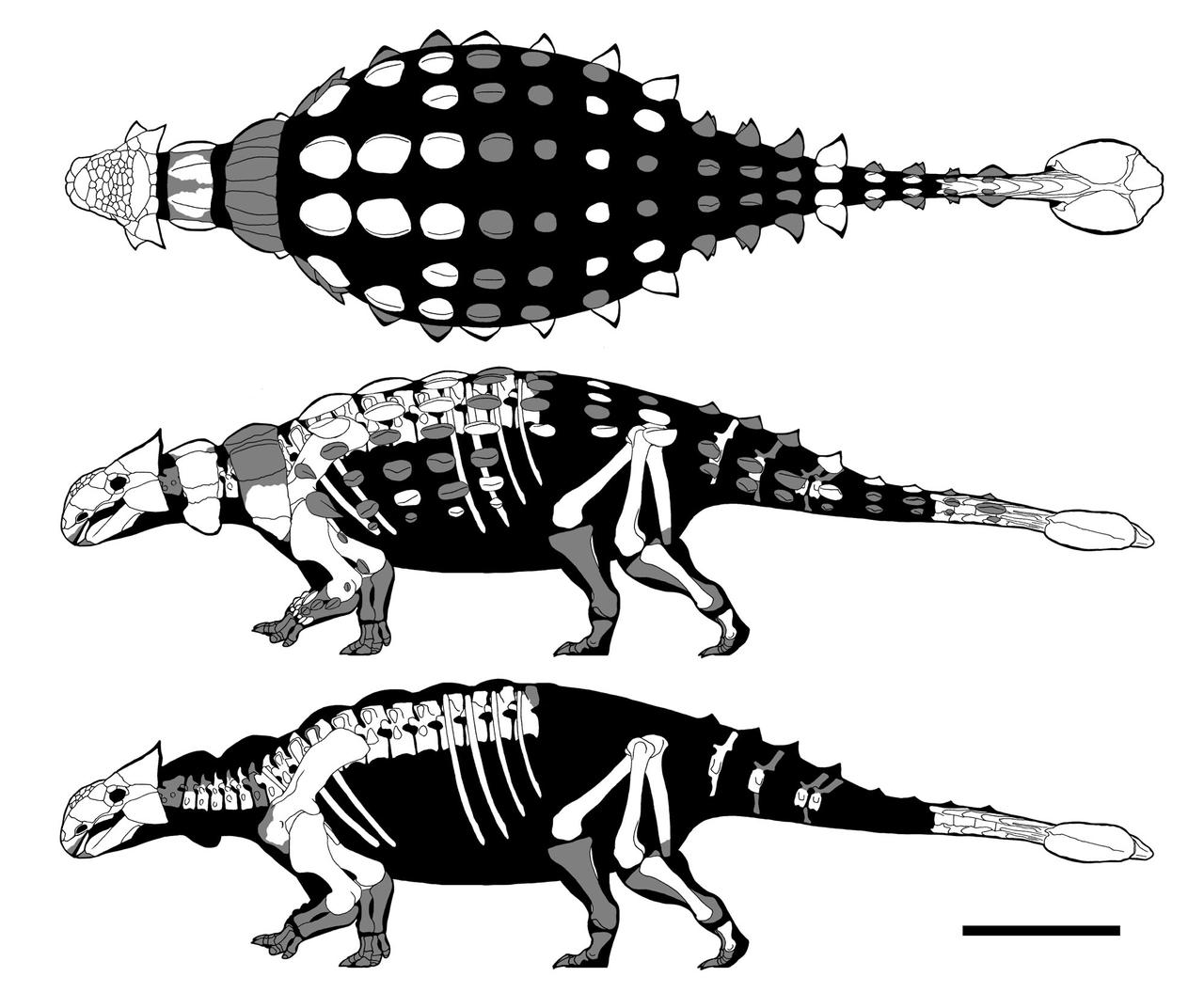
The best known part of Ankylosauridae is the more derived clade Ankylosaurinae there are extremely complex air chambers in the skull (convergently evolved with Gastonia)
Additionally, ankylosaurines are further transformed relative to other armored dinosaurs in having:
Ankylosaurids ranged from about 3 m to 8-10 m (the latter size includes Tarchia of Mongolia and gigantic Ankylosaurus of western North America). Pinacosaurus of Asia is one of the best known primitive ankylosaurines. Other Asian forms from the Late Cretaceous include Zaraapelta, Saichania, and Talarurus. Euoplocephalus, Dyoplosaurus, Akainacephalus, Zuul, Ziapelta, Scolosaurus and Anodontosaurus of the Late Cretaceous of western North America are the best studied.
Ankylosaurines are only known from the late Early Cretaceous of Asia and the Late Cretaceous of Asia and western North America at present; similar distributions are known for various other dinosaurs, as we shall see.
Ankylosaurids are more than their armor! Recent work has demonstrated that the bones which control the tongue and throat are incredibly well-developed in these dinosaurs: far more so than all other dinosaurs (outside of some birds.) They may have had long, powerful tongues: possibly for insect eating, possibly for grabbing plants, and possible for both and more. However, even more recent discoveries show that these bones also supported a powerful larynx (voice box): this suggests that ankylosaurids (and perhaps other dinosaurs) might have had strong vocal communications.
EVOLUTIONARY PATTERNS IN THYREOPHORA
Defense:
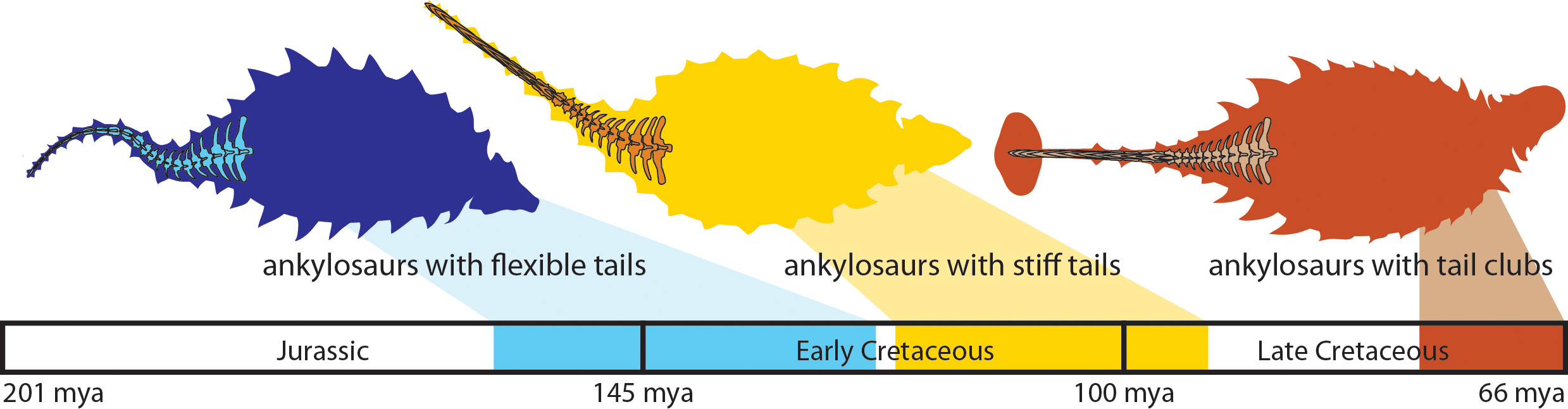
Probably the most conspicuous aspect of their evolution. Thyreophorans begin with a set of small scutes; develop larger scutes at
the expense of bipedalism (and speed); then split between specialists in active (Stegosauria) vs. passive (Ankylosauria) defenses. Within the ankylosaurs, though, Parankylosauria and Ankylosaurinae independently evolve an active defensive tail weapon.
Relative success
Stegosaurs and ankylosaurs, as sister taxa, have their origins at the same time. However, stegosaurs flourish first (in the Middle and Late Jurassic), only to peter out during the Early Cretaceous and disappear before its end. Ankylosaurs are typically rare in the Jurassic (although at one location Gargoyleosaurus is very common), but become abundant in the Cretaceous.
Feeding adaptations:
Narrow-snouted basal thyreophorans and stegosaurs differ from broader-snouted ankylosaurs. The latter have a greatly reduced predentary bone, which may have allowed for more complex motion of the dentaries (for additional munching power) while feeding. The powerful tongue of ankylosaurids hint at some specialized form of feeding.
Group behavior:
Thyreophorans are only rarely found in mass death assemblages, and so (unlike some sauropods, ornithopods, and marginocephalians) probably did not live in large groups. The presence of tail-club generated wounds on ankylosaurids, and the existence of lateral shoulder spines on many non-ankylosaurid ankylosaurs, strongly suggests within-species competition (i.e., dueling matches.)
Display structures:
The spikes, plates, and osteoderms of thyreophorans almost certainly had a defensive function. But many are very broad (particularly stegosaur plates), and the patterns tend to be specific to each species. They may have served an additional function: as visual displays of species recognition. There may also have been a sexual display function to them, but at present it is uncertain if the variation we see in some thyreophorans is from sexual dimorphism or if it is from multiple species of the same genus living together.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}